
金边鲤群体的遗传结构及个体间遗传差异的QTL标记分析
2021-07-15 12:33:52鲁翠云杜雪松郑先虎孙志鹏孙效文
淡水渔业 2021年4期
鲁翠云,杜雪松,郑先虎,李 超,程 磊,孙志鹏,孙效文,陈 忠
(1.中国水产科学研究院黑龙江水产研究所,哈尔滨 150070;2.广西壮族自治区水产科学研究院,南宁 530021)
鲤(CyprinuscarpioL.)在我国具有悠久的养殖和选育历史,形成了许多极具地方特色的优良种质,禾花鲤就是华南一带适合稻田养殖的特色地方种质,因其在稻田中养殖,采食落水禾花而得名。经过长期的环境适应和定向选育,不同区域又选育出了不同的禾花鲤地方品系,例如桂林品系、华南鲤、石鲤等,普遍具有的特点是头小腹圆、不善跳跃、易捕捞,适合稻田养殖生态环境[1]。近几年,稻渔综合种养模式在全国迅速推广,2018年已经有27个省份开展了稻渔综合种养,水产品产量达到233万吨,比2017年增加了19.81%,其中以鲤、鲫为主养鱼类的稻田养鱼占42.10%[2]。金边鲤(Cyprinuscarpiovar.Jinbian,暂定名)是由广西壮族自治区水产引育种中心联合多家单位针对稻田养殖从融水田鲤中选育出的禾花鲤新品系,因背鳍两侧从头部顶端到尾部的皮肤为金色,似一条金边而得名,具有肉多刺柔、肉质鲜美、无腥味等特点[3],研究表明稻田养殖金边鲤的肌肉品质优势明显[4],市场价格较高,具有广阔的产业开发前景。
在品种选育和使用过程中注重对遗传多样性的保护,最大限度地避免近亲繁殖不仅能够优化种群结构、提高生产性能,还能够延长选育品种的使用寿命。因此,对亲本群体进行遗传多样性评估,制定有效的繁殖配组策略就成为重点。