
江苏东台开庄遗址动物遗存研究报告
2021-07-17 07:21乙海琳甘恢元许晶晶
东南文化 2021年3期
乙海琳 甘恢元 许晶晶
(1.杜伦大学 英国杜伦;2、3.南京博物院 江苏南京 210016)
内容提要:开庄遗址2018-2019年的发掘工作中收获了丰富的动物遗存,包含龟科、大雁、狗、熊、猪、梅花鹿、水鹿、麂、獐、麋鹿、水牛等,尤其以麋鹿等野生动物种类占优。对H13性质的分析及麋鹿各部位出现频率的讨论显示出开庄遗址和其他遗址之间可能存在搬运行为或其他联系。生业经济方面,开庄遗址良渚晚期是以饲养家猪和狩猎野生动物相结合的模式,而到了龙山文化时期,则迅速转变为大量采集狩猎的资源获取模式。
一、遗址基本情况介绍
开庄遗址位于江苏省东台市溱东镇开庄村。1995年,当地砖瓦厂取土首次发现该遗址[1];2018年底,由南京博物院联合东台市博物馆对开庄遗址进行考古勘探并发掘。此次发掘,共发现各类遗迹51个(组),出土各类遗物200余件[2]。
本文将对开庄遗址出土的动物遗存进行介绍和分析,近现代地层的动物遗存只提供部分数据(表一)。

表一// 开庄遗址新石器时代动物骨骼可鉴定标本数(NISP)和最小个体数(MNI)统计表(单位:个)
二、材料与方法
开庄遗址2018—2019年度的发掘共发现动物遗存9384件,总重量约238965克。这些动物遗存绝大多数出土于新石器时代地层,包含良渚文化晚期和龙山文化时期两个部分,其中又以龙山文化地层的堆积较为丰富。由于发掘区域较为集中,无法进一步界定同时期不同遗迹单位中动物遗存的独立性(例如同属良渚文化晚期的两个灰坑,因其在空间上相距较近,很难判断两个灰坑中的动物骨骼是否来自于同一动物个体),因此本文将按照良渚文化和龙山文化两个大的时期,对该遗址出土的动物遗存进行分类和统计。
种属鉴定工作主要参照《动物骨骼图谱》[3]、《考古遗址中的鸟骨鉴定手册》[4]等动物骨骼图谱及部分现生标本照片;动物的性别年龄鉴定主要参照《考古遗址中动物骨骼的年龄性别》[5];动物骨骼的尺寸数据测量参照《考古遗址出土动物骨骼测量指南》[6]等书中的标准。测量工具使用山东大学动物考古实验室统一采买的电子数显卡尺,称重使用电子秤,并制作标签。该遗址出土的动物种属按照林奈生物分类法分类。
(一)良渚文化晚期动物遗存概况
该时期出土动物遗存349件,其中113件仅能鉴定为哺乳动物,根据其尺寸可大致分为大型哺乳动物(85件)和中型哺乳动物(28件)。其余可鉴定到目一级的动物遗存共计236件,包含龟鳖目、雁形目和偶蹄目。
1.龟鳖目(Testudinata)
龟科(Emydidae)腹甲残片85件,至少代表1个个体。
2.雁形目(Anseriformes)
雁属(Anser)仅有1件左侧股骨近端。
3.偶蹄目(Artiodactyla)
(1)猪科(Suidae)
猪属(Sus)共计21件,为上下颌及牙齿、枕骨、肩胛骨、肱骨、尺骨、髋骨等,至少代表不同年龄段的5个个体。
(2)鹿科(Cervidae)
麂属(Muntiacus)共计2件标本,为下颌骨和角。
中型鹿,1件,为左侧下颌游离M3。
麋鹿(Elaphurus davidianus)126件,为角、头骨带角、枕骨、上下颌及牙齿、枢椎、肩胛骨、肱骨、尺骨、桡骨、腕骨、掌骨、髋臼、股骨、髌骨、胫骨、距骨、跟骨、中央跗骨、跖骨、趾骨等。至少代表4个个体。
(二)龙山文化时期动物遗存概况
