
西瓜 PIP5K基因家族鉴定及其在雄性不育花蕾中的表达分析
2021-07-16 06:36:32张高原魏兵强
西北农业学报 2021年6期
张高原,丁 谦,魏兵强
(1.甘肃农业大学 园艺学院,兰州 730070;2.潍坊工程职业学院 花卉学院,山东青州 262500)
磷脂酰肌醇4-磷酸5-激酶(PIP5K)是磷脂酰肌醇(phosphatidylinositol,PI)信号转导途径中的关键酶,在多种细胞的功能发挥方面起着重要调控作用[1]。大多数PIP5K蛋白的N端具有多个MORN(Membrane Occupation and Recognition Nexus)结构域,C端具有1个起催化功能的PIPKc结构域,这些结构域在真核生物中高度保守[2-3]。研究表明,PIP5K基因对植物的多个组织器官的发育起调控作用,如气孔[4]、根[5-7]、花[8-11]等;此外,还参与调控植物逆境胁迫响应,如盐害[12]、水分胁迫[13]等。目前,在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、银杏(Ginkgobiloba)和大豆(Glycinemax)中分别发现11、11、7和22个PIP5K基因[14],部分基因已被克隆。
虽然西瓜基因组数据已公布[15],但是关于西瓜PIP5K基因的鉴定及其在雄性不育花蕾中的表达分析未见相关报道。本试验拟利用生物信息学方法对西瓜PIP5K基因进行筛选和鉴定,并对其蛋白理化性质、结构域、基因结构、motif分布、染色体定位、进化树、共线性、顺式作用元件等特征以及组织表达模式进行分析,以期为进一步研究其功能奠定前期基础。
1 材料与方法
1.1 西瓜花蕾材料
西瓜雄性两用系中可育系‘MF-1’和不育系‘MS-1’种子均由甘肃农业大学瓜类研究所提供。种子于2020年4月22日播种于兰州市皋兰县忠和镇实试验田,于7月8日分别采取雄性可育株和不育株的子蔓茎尖花蕾(长度为3~5 mm)和茎尖下方4~5节处花蕾(长度为7~9 mm),每种花蕾至少采集30个,储存在液氮中保存,备用。
1.2 西瓜PIP5K家族成员的获取
首先从西瓜基因组数据库(ftp://cucurbitgenomics.org/pub/cucurbit/genome/ watermelon/97103/v1/)[15]下载蛋白组和基因组序列,其次利用Blast本地化软件(Blast2.2.28)(https://blast.ncbi.nlm.nih.gov/Blast.cgi)构建西瓜蛋白组和基因组数据库,然后以Pfam数据库(http://pfam.xfam.org)[16]中的PIP5K保守结构域的隐马尔可夫模型(Hidden Markov Model; HMM)文件(序列号为PF01504)为搜索序列,利用HMMER 3.2软件(http://www.hmmer.org/)[17]在本地西瓜蛋白组数据库中进行BlastP搜索,阈值为E-value<10-10,获取西瓜PIP5K候选成员。为确保候选成员中含有PIPKc结构域,利用SMART (http://smart.embl.de/)在线软件鉴定西瓜PIP5K候选成员。最后从本地西瓜蛋白组和基因组数据库中下载鉴定后的西瓜PIP5K家族成员的蛋白和基因序列,作为后期分析的基础。拟南芥PIP5K成员[14]的基因和蛋白序列来源于拟南芥数据库(http://www.arabidopsis.org/)。