基于亲本间遗传距离的鲤分子育种技术已经开发多年,其中以微卫星标记计算的遗传距离指导家系和群体选育已经在鲤[5,6]、大黄鱼(Larimichthyscrocea)[7]、红鳍东方鲀(Takifugurubripes)[8]、牙鲆(Paralichthysolivaceus)[9]、半滑舌鳎(Cynoglossussemilaevis)[10]等选育中进行了应用,取得了良好的选育效果。研究表明随着亲本间遗传距离的增加,家系平均体质量等生长性状呈现出先升高后下降的趋势,选择遗传距离中等的亲本更易于获得优良家系[5];进一步的研究发现,选育亲本群体雌、雄个体间遗传距离也呈现出正态分布的特点,选择遗传距离中间值作为配组的依据切实可行[11]。基于分子标记的遗传距离预测子代的生长优势在农作物开展的研究较多,研究表明使用与目标性状紧密连锁的功能基因标记或QTL标记计算遗传距离能够增强预测效果[12,13]。然而,以往在评估鲤亲本个体间遗传差异时,使用的多为随机多态的微卫星标记,缺乏与目标性状的关联性。随着鲤基因组资源的开发[14],与生长性状紧密连锁的QTL标记已经积累了很多,其中不乏在鲤品种间共享的QTL标记[15,16]。本研究用24个与体质量、体长等生长性状相关的QTL标记对金边鲤亲本群体进行遗传评估,分析雌、雄个体间的遗传距离分布,为金边鲤的繁殖配组及持续利用提供参考。
1 材料与方法
1.1 实验材料
从广西壮族自治区融水地区收集背鳍两侧从头部顶端至尾鳍基部有完整金色鳞片的个体组成金边鲤基础群体,养殖于广西农业良种海南南繁育种基地。选择体形匀称、无伤病且性腺发育良好的个体192尾组成亲鱼群体,其中雌性个体86尾,雄性个体106尾。对每尾亲鱼植入电子标记,记录每个样本电子标记号,并剪取部分鳍条组织,编号后置于滤纸晾干,以备提取基因组DNA[17]。
1.2 微卫星标记筛选
从鲤高密度连锁图谱及镜鲤生长性状显著相关的QTL标记中筛选24个用于金边鲤亲本群体的遗传分析[14,15]。这些标记分布于11个连锁群,其中2个连锁群含有镜鲤与建鲤共享QTL区间,12个标记是镜鲤与建鲤共享QTL标记[15],在鲤种间具有较好的保守性和适用性(表1)。