该时期出土动物遗存共计8999件。1532件标本由于过于破碎或缺少鉴定特征,记为碎骨或残骨,其主要出于H13内;3476件标本仅能鉴定为哺乳动物,根据其尺寸可分为大型(2367件)、中型(1108件)和小型(1件)哺乳动物;其余可鉴定到目一级的动物遗存共计3991件,包含龟鳖目(36件)、雁形目(3件)、食肉目(4件)和偶蹄目(3948件)。
1.龟鳖目(Testudinata)
龟科(Emydidae)腹甲残片36件,因花纹有明显不同,至少代表2个个体。
2.雁形目(Anseriformes)
(1)大型鸭科(Anatidae)肢骨残段1件。
(2)雁属(Anser)左侧胫骨远端2件,至少代表2个个体。
3.食肉目(Carnivora)
桡骨近端1件,因无法确定其具体种属,记为大型食肉动物,该个体可能存在病变。
(1)狗(Canis familiaris)2件,为尺骨和右侧下颌带牙齿,至少代表1个个体。
(2)熊属(Ursus)仅有1件左侧第五掌骨,代表1个个体。
4.偶蹄目(Artiodactyla)
(1)猪科(Suidae)
猪属(Sus)137件,为头骨(顶骨枕骨部)、上下颌及牙齿、寰椎、枢椎、肩胛骨、肱骨、尺骨、桡骨、髋骨、股骨、髌骨、胫骨、跟骨、距骨、掌骨、跖骨、趾骨等。至少代表9个个体。
(2)鹿科(Cervidae)
小型鹿34件,其中可以确定具体种属的是獐属(Hydropotes)犬齿1件和麂属(Muntiacus)左侧头骨带角柄1件,其余部位仅能归为小型鹿,为下颌、肩胛骨、肱骨、桡骨、股骨、胫骨、掌骨、跖骨、近端趾骨等。至少代表4个个体。
中型鹿41件,其中可以确定种属的是梅花鹿(Cervus nippon)角残块2件和水鹿(Rusa unicolor)自然脱落角环部2件,其余部位骨骼归中型鹿,为头骨带角、角、枕骨、前颌骨、下颌及牙齿、肩胛骨、尺骨、胫骨、跟骨、距骨、掌骨、跖骨、末端趾骨等。至少代表3个个体,其中1个为幼年个体。
麋鹿(Elaphurus davidianus)3702件,为角、头骨带角、头骨残块、前颌骨、枕骨、上下颌及牙齿、寰椎、枢椎、颈椎、胸椎、腰椎、荐椎、肩胛骨、肱骨、尺桡骨、尺骨、桡骨、腕骨、掌骨、坐骨、耻骨、髂骨、髋骨、股骨、髌骨、胫骨、距骨、跟骨、跗骨、跖骨、趾骨等。至少代表84个个体,其中70个为成年个体,14个为小于1个月的幼年个体。
(3)牛科(Bovidae)
水牛属(Bubalus)34件,为上下颌游离臼齿、肋骨、肩胛骨、桡骨、腕骨、掌骨、股骨、距骨、跟骨、跗骨、跖骨、趾骨等。至少代表2个个体。
三、数据分析
开庄遗址2018—2019年度的考古发掘共发现动物遗存9384件,除去36件出自近现代地层和采集的标本,其余9348件动物骨骼均为新石器时代,这是本文的研究对象,下文中所有的“总标本数”“总数”等概念均指新石器时代动物标本总数。其中可鉴定标本数(NISP)为4226件,约占总标本数的45.21%。
从文化分期来看,新石器时代的动物遗存中,有8999件属龙山文化时期,可鉴定标本数为3990(44.34%);良渚文化晚期仅出土标本349件,可鉴定标本数为236(67.62%)。
从遗址动物群构成来看,该遗址出土的动物骨骼有爬行动物、鸟类和哺乳动物。不论是良渚文化时期还是龙山文化时期,哺乳动物的数量都是最多的,分别占两个时期标本总数的74.7%和82.0%。哺乳动物中又以大型哺乳动物占绝对主导地位,中型哺乳动物次之,小型哺乳动物仅在龙山文化地层发现1件(图一、图二)。