1.3 西瓜PIP5K家族成员基本信息分析
蛋白序列长度、分子量及等电点等理化特征利用ProtParam(http://web.expasy.org/protparam/)在线软件进行分析。亚细胞定位分别利用Euk-mPLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/euk-multi-2/)[18]和ProtComp version 9.0 server(http://www.softberry.com)进行分析。根据基因序列起始位点,利用MapChart 2.2软件绘制西瓜PIP5K基因染色体分布图。利用MCScanX软件(http://chibba.pgml.uga.edu/mcscan2/)[19]进行西瓜和拟南芥PIP5K基因的共线性分析,结果用circos软件(http://circos.ca/)绘制[20]。进化树由MEGA-X软件[21]构建,序列比对方法均为ClustalW,Bootstrap的重复次数设置为1 000,其中西瓜PIP5K家族成员的蛋白和编码序列进化树的构建方法均为邻接法(Neighbor-joining,NJ),执行参数分别为p-distance和pairwise deletion[22-24],剩余设置选择默认选项;拟南芥和西瓜PIP5K家族成员的蛋白和编码序列系统进化树的构建方法均为最大似然法(Maximum Likelihood,ML),执行参数分别为Jones-Taylor-Thornton(JTT) model、Gamma Distributed (G)和Use all sites[24],剩余设置选择默认选项。利用OrthoFinder 2.3.11软件[25](http://www.steve-kellylab.com/software/orthofinder)分析拟南芥和西瓜PIP5K基因家族之间的直系同源基因和旁系同源基因。利用MEME 5.1.1软件[26](http://meme-suite.org/)对PIP5K蛋白序列进行motif分析,其中motif基序长度范围为6~50,数量最大为10,然后利用WebLogo 3.7.4(http://weblogo.threeplusone.com/)生成motif基序标签[27]。利用GSDS2.0(Gene Structure Display Server)(http://gsds.gao-lab.org/index.php)进行基因结构分析[28]。截取西瓜PIP5K编码基因上游2 000 bp序列,利用PlantCARE软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行顺式作用元件分析[29],结果用Evolview v2绘制[30]。
1.4 西瓜PIP5K家族成员转录组分析
利用NCBI-SRA数据库(https://www.ncbi.nlm.nih.gov/sra/)下载西瓜栽培品种97103的生长点、果柄、果肉、下胚轴、叶和根相关的RNA-seq数据(SRP192188)[31],然后利用HISAT 2.2.0软件[32](https://daehwankimlab.github.io/hisat2/)进行RNA-seq分析,R语言的 Rsubread包的featureCounts进行TPM(Transcripts Per Million)表达定量分析[33-34]。