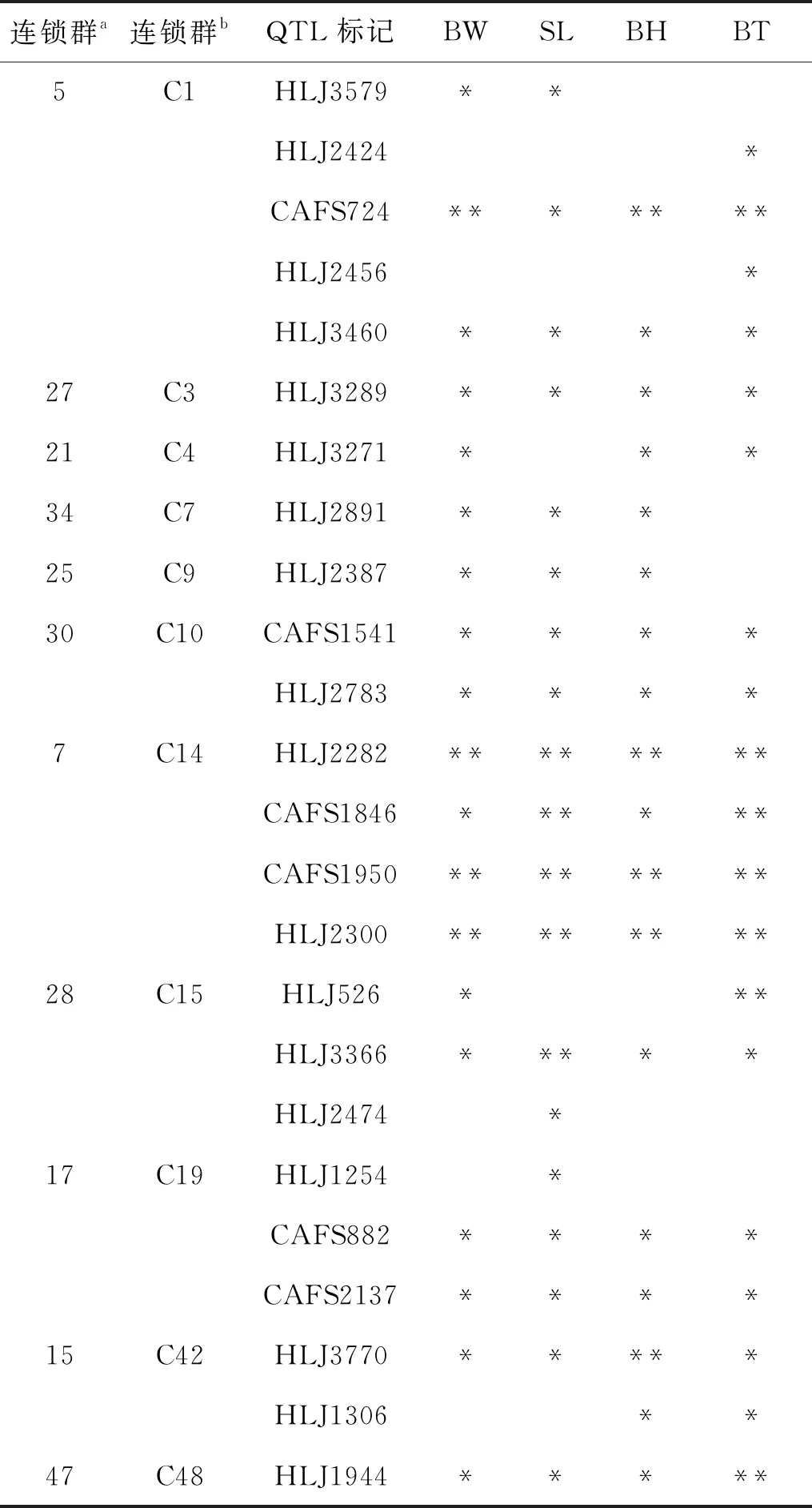
表1 鲤QTL标记所在连锁群与生长性状的相关性
1.3 PCR扩增及检测
用组织基因组DNA提取试剂盒(天根生物)从鳍条组织中提取金边鲤基因组DNA,紫外分光光度计定量后稀释成50 ng/μL。采用毛细管凝胶电泳技术检测192个样本的基因型,在正向引物的5′端标记蓝色(FAM)、绿色(HEX)或黄色(TAMRA)荧光,PCR反应体系含有10 mmol/L Tris-HCl(pH 8.0)、50 mmol/L KCl、1.5 mmol/L MgCl2、200 μmol/L dNTP、0.2 mmol/L上下游引物、1 U Taq DNA聚合酶及100 ng DNA模板。反应程序为:94 ℃预变性3 min;94 ℃变性30 s,56 ℃复性30 s,72 ℃延伸30 s,25个循环;最后72 ℃延伸5 min。反应结束后,各取0.7 μL的三色荧光PCR产物(共2.1 μL),5.9 μL的Hi-DiTM甲酰胺和0.1 μL LIZ-500制备成电泳混合样。将混合样置于PCR仪上于95 ℃变性5 min,立即置于冰上冷却5 min,在3730XL遗传分析仪(ABI)上进行毛细管凝胶电泳,电泳结束后利用GeneMap-per V4.1软件进行图像收集和数据分析,建立个体基因型档案。
1.4 数据处理
使用软件“鱼类种质资源遗传分析装置(ZL200710144749.3)”进行数据转换,用PopGene32(Version 3.2)软件计算每个微卫星标记的等位基因频率(P)、等位基因数(No)、有效等位基因数(Ne)、观测杂合度(Ho)和期望杂合度(He)。标记的多态性信息含量(PIC)按照Botstein等[18]的方法计算,公式如下:
其中,n为某一位点上等位基因数,Pi、Pj分别为第i和第j个等位基因在群体中的频率,j=i+1。