图一// 良渚文化晚期动物群构成

图二// 龙山文化时期动物群构成
对动物群再细分,新石器时代可鉴定到“科”及以下的标本有龟科、雁属、狗、熊属、猪、梅花鹿、水鹿、麂属、獐属、麋鹿、水牛属。良渚文化晚期,麋鹿的可鉴定标本数最大,龟科次之,猪再次之,雁属、小型鹿、中型鹿的标本数量非常少,而最小个体数则是猪和麋鹿为最多(图三)。由此可见,麋鹿和猪分别在两个维度上数量显著,占据着该时期标本构成的优势地位,而龟科标本在两个维度上的落差应该是由于龟腹甲残块较多造成的统计误差。龙山文化时期,种属之间的数量差异则十分明显,麋鹿无论是在可鉴定标本数还是最小个体数上都占据着绝对优势;猪次之;再就是梅花鹿、水鹿、麂属等小、中型鹿类动物;其他种属仅有零星的几件(图四)。
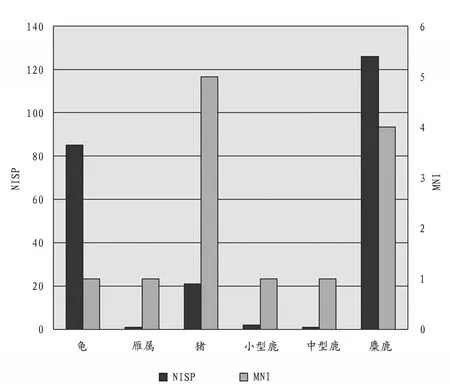
图三// 良渚文化晚期动物骨骼NISP和MNI统计图

图四// 龙山文化时期动物骨骼NISP和MNI统计图
四、讨论
(一)古环境复原
开庄遗址位于江淮东部,如今该地区水网密布,地质地貌上以冲积平原及滨海滩涂地为主,缺乏丘陵及山地地带。
依据均变论原理,可以通过遗址中出现的动物种类来还原该遗址及周边地区的古环境。龟科可半栖水,喜淡水河流、湖泊及沼泽地;雁属多为候鸟,以鱼、虾、水草、种子为食,喜在淡水沿水岸水生植物或水岸植物稠密区觅食,淡水域为其活动范围;野猪可栖息于多样的环境,但喜欢栖居在茂密的灌丛、潮湿的阔叶林或草地[7];熊多栖息于阔叶林或混交林中,可于低丘活动;獐属也称河麂,沿江湖两岸的湿地、苔草地、芒丛、芦苇地以及临近低山、丘陵坡地是獐比较适宜的生存环境,其主要靠芦苇和高草丛隐蔽;梅花鹿的南方亚种喜栖息于植被以芒、草等草本植物和灌木丛为主的山丘;麋鹿喜温暖湿润气候,其蹄宽大分裂,趾间有皮腱膜,适于在沼泽地行走,牙齿纤弱,适于采食鲜嫩水生植物[8]。
综上,开庄遗址新石器时代的古气候环境应该与现在江淮东部地区差不多,整体温暖湿润,沼泽广布,水源丰富。遗址周围河流、湖泊星罗棋布,从而孕育了大量的水岸水生植物,是喜水鸟类、鹿类动物的天堂。遗址地处平原地区,草地广布,远处可能有部分低丘灌木丛以及潮湿的阔叶林,又是野猪、熊、梅花鹿等动物的理想栖息地。
(二)生业模式的探讨
通过对开庄遗址出土动物骨骼数量的比对分析,可以清楚地看到,麋鹿在良渚文化和龙山文化两个时期都占据着优势地位,应该是当时人们的主要肉食来源。麋鹿及各种体型的鹿类动物,均为野生,说明狩猎野生鹿类动物,尤其是麋鹿,是开庄先民生业经济的重要组成部分。
遗址中还出土了一定量的猪骨,判断其是家养还是野生对该遗址的生业经济研究至关重要。笔者将分别对良渚文化晚期和龙山文化时期的猪骨进行讨论。
1.龙山文化时期
因为龙山文化时期的猪骨性质更为清晰明了,所以先对其分析判断。
从数量比例来看,该时期猪的可鉴定标本数和最小个体数的占比分别仅为3.4%和8.3%,与其他同时期出土家猪的安徽尉迟寺[9],山东尹家城[10]、庄里西[11]、丁公[12]等遗址相比较,其猪骨的数量比例相差甚远。由于猪的牙齿尺寸会随着驯化过程而逐渐缩小[13],因此下颌M3尺寸可以作为衡量家猪还是野猪的参考。该时期共发现3件下颌M3标本,分别为:TS1E3③层的游离下颌M31件,M3长44.18毫米;TS1E3⑤a层的左侧下颌带M1-M31件,M3长33.45毫米;TS1E2⑦的右侧下颌带P3-M31件,M3长44.66毫米(图五)。