1.5 西瓜花蕾材料的采集及表达分析
利用TRNzol Universal总RNA提取试剂(TIANGEN,中国北京)提取西瓜雄性可育和不育花蕾总RNA。利用PrimeScriptTM RT试剂盒(TaKaRa,中国大连)合成cDNA。利用Primer 5.0设计西瓜PIP5K基因以及内参基因 (β-actin; Cla007792)的特异性引物(表1)。采用罗氏 LightCycler96 实时荧光定量PCR仪(Roche,Basel,Switzerland)进行基因表达量检测。Quantitative real-time PCR (qRT-PCR)反应体系如下:10 μL SYBR Green Master mix (2×),0.8 μL primer F(10 μmol/L),0.8 μL primer R(10 μmol/L),6.4 μL ddH2O和2 μL cDNA模板。PCR反应程序如下:95 ℃预变性30 s,然后95 ℃ 5 s,60 ℃ 30 s 进行50个循环。以雄性可育花蕾(长度为3~5 mm)为对照,计算其他花蕾中ClaPIP5K基因相对表达量,设置3次重复,计算方法采用2-△△Ct方法[35]。采用SPSS17软件中的t检验对基因表达量进行统计分析。

表1 西瓜 PIP5K基因的qRT-PCR引物Table 1 qRT-PCR primers of watermelon PIP5K genes
2 结果与分析
2.1 ClaPIP5K成员的染色体定位和共线性分析
西瓜基因组中共鉴定出8个PIP5K家族成员,根据其在染色体上的位置进行编号 (ClaPIP5K1~ClaPIP5K8)。结果如图1-A所示,西瓜数据库中将无法定位的染色体片段组合起来称为0号染色体,除了ClaPIP5K1定位在0号染色体外,其余7个ClaPIP5K基因分布在其他6条染色体上,其中5号染色体分布2个ClaPIP5K基因,其他5条染色体分别分布1个ClaPIP5K基因。为了检测ClaPIP5K和AtPIP5K基因间的进化关系,利用MCScanX软件进行共线性分析。结果如图1-B所示,4个ClaPIP5K基因与5个拟南芥AtPIP5K基因存在共线关系,如ClaPIP5K7分别与AtPIP5K6、AtPIP5K9和AtPIP5K10共线,ClaPIP5K8分别与AtPIP5K1和AtPIP5K2共线,ClaPIP5K3与AtPIP5K2共线;此外,1个ClaPIP5K基因对(ClaPIP5K5~ClaPIP5K7)发生片段复制事件。
2.2 ClaPIP5K成员的蛋白和基因信息
ClaPIP5K基因的CDS序列长度为606~ 2 499 bp,蛋白序列长度为201~832 aa,分子质量为22.62~93.76 ku,等电点介于6.6~ 8.87,其中4个蛋白呈碱性(表2)。亚细胞定位分析显示多数ClaPIP5K蛋白位于细胞质或细胞膜上(表2)。蛋白结构域分析显示,所有的ClaPIP5K成员在C端都具有1个起催化功能的PIPKc结构域,其中6个ClaPIP5K成员在N端还具有 7~8个MORN结构域(图2)。

表2 西瓜PIP5K家族成员的基本信息Table 2 Information of PIP5K family members in watermelon
2.3 ClaPIP5K成员的蛋白序列进化树及保守基序分析