用GenePop(Version 4.7)软件进行χ2检验估计群体Hardy-Weinberg平衡偏离。用PopGene32软件计算每个位点群体的近交系数(Fis),近交系数为正值表明群体在该位点杂合子缺失,负值表明该位点杂合子过剩。用NeEstimator(version 2.1)软件采用连锁不平衡(linkage disequilibrium,LD)方法估算有效群体大小。
运行PHYLIP(version3.695)软件的gendist程序,计算两两雌、雄个体间的遗传距离,用neighbor程序绘制基于邻接法(neighbor joining method,NJ)的聚类图,划分繁殖组,并用Arlequin 3.11软件中的分子方差分析(AMOVA)计算繁殖组间的遗传变异组成及两两繁殖组间的遗传分化系数(Fst)。
2 结果
2.1 扩增情况
24个QTL标记在金边鲤亲本群体扩增获得清晰、稳定的条带,并在个体间表现出多态性。扩增的片段大小为100~310 bp,各标记检测到的等位基因数(No)在4个(HLJ3579)到12个(HLJ1944)之间,共检测到184个等位基因,群体平均等位基因数为7.667个。根据Botstein等[18]对标记多态水平的划分标准,22个标记为高度多态(PIC≥0.5),只有HLJ2300和CAFS882分别为中度多态(0.25≤PIC<0.5)和低度多态(PIC<0.25)水平的标记。
2.2 亲本群体遗传结构分析
用24个QTL标记分析了金边鲤亲本群体的遗传结构,有效等位基因数(Ne)为1.170~7.132,平均为3.941;观测杂合度(Ho)为0.156~0.859,平均为0.590;期望杂合度(He)为0.146~0.862,平均为0.707;多态信息含量(PIC)为0.138~0.845,平均为0.670,结果显示金边鲤亲本群体处于高度多态水平(PIC≥0.5)。