这三件标本,尤其是两件M3长度大于44毫米的标本具有极强的野生猪的形态特征,此外这3件标本M3长度的平均数据约为40.76毫米,也超出了一些学者提出的“下颌M3平均长度小于39毫米(即出现家猪)”[14]的标准。从年龄结构来看,该时期可以判断具体年龄范围的个体有7个(7—9个月1个;10—12个月1个;15—17个月1个;20—21个月1个;21—23个月2个;23—27个月1个),虽然这些个体的年龄大部分落在了2岁以下,似乎符合家猪饲养的年龄结构,但事实上,还有部分标本由于关键牙齿的缺失,只能鉴定到年龄下限,而无法判断上限,导致有不少“大于18/22/25/29个月”(可能大于2岁的个体)的标本无法被统计在内,从而使得表面上死亡年龄偏小,在实际操作中,2岁以上的个体应该更多,而猪的整体年龄结构是偏大的。

图五// 龙山文化时期野猪下颌标本
综上所述,龙山文化时期出土的猪骨应该大概率为野猪,但鉴于遗址年代偏晚,不排除有家猪饲养的可能性。
2.良渚文化晚期
相较于龙山文化时期,良渚文化晚期的动物骨骼虽数量远不及龙山文化时期,但猪骨的成分更为复杂。相对数量比例方面,该时期猪的最小个体数是最高的,高于麋鹿的比例,占总最小个体数的41.7%,虽然猪骨的可鉴定标本数仅占约8.9%,但这个情况与龟腹甲的统计误差是息息相关的,由于龟腹甲较为破碎,导致其统计出的可鉴定标本数较大,这样在一定程度上就会拉低猪骨标本所占的比例。“一个考古遗址出土的主要哺乳动物群中,若猪所占的相对比例(NISP比例、MNI比例或者MW比例都可以)超过30%以上,我们就基本可以肯定该遗址猪群中很可能已经出现了家猪。”[15]基于此,良渚文化晚期的猪骨标本中应该存在家猪。牙齿形态方面,该时期并未发现可以作为牙齿形态参考的下颌M3标本。年龄结构方面,有5个个体可判断具体年龄,其中7—10个月2个,16—17个月2个,21—23个月1个,全部是小于2岁的个体,符合出现家猪饲养的死亡年龄构成。与龙山文化时期情况不同的是,这是该时期全部的牙齿标本,不存在有潜在的个体被忽略的情况,因此基本可以代表该阶段猪的死亡年龄结构。
综上,良渚文化晚期存在家猪,也可能存在部分狩猎来的野生个体,但应该是以家猪为主。
此外,我们还可以通过对比来考察两个时期中猪的性质以及生业模式的不同。从良渚文化晚期的猪(NISP占比8.9%,MNI占比41.7%)到龙山文化时期的猪(NISP占比3.4%,MNI占比8.3%),两个指标均有所下降,而野生麋鹿的数据则直线上升(图六);龙山文化时期猪的年龄构成整体上比良渚文化晚期的要大,该时期野生形态特征也表现得更为明显。尽管两个时期的猪骨性质并不能单纯地以野生或家养来定义,但肉食资源性质从良渚文化时期表现出“家养与野生掺杂”到龙山文化时期“野生占主导”,这种变化本身也可以说明问题,且更突出了二者的不同。此外,植物方面的材料也显示出了相似的规律,与家猪饲养相辅相成的稻作农业,在开庄遗址良渚文化晚期阶段延续着江淮东部的传统并保持着一定强度,而到了龙山文化时期,稻作农业似乎未表现出持续强化的发展态势[16]。从良渚文化晚期到龙山文化时期,开庄遗址的野生动物资源占比迅速增加,野生植物资源也趋于强势,是什么原因导致了该地区肉食资源获取方式的“逆发展”?这值得今后更多地探讨。
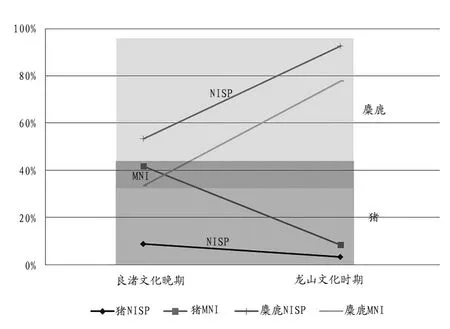
图六// 开庄遗址良渚晚期到龙山时期猪和麋鹿的数量比例变化