为了检测ClaPIP5K和AtPIP5K成员的蛋白序列进化情况以及同源序列对(组)所包含的保守基序数量及种类,利用MEGA X软件进行进化树分析,Orthofinder软件进行同源序列分析,MEME软件进行保守基序分析。结果如图3所示,蛋白序列进化树显示ClaPIP5K和AtPIP5K成员被分成4个组(GroupⅠ~Ⅳ),其中 GroupⅠ所含成员最多,为6个,GroupⅡ和Ⅲ次之,分别为5个,GroupⅣ最少,为3个。蛋白保守基序分析显示ClaPIP5K蛋白中含有10个保守基序,每个保守基序含有29~50个氨基酸,其中motif1、4、5和7所含氨基酸数量最多,平均为50个,motif10所含最少,为29个。此外,GroupⅠ~Ⅲ亚组中,除了ClaPIP5K1和AtPIP5K3外,其他PIP5K成员中均含有10个motif。同源序列分析发现ClaPIP5K和AtPIP5K之间存在5个同源序列对(组),其中4个同源序列对(组)(ClaPIP5K4/AtPIP5K4/AtPIP5K5、ClaPIP5K3/ClaPIP5K8/AtPIP5K1/AtPIP5K2、ClaPIP5K7/AtPIP5K9和ClaPIP5K2/ ClaPIP5K6)所含 motif数量和种类均相似。而ClaPIP5K1和ClaPIP5K5由于部分氨基酸的缺失,导致其motif种类和数量上与同组的AtPIP5K成员有所差异。
2.4 ClaPIP5K成员的基因序列进化树及基因结构分析
为了检测ClaPIP5K和AtPIP5K成员的基因序列进化情况以及同源序列的内含子和外显子分布情况,利用MEGA X软件进行进化树分析,Orthofinder软件进行同源序列分析,GSDS软件进行基因结构分析。结果如图4所示,基因序列的系统进化树和同源序列分析结果与蛋白序列的(图3)基本一致;基因结构分析显示75%(6/8)ClaPIP5K基因含有7个内含子(图4),并且4个同源序列对(组) (ClaPIP5K4/AtPIP5K4/AtPIP5K5、ClaPIP5K3/ClaPIP5K8/AtPIP5K1/AtPIP5K2、ClaPIP5K7/AtPIP5K9和ClaPIP5K2/ClaPIP5K6)所含内含子数量均为7个。另外,与AtPIP5K6相比,ClaPIP5K2和ClaPIP5K6的第一个内含子发生了缺失现象,ClaPIP5K1和ClaPIP5K5出现了部分内含子和外显子的缺失现象,导致与同组的AtPIP5K基因在基因结构上有所差异。
2.5 ClaPIP5K成员的顺式作用元件分析
为了明确ClaPIP5K基因启动子所含的顺式作用元件种类及分布情况,利用PlantCARE软件进行分析。结果如图5所示,ClaPIP5K基因启动子处主要包含2大类顺式作用元件:①激素响应元件,如响应生长素IAA的AuxRR-core(1/8;含元件基因数/总基因数,下同),响应赤霉素GA 的TATC-box(1/8)、GARE-motif(2/8)和P-box(4/8),响应水杨酸SA 的TCA-element(5/8),响应脱落酸ABA的ABRE(4/8)和响应茉莉酸甲酯MeJA的CGTCA-motif(3/8)等元件;②胁迫响应元件,如响应干旱的MYB结合位点MBS(5/8),响应低温的LTR(4/8),响应防御与胁迫的TC-richrepeats(5/8),厌氧诱导的ARE(7/8)和响应损伤的WUN-motif(6/8)等元件。其中ClaPIP5K4基因所含顺式作用元件种类和数量最多,共9种16个(1个AuxRR-core、1个GARE-motif、1个TCA-element、2个ABRE、1个CGTCA-motif、1个LTR、1个TC-richrepeats、7个ARE和1个WUN-motif),ClaPIP5K3所含顺式作用元件种类和数量最少,共3种6个(1个MBS、4个ARE和1个WUN-motif)。另外,ClaPIP5K2和ClaPIP5K6所含顺式作用元件种类和数量较相似。