对金边鲤亲本群体进行Hardy-Weinberg遗传平衡评估,结果经Bonferroni校正显著性阈值后,群体尚有10个位点显著或极显著偏离Hardy-Weinberg遗传平衡,其中HLJ2456、HLJ3271、HLJ2387等9个位点表现为杂合子极显著缺失(P<0.000 4)。群体近交系数能够直观地表现出杂合子的缺失或过剩,结果群体在19个位点出现杂合子不同程度的缺失,占79.17%;仅5个位点表现为杂合子过剩。采用LD方法估算金边鲤亲鱼有效群体大小为113.6尾(CI95%)。具体统计数据见表2。
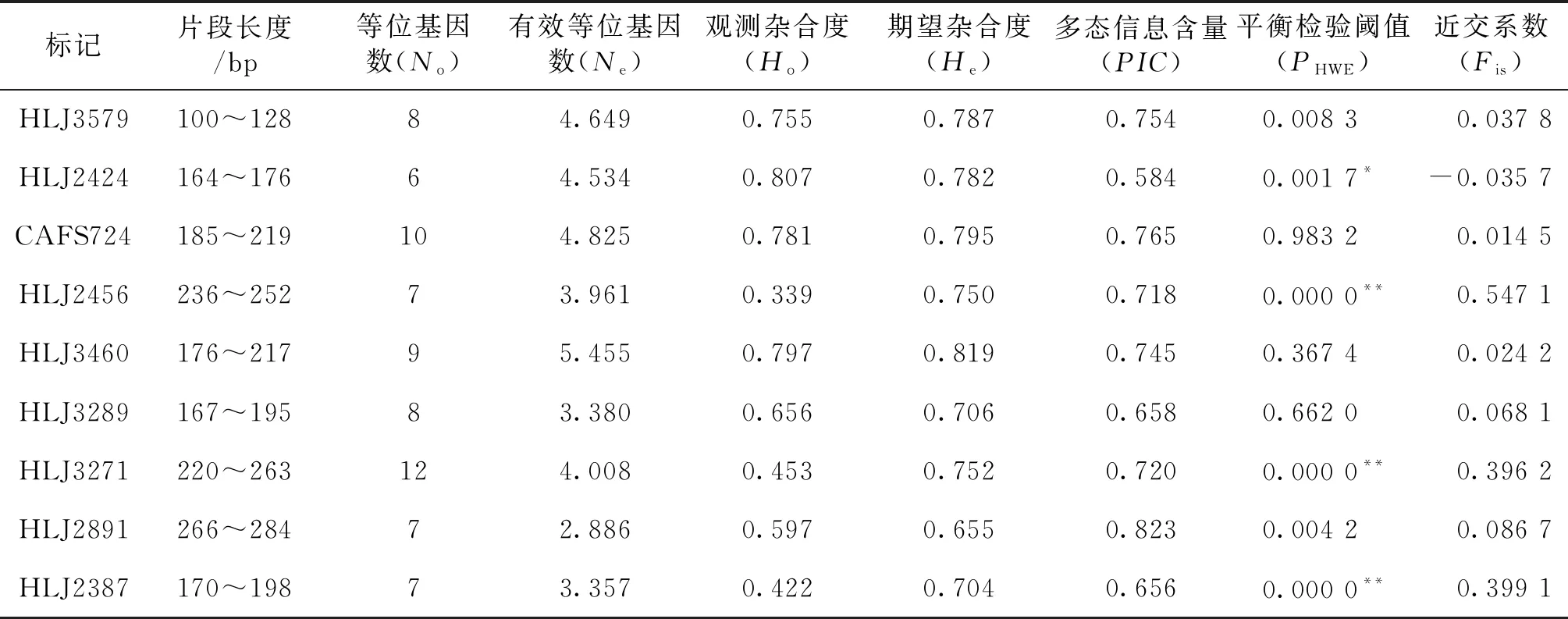
表2 金边鲤亲本群体在24个QTL标记的遗传多样性参数
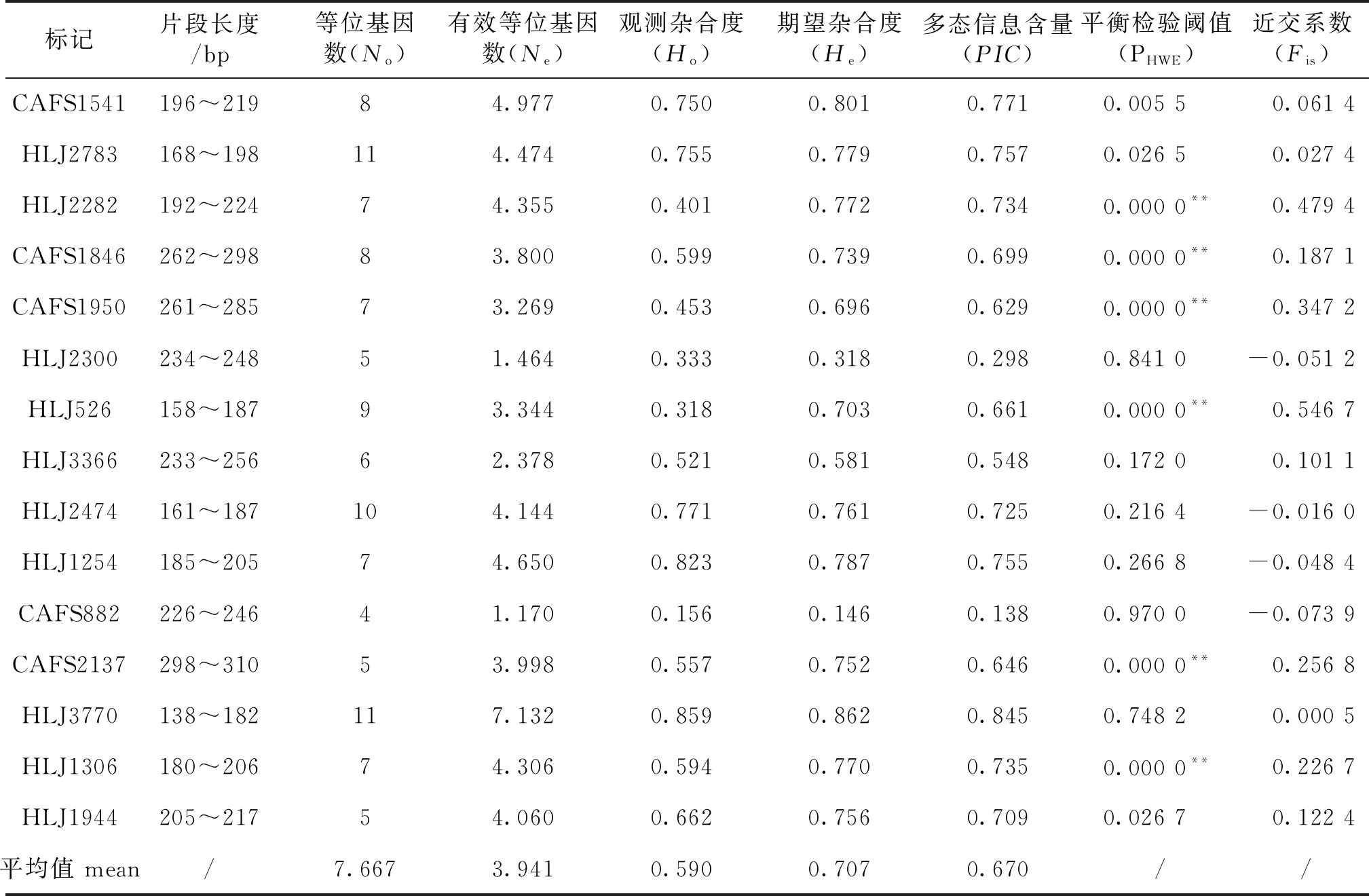
续表2
2.3 亲本群体雌雄个体间遗传距离分布及繁殖组间的遗传分化
用24个QTL标记分析了金边鲤亲本群体雌、雄个体间的遗传距离分布,结果雌、雄个体间遗传距离位于0.1231~1.8563之间,遗传距离呈正态分布,其中峰值位于0.7~0.9之间,占38.36%(图1)。基于个体间遗传距离的NJ聚类图将亲本个体聚为5个大小不等的分支,显示出群体内出现一定程度的遗传分化(图2),结合育种实际将亲本群体划分为3个繁殖组,Group I包含24尾雌鱼和36尾雄鱼;Group II包含34尾雌鱼和50尾雄鱼;Group III包含28尾雌鱼和20尾雄鱼。
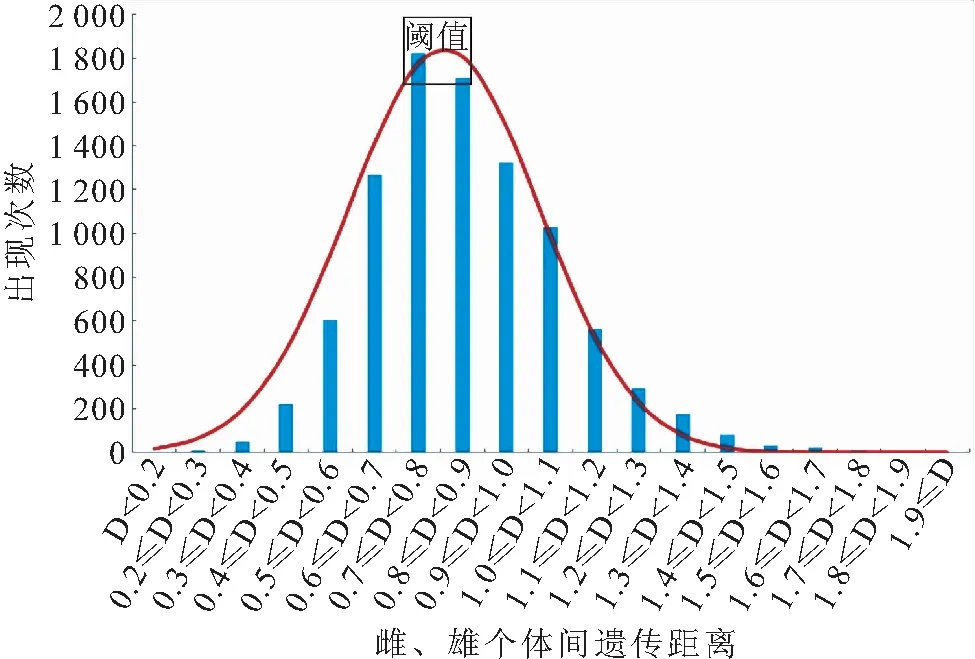
图1 金边鲤亲本群体雌、雄个体间遗传距离分布图
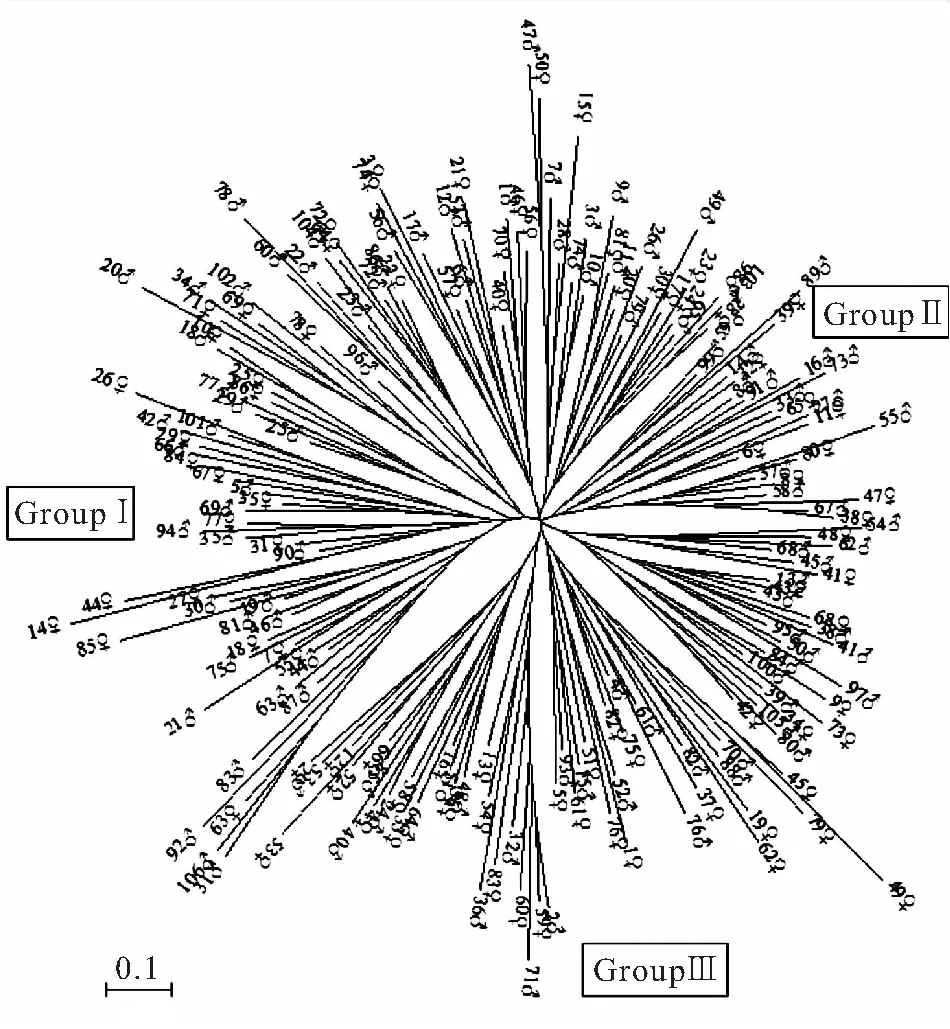
图2 金边鲤亲本群体基于NJ算法的个体间聚类图