开庄遗址地处水网密布的地势平缓地区,自然资源极为丰富,资源域范围很大。动物方面,良渚文化晚期的肉食获取方式可能是以小规模饲养家猪与狩猎相结合的模式,在饲养家猪的同时会捕获一些野生动物以补充肉食资源;而龙山文化时期的先民主要用狩猎的方式来获取资源,以麋鹿为主的大型野生哺乳动物是其主要肉食来源,辅之以水牛、野猪、中小型鹿类动物等。植物方面,采集植物是重要来源,农业在聚落生产中不占绝对优势。稻作是最重要的农耕生产内容,良渚文化晚期保持着一定强度的稻作农业传统,而龙山文化时期并未发现持续强化的态势[17]。骨角器制作方面,人们会将食用后的麋鹿长骨加工成骨镞、骨凿等骨器,獐牙制成牙刀,牛肩胛骨制成骨耜,采集或狩猎到的鹿角加工制作为角器。
(三)H13性质的讨论
H13是龙山文化时期一个非常特殊的灰坑,仅发掘了北半部。在距开口深约21厘米以下部分,堆填了大量未成年哺乳动物的细碎骨骼[18]。这些动物骨骼共约3000件,除34件属于成年大、中型哺乳动物之外,其余均属于未成年个体。整体来看,这些骨骼非常细碎且脆弱,所有部位的骨骺均未愈合(图七),没有发现鹿角。综合各个部位的尺寸和特征,判断这些骨骼应属于大型鹿,再结合遗址中出土了大量麋鹿的信息,认为其应该为麋鹿。另外,发现了27件(左14、右13)保存较好的距骨,基于此,H13中的未成年骨骼遗存至少代表了14个未成年麋鹿个体。

图七// H13未成年麋鹿骨骼愈合状况
对这14个未成年麋鹿个体的年龄判断是进行深入研究的关键。参考唐纳德·查普曼(D.I.Chapman)等对黄麂年龄判断的指标[19],日本学者Hiroko Koike和Noriyuki Ohtaishi关于梅花鹿牙齿萌出时间的标准[20],以及针对马鹿[21]的牙齿萌出时间研究等,这14个未成年麋鹿个体的年龄应该处于围产期,即出生前后,它们可能是刚出生约1周的新生儿,或者是仍未出生的胎儿。最能说明这个问题的是中央跗骨标本,随着动物体重的增加,跗骨愈合生长的速度非常快,但如图七∶3所示,中央跗骨两半并未开始愈合且形状非常小,因此这些未成年小鹿的年龄确实非常年轻。而下颌牙齿的萌出情况也证实了这一点(图八)。

图八// H13未成年麋鹿牙齿萌出状况
这些麋鹿新生儿/胎儿似乎并没有被人们所利用,而是被直接丢弃在了H13之中。综合整个遗址的动物骨骼情况来看,仅在H13中发现了围产期麋鹿骨骼的集中堆积,除了H39中发现的1件小于1个月的麋鹿下颌,其余遗迹单位并未发现,试想如果人们将麋鹿胎儿分而食之或者作他用,必定会在别处发现相似的骨骼,然而事实上并非如此。从常识来说,这些个体发育不完全,肉量及骨髓少,无法提供人们想要获取的肉食资源。可与H13对比的有丹麦中石器时代的采集狩猎遗址——林克洛斯特(Ringkloster)遗址,该遗址出土的动物骨骼中28.4%的马鹿和18.6%的欧洲狍为新生儿或胎儿。但是后者因掌、跖骨远端发现有一圈切割痕迹,作者推测这些幼鹿可能是为了剥皮而被宰杀的[22],这显然与H13不同。
还有一个值得注意的问题,既然是直接将小鹿丢弃至灰坑中,那为什么在发掘现场没有发现完整的骨架?这一点可以从骨骼状态本身和埋藏学角度来分析:首先,这种骨壁薄、骨质松的胎儿骨骼是极为脆弱的,在肌肉、皮毛组织腐烂之后,其很难在漫长的埋藏岁月中保持原本的状态;其次,根据对H13内容物的描述,这些骨骼发现于灰坑开口21厘米以下,说明在人们集中丢弃了一批麋鹿胎儿之后,这个灰坑可能长期暴露在外,受到了外界的扰动。