2.6 ClaPIP5K基因在不同组织中的表达模式分析
为探索ClaPIP5K基因在西瓜生长发育过程中潜在的生物学功能,对已公布的西瓜栽培品种97103的生长点、果柄、果肉、下胚轴、叶和根的RNA-seq数据进行组织表达模式分析。结果如表3所示,4个ClaPIP5K基因在6个组织的不同发育时期表现出较高的表达量(TPM>10),其中ClaPIP5K1基因在授粉34 d后的生长点中表达量最高,ClaPIP5K3基因在授粉18 d后的叶片中表达量最高,ClaPIP5K8和ClaPIP5K7基因分别在授粉10 d和26 d后的果肉中表达量最高。此外,ClaPIP5K5基因仅在生长点的4个时期表现出较高的表达量,具有明显的组织表达特异性。另外,本试验利用qRT-PCR技术检测了ClaPIP5K基因在西瓜雄性可育和雄性不育花蕾中的表达分析。结果如图6所示,西瓜雄性可育植株花蕾和花药发育正常、饱满(图6-A),而不育植株花蕾和花药发育不正常、干瘪(图6-B);另外,基因表达分析中(图6-C),除了ClaPIP5K5基因外,其他7个ClaPIP5K基因在西瓜雄性可育和不育花蕾中均检测出不同程度的表达量。其中,与可育植株的7~9 mm长的花蕾对比后发现,4个ClaPIP5K基因(ClaPIP5K1、ClaPIP5K3、ClaPIP5K7和ClaPIP5K8)在不育花蕾中具有明显的较高表达量,然而ClaPIP5K2表现出明显的较低表达量;此外,在3~5 mm和7~9 mm长的花蕾中,ClaPIP5K4基因在不育花蕾的表达量均明显低于可育花蕾,然而ClaPIP5K6基因出现相反的表达趋势。以上结果表明ClaPIP5K基因可能在西瓜生长点、叶片、果肉以及花蕾花药等组织生长发育方面具有重要作用。
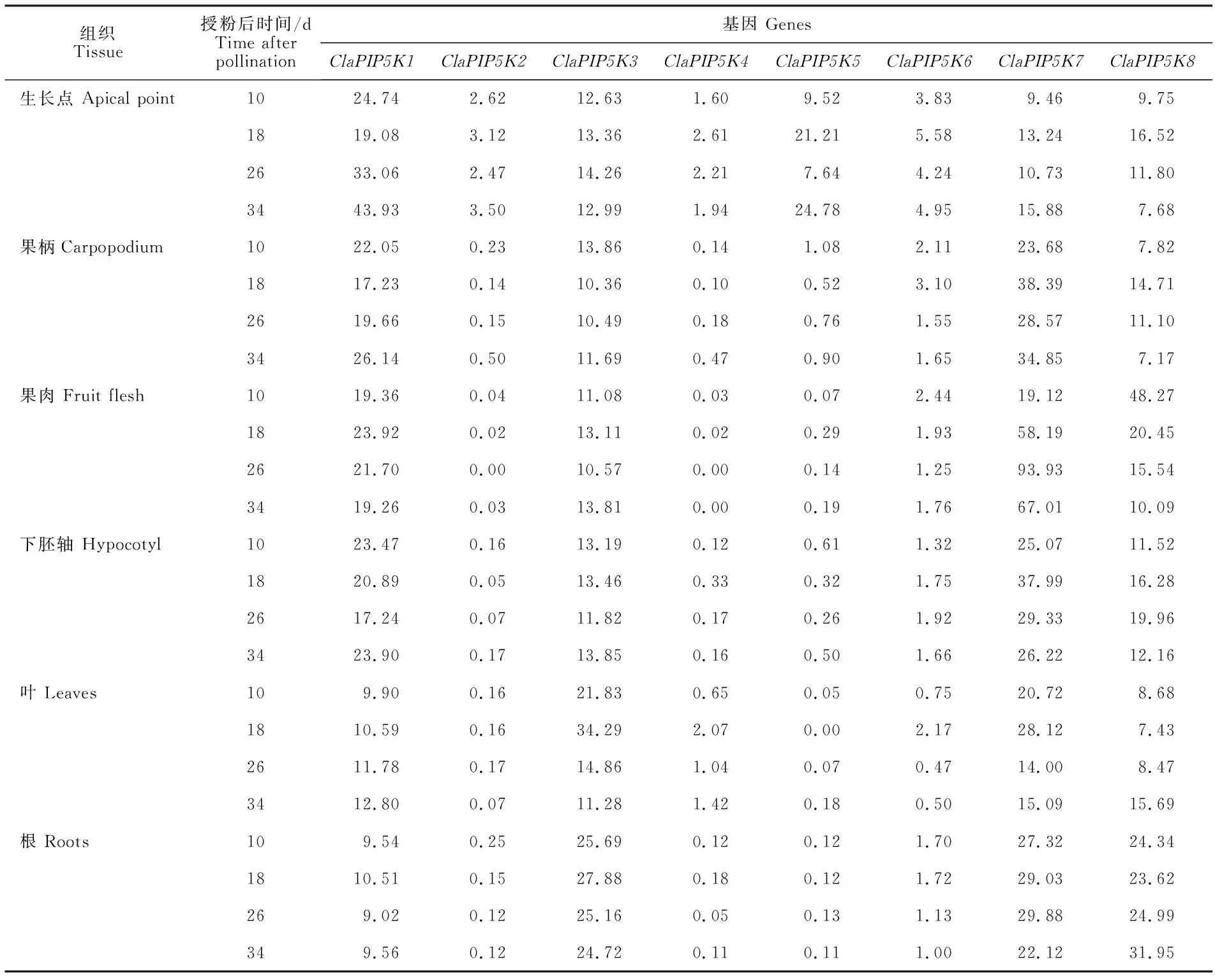
表3 ClaPIP5K基因在西瓜栽培品种‘97103’的不同组织中的表达分析Table 3 Expression analysis of ClaPIP5K genes in different tissues of watermelon cultivar ‘97103’
3 讨论与结论
磷脂酰肌醇4-磷酸5-激酶(PIP5K)是植物内一种磷酯类激酶,在不同植物中的成员数量有所不同。拟南芥中发现含有11个PIP5K成员,大致分成4组(Ⅰ~Ⅳ)。除了第Ⅳ组含有2个AtPIP5K成员外,其他组均含有3个。西瓜含有8个ClaPIP5K成员,除了第Ⅰ组所含ClaPIP5K成员数量与拟南芥一样外,其他组的数量均比拟南芥的少1个,说明这3组(Ⅱ~Ⅳ)的ClaPIP5K成员在进化过程中出现了缺失现象。除了C端具有起催化功能的PIPKc结构域外,N端多MORN结构域也是植物大多数PIP5K家族成员所特有的特征[2-3]。例如,拟南芥中有9个AtPIP5Ks具有7~8个MORN结构域,水稻中有5个OsPIP5Ks 具有7~8个MORN结构域[2],而西瓜中也发现6个ClaPIP5K成员具有7~8个MORN结构域(图1)。亚细胞定位预测显示多数ClaPIP5K定位在细胞质或细胞膜上,说明这些基因可能在细胞质或细胞膜上行使其功能。以前的研究也证实了PIP5K基因作为一种细胞膜传感器,在磷脂类代谢中发挥着重要作用[36]。