对3个繁殖组进行遗传变异分析,结果2.67%的遗传变异来自繁殖组间(表3)。繁殖组间遗传分化系数为0.025 48(Group II和Group III)~0.035 09(Group I和Group III),两两繁殖组间的遗传分化系数均达到了显著水平(P<0.05)。
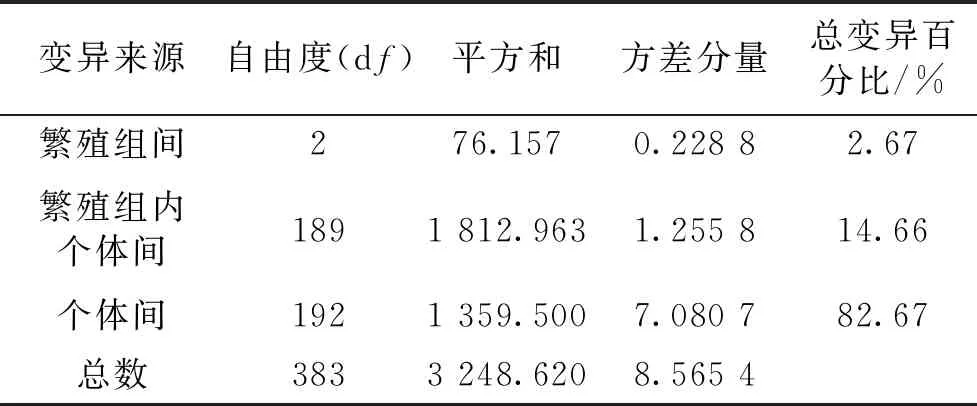
表3 繁殖组间遗传变异的分子方差分析
3 讨论
3.1 关于金边鲤亲鱼群体的遗传多样性问题
本研究用24个与生长性状相关的QTL标记分析金边鲤亲本群体的遗传结构及遗传分化,结果群体平均等位基因数(No=7.667)显著高于有效等位基因数(Ne=3.941),表明群体在生长性状相关的位点还没有显著受到选择育种的影响,仍然保持了丰富的稀有等位基因,在生长性状相关位点尚未显示出明显的富集效应。群体平均观测杂合度(Ho=0.590)显著低于期望杂合度(He=0.707),表明群体已经出现了一定比例的杂合子缺失。Hardy-Weiberg平衡检验近交系数的结果也表明:群体在19个(79.17%)QTL位点出现不同程度的杂合子缺失,9个(37.50%)位点出现极显著的杂合子缺失,显示出人工选择的迹象。群体多态信息含量能够直观地反映出群体遗传多样性水平,按照Botstein等[18]的划分标准,金边鲤亲本群体处于高度多态水平(PIC=0.670>0.5),遗传多样性较好,具备进一步生产及选育优良品种的遗传潜力,同时也体现出金边鲤主要以外形为选育目标,而以生长为目标的选育尚未完成。
潘贤辉等[19]用mtDNA分析的结果表明金边鲤与禾花鲤桂林品系和野鲤分化显著,且遗传多样性低于二者。甘宝江等[20]用10个微卫星标记的分析结果表明:虽然金边鲤群体处于高度多态水平,但是却低于黑龙江和太湖野生鲤群体、福瑞鲤和建鲤等养殖群体,且与两者具有中等程度的遗传分化。