民族学材料表明每年农历的二、三月份即所谓的“鹿胎期”是母鹿怀孕的季节,最适合猎鹿[23]。猎获的鹿胎自母体取出后,一般并不食用,多被直接丢弃。
综上,笔者推测H13是一处专门用来丢弃麋鹿胎儿的灰坑,在人们将猎杀的怀孕麋鹿带回营地之后,处理过程中将其腹中的胎儿直接丢进了H13,并未对其进一步地利用。
(四)麋鹿各部位出现频率的探讨
麋鹿作为遗址中出现频率最高的动物,是人们的主要肉食来源,其各部位的出现频率可以在一定程度上帮我们还原先民的行为。本文采用MNE(Minimum numbers of elements,即某种动物用于鉴定分类时的解剖学部位的最小数量)和MAU( Minimum animal unit,其用于 MNE 频率之间的对比)两个概念来对解剖学部位的出现频率进行量化分析。宾福德认为对于采集狩猎遗址来说,人们会对动物肢解和搬运,动物某一个部位的出现并不代表其整个个体在遗址中的出现,因此MNI存在着很大的误差,而相较于MNI,MNE和MAU可以更清晰地表现出某个部位在遗址中出现的频率,而不受性别、年龄的影响[24]。
对开庄遗址出土的麋鹿骨骼的MNE、MAU和%MAU[25]进行统计(表二),各部位出现频率(%MAU)见图九。

表二// 开庄遗址麋鹿解剖学单位MNE、MAU及%MAU统计表(单位:个)

图九// 开庄遗址良渚文化、龙山文化、H13(胎儿)麋鹿各部位频率图
由于H13内的胎儿骨骼很可能均为完整个体,因此将H13内的胎儿标本单独进行了计算,一定程度上可以将其作为一个参照标准,但同时也有较大的弊端:H13中的麋鹿个体为骨骼未发育完全的新生儿或胎儿,鉴定过程较为困难,很多具有鉴定特征的部位缺失、未愈合或者是未成形,因此可能会造成某些解剖部位的数据偏差及缺失,如图九H13中的鹿角、头骨和桡骨近端。除去因材料本身限制导致的鉴定结果误差,H13中其余部位的频率分布还是比较均衡的,中轴骨(寰椎、枢椎、颈椎、胸椎、肋骨、腰椎、荐椎、髋骨)、前肢(肩胛骨、肱骨、尺骨、桡骨、掌骨)、后肢(股骨、胫骨、中央跗骨、根骨、距骨、跖骨)以及末端骨骼(趾骨)的丰度都比较高。
由于良渚文化晚期的标本量较小,所以会有部分部位缺失的情况,但良渚文化晚期和龙山文化时期的部位频率分布与走势较为相似,均整体呈现四肢骨多,中轴骨少的规律。具体表现为:后肢关节部(中央跗骨、根骨、距骨)最高,这与3个骨骼部位的形状有关,其不易因受到外力而破碎,更容易在遗址中保存下来,也更容易被辨认;前肢、后肢和头部(包括头骨和角)的频率分布较为平均;而中轴骨出现频率较低(图一〇[26])。成年个体与H13中的未成年个体所表现出的部位频率是明显不同的。

图一〇// 开庄遗址成年麋鹿相对缺失(出现频率低)部位示意图(黑色部分)
在对比成年和未成年个体的部位频率差异之前,先要解释两个现象的存在:第一,良渚文化和龙山文化两个时期的成年麋鹿的下颌及牙齿数量都不多,而相较于马家浜遗址[27]等出土了大量麋鹿牙齿、下颌标本的史前遗址来说,开庄遗址的麋鹿牙齿及下颌出现频率是非常低的;第二,开庄遗址发现的26件(良渚文化晚期2件、龙山文化时期24件)保存相对完整的顶骨都带角(包含角柄、角环和角),且大多发现有加工痕迹。由于雄性麋鹿长角而雌性不长角,因此这些标本都属于雄性个体。开庄并未发现属于雌性个体的不带角的顶骨标本。出现上述牙齿下颌部频率低和仅发现有雄性头骨的两个现象的原因,可能是由于在狩猎过程中,人们为了减少路途上的负担,而直接将沉重的头骨及下颌部位丢弃,很少将其带回营地,但为了提供制作角器的原材料,带角的雄性头骨会被保留。