通过进化树、同源序列、保守基序和基因结构等分析,发现位于同一组的拟南芥和西瓜PIP5K成员在保守基序种类和基因结构上具有高度的相似性。例如4个同源序列对(组) (ClaPIP5K4/ AtPIP5K4/ AtPIP5K5、ClaPIP5K3/ ClaPIP5K8/ AtPIP5K1/ AtPIP5K2、 ClaPIP5K7/AtPIP5K9和 ClaPIP5K2/ ClaPIP5K6)都具有10个motifs和7个内含子。但是也发现了一些差异,如AtPIP5K6与 ClaPIP5K2/ ClaPIP5K6的motif种类分布上相似,但由于AtPIP5K6在N端多了1个内含子,其基因结构与 ClaPIP5K2/ ClaPIP5K6有所不同; ClaPIP5K1和ClaPIP5K5由于大量序列的丢失,导致与同组的AtPIP5K在保守基序分布和基因结构上差异较大。以上结果表明多数PIP5K成员在进化过程中相对保守。
基因复制事件在基因家族成员数量扩增、生物多样性和物种形成等方面具有重要作用[14]。本研究发现,4个ClaPIP5K基因与5个拟南芥AtPIP5K基因存在复制情况,1个ClaPIP5K基因对(ClaPIP5K5-ClaPIP5K7)存在片段复制现象,说明全基因组复制或者片段复制在ClaPIP5K基因复制中扮演着重要作用。
启动子分析显示ClaPIP5K基因含有响应IAA、GA、ABA、SA、MeJA等激素以及干旱、低温等非生物胁迫等方面的顺式作用元件,这与玉米PIP5K基因启动子生信分析结果相似[37]。基于已公布的西瓜RNA-seq数据,分析了ClaPIP5K基因在不同组织的不同发育时期的表达模式。结果显示4个ClaPIP5K基因 (ClaPIP5K1、ClaPIP5K3、ClaPIP5K7和ClaPIP5K8)在生长点、果柄、果肉、下胚轴、叶和根的不同发育时期均表现出较高的表达量,而ClaPIP5K5只在生长点表现出较高的表达量,说明ClaPIP5K基因可能协同或者单独地参与调控西瓜不同组织形态建成相关途径。此外,以前研究表明拟南芥AtPIP5K4或AtPIP5K5缺失突变株的花药和花粉发育受到严重抑制[9-10]。本研究发现ClaPIP5K4基因在雄性不育花蕾发育的 2个阶段中一直维持着显著的低表达量状态,并且ClaPIP5K4与AtPIP5K4和AtPIP5K5为同源基因,表明ClaPIP5K4基因可能参与调控西瓜花药或花粉的生长发育。总之,本研究首次利用生物信息学方法对西瓜PIP5K基因家族进行了鉴定,包括蛋白理化性质、结构域、保守基序、基因结构、进化情况、启动子以及组织表达模式等方面的分析,为后期研究其功能奠定了一定基础。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
生命科学研究(2018年1期)2018-05-29 01:12:47
作文与考试·小学低年级版(2017年13期)2017-07-06 21:15:38
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
第二课堂(课外活动版)(2015年5期)2015-10-21 19:37:04
作文与考试·小学低年级版(2015年8期)2015-09-25 01:15:53