本研究用QTL标记计算的金边鲤群体5项遗传参数均低于甘宝江等[20]的分析结果,分析原因一方面QTL标记因与性状紧密连锁而具有较高的保守性,多态性水平相对较低;另一方面群体遗传多样性的分析结果不仅受群体遗传背景的影响,样本量、标记数量、标记多态性水平、标记类型、分型技术等因素也对分析结果具有较大的影响[21,22]。朱华平等[23]的研究发现华南鲤F5的遗传多样性低于地方品种,且随着选育世代的增加,遗传多样性参数呈现逐代下降的趋势[24],其中F5平均等位基因数和多态信息含量较F1分别降低了28.17%和15.36%,认为人工选育加快了华南鲤选育品种与地方品种的遗传分化,也导致了选育品种遗传多样性下降。因此,金边鲤在选育过程中,要特别注意对选育亲鱼群体遗传多样性的保护,以提高选育效率,保持良种的生产性能,延长良种的使用寿命。
3.2 关于亲本间遗传距离分布规律及配组问题
基于分子标记估算的亲本遗传差异指导繁殖配组的育种技术已经开发了成套的选育种软件[25],在多个物种的推广应用中取得了良好的选育效果。面对庞大的基因组资源,如何选择标记用于遗传背景的评估是需要考虑的问题。以往的研究对标记的选择多带有随机性,然而,由于微卫星标记遍布整个基因组,随机选择微卫星标记计算的遗传距离虽然能够反映亲本间的遗传差异,但是却不能反映亲本间在目标性状的遗传差异,而大量与研究目标性状无关的位点也将稀释遗传距离与目标性状间的相关性。
本研究参考周康奇等[26]的研究结果,用鲤体质量、体长、体高和体厚4个性状的QTL标记评估了金边鲤亲本间遗传距离,提高以生长性状为目标的选配效率。结果雌、雄个体间遗传距离呈正态分布,峰值位于0.7~0.9,高于镜鲤和豫选黄河鲤[5,11],低于福瑞鲤和半滑舌鳎[10,11]。综合以上几种鱼类亲本个体间遗传距离的分布发现:个体间遗传距离呈正态分布,与子代生长性状随亲本遗传距离增大先升高后下降的趋势相似;遗传距离的正态分布曲线是变动的,个体间遗传距离的大小不仅取决于群体的遗传背景,同样还受到样本量大小、标记的多态性、标记类型以及检测技术的影响。