回到上一个问题,用以食用的成年个体与未被利用的未成年个体之间的部位频率的差异反映了人们对成年麋鹿的特殊处置。那是什么行为造成这样的现象呢?笔者认为有两种可能:第一,营地间的搬运行为。猎人们将猎来的麋鹿带回营地,并对其进行简单肢解,从肩胛骨部剖解下前肢,从股骨部剖解下后肢,四肢留在此处进行食用,而将中轴骨部尤其是胸椎腰椎带肉部位搬运到其他营地,由此推测开庄遗址可能是一个获取营地或是一个对猎物进行前处理的场所。第二,此处为骨、角器取料加工场所。开庄遗址出土了骨器、角器、牙器,以及制作骨、角器的原料、坯料、废料等共计291件[28]。其中很明显有利用麋鹿长骨,尤其是掌、跖骨制作骨镞、骨凿等工具的现象,也出现大量麋鹿角上残留有劈砍、切锯等取料和加工的痕迹,由此推断开庄遗址可能是一处对骨、角器前处理或加工的场所,因此会保留有更多的四肢骨。
上述两种可能并不对立,开庄遗址可能既是一处获取营地,也是一处骨、角器加工地。人们于临近沼泽处狩猎麋鹿,后将其带回营地进行初步肢解,四肢部留在原地直接食用,之后再对其进行简单处理,加工制成工具,而一部分人将麋鹿中轴带肉部搬运到其他的营地或聚居地。
五、结语
长江中下游地区河网密布,自然资源丰富,孕育了众多史前文化。相较于宁绍平原、太湖平原,江淮东部地区开展过系统的动植物研究的遗址还相对较少,开庄遗址的发掘以及后续进行的分支研究,为重建该地区史前社会提供了宝贵的资料。
开庄遗址2018—2019年度的发掘中,动物遗存包含龟科、大雁、狗、熊、猪、梅花鹿、水鹿、麂、獐、麋鹿、水牛,数量以野生哺乳动物为主,尤以麋鹿为甚。良渚文化晚期是以饲养家猪和狩猎野生动物相结合的生业经济模式,而龙山文化时期的肉食来源以狩猎野生鹿类动物为主。通过对H13性质以及麋鹿各部位出现频率的讨论,开庄遗址可能为一处获取营地,是作为猎人们对猎物进行前处理以及制作骨、角器的场所,各营地间存在搬运行为。
值得注意的一点是,开庄遗址的两个阶段——良渚文化晚期和龙山文化时期,处于新石器时代晚期甚至末期,当时均已经出现了农业种植和家畜饲养的技术。距今5300—4300年的良渚文化时期,农业经济达到全盛,形成了以水稻生产与家猪饲养相结合的生业经济模式[29],而龙山文化时期的众多遗址也多形成了以食用家猪为主的肉食结构[30]。在这种大趋势下,开庄先民反其道行之,选择了狩猎(或者是半家养半狩猎)的肉食获取方式,动物构成以野生大型鹿为主。这个情况在江淮下游地区并不是个例,良渚文化时期的上海广富林遗址[31]、江苏蒋庄遗址[32],马桥文化时期的上海马桥遗址[33]、江苏绰墩遗址[34]的动物遗存也均显示出了以野生动物为主的现象。这些遗址所显示出的共同现象,为我们今后研究良渚文化与后良渚时期的社会经济与社会进程,以及该时期南北方文化的交流融合提供了思路和资料。
猜你喜欢
军事文摘(2022年16期)2022-10-28
初中生学习指导·中考版(2020年12期)2020-09-10
文化交流(2020年4期)2020-04-20
环球慈善(2019年7期)2020-01-06
杭州(2019年26期)2019-09-08
当代工人·精品C(2016年6期)2017-01-12
少年科学(2009年12期)2009-07-07
小哥白尼·趣味科学画报(2009年5期)2009-06-19