本研究用毛细管凝胶电泳技术对金边鲤亲本群体进行基因型分析,提高了分型的准确性,也会使群体的遗传多样性高于垂直版聚丙烯酰胺凝胶技术的分析结果;选取峰值临近的2个遗传距离范围作为繁殖阈值切实可行,一方面可以避免近亲交配,另一方面可以最大限度地扩大亲本的可选择性,提高配组效率。
金边鲤的种源(广西融水田鲤)组成复杂[3],加之其选育时间较短,因此群体遗传多样性较高,个体间遗传距离较大。基于遗传距离的聚类图也显示出明显的呈簇分支,依据聚类图划分的繁殖组间的遗传分化系数也达到显著水平,表明群体来源的复杂性和差异性。选择不同遗传距离范围的亲本个体繁殖3个禾花鲤家系进行网箱生长对照实验,结果来源于不同繁殖组且位于遗传距离阈值的家系生产性能最佳[27]。据此,建议金边鲤家系繁殖配组时,宜从不同繁殖组选择个体间遗传距离位于最佳“阈值”0.7~0.9之间的个体作为亲本;群体繁殖时,宜繁殖组间雌、雄交叉配组,有助于提高群体的生产性能、保持群体的遗传多样性水平。
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
智慧健康(2021年17期)2021-07-30 14:38:32
环球人物(2020年13期)2020-07-13 09:33:52
孩子(2020年4期)2020-06-08 10:44:48
中国果业信息(2019年11期)2019-01-05 20:47:24
天然产物研究与开发(2018年6期)2018-07-09 06:01:30
现代检验医学杂志(2016年5期)2016-08-20 03:16:54
西南农业学报(2016年5期)2016-05-17 05:42:23
法医学杂志(2015年4期)2016-01-06 12:36:40
新疆农垦科技(2014年10期)2014-02-28 19:21:08
