
‘满天星’西番莲花芽分化与坐果期生理代谢效应
2021-07-10 01:30黄诚梅罗海斌黄永才魏源文吴兴剑叶丽萍曹辉庆蒋胜理
热带农业科技 2021年3期
黄诚梅,罗海斌,黄永才,魏源文,杨 柳,吴兴剑,叶丽萍,曹辉庆,蒋胜理
(广西作物遗传改良生物技术重点开放实验室,南宁530007)
西番莲(Passiflora edulis)又名百香果、鸡蛋果,属于西番莲科(Passifloraceae)西番莲属(PassifloraL.),主要包括紫果西番莲和黄果西番莲,以及两个品种之间相互杂交得到的系列品种,为热带和亚热带多年生草质到半木质藤本攀缘植物,我国主要种植区域分布在台湾、广西、广东、海南、福建、云南、浙江、四川和贵州等省区[1-3]。西番莲果实营养价值很高,富含有人体所需17 种氨基酸、多种维生素和类胡萝卜素、矿物质元素等,有鲜食、药用、油用和绿色观赏等多种用途,具极高的经济价值[4-5]。
西番莲生长周期短、产量高、效益好、适应性广,具有非常好的商业价值,在我国南方地区多地推广种植,被广西不少区县列为“短、平、快”的精准扶贫项目,在桂林、河池、玉林、柳州、梧州、南宁等地均发展快速,种植面积不断扩大。2017年广西种植面积达2 万hm2,产量25 万t,占全国六成以上;2018 年西番莲种植面积超过2.67 万hm2,产量达34 万t。种植品种以紫色西番莲(台农1 号、紫香)、黄金果(芭乐味、蜂蜜味、荔枝味等)、‘满天星’等为主,其中‘台农1 号’为各地的主栽品种,也引进试种‘小果黄金果’‘大果黄金果’等新品种[3,5]。
西番莲坐果率与气象因子有密切相关性,尤其是温度和湿度[7-8]。在高温季节,高温会不利于授粉受精从而导致花器发育不健全,甚至会出现枯花、落花现象;雨水过度也会导致坐果率的下降,适当的降雨则可以提高坐果率[3,8-11]。台农1 号西番莲在气温超过32℃时花芽出现退化现象,植株不能坐果,广西东部和南部6-9 月处于高温高湿季节,台农1 号花芽分化后发生退化。台农1 号西番莲的单株坐果率不高,平均坐果率为36.5%,尤其在高温高光季节如8 月中下旬和9 月中上旬的单株坐果率均未超过30%[12]。本研究以当前种植品种‘满天星’西番莲为材料,分析在广西高温季节7-9 月份期间其花芽形成与坐果的生理生化代谢变化,如抗氧化物质、抗氧化酶、渗透调节物质以及光合作用与氮代谢关键酶活性的变化,为研究如何在高温胁迫下提高其花芽分化与耐热性等提供科学依据。
1 材料和方法
1.1 试验地点与供试材料
试验于2020 年3 月30 日在广西农业科学院里建科学研究基地进行。供试品种为‘满天星’西番莲,果皮呈紫黄色,外表皮带白色的小斑点。
1.2 试验设计与样品采集
试验采用随机区组设计,种植的西番莲苗为嫁接苗(嫁接砧木为黄果实生苗),苗高25~40 cm,于2020 年3 月30 日移栽大田,采用高垄覆膜与篱架种植模式,行株距为2 m×1.5 m,种植密度为160 株/667m2。移栽基肥、苗期施肥与开花结果期施肥管理,及其田间病虫害管理等,均参照常规田间管理方法,并统一进行枝蔓修剪和管护工作。
田间从8 月中旬开始花芽分化,9 月底到11月初花量较大,达到盛花期,11 月底花量减少。试验从有花芽形成开始,全田选择植株长势比较一致的地块分成3 个小区,每个小区50 株,调查其花芽分化、坐果情况。
于连续3 d 及以上晴朗无雨的天气开始进行样品采集,分别于2020 年7 月17 日(当地气象温度:26~34℃,晴天)、7 月 30 日(当地气象温度:26~35℃,晴天)、8 月19 日(当地气象温度:24~33℃,晴天)、8 月 25 日(当地气象温度:25~34℃,多云转晴)、9 月11 日(当地气象温度:24~32℃,多云转晴)、11 月4 日(当地气象温度:18~25℃,多云转晴),编号:A1、A2、A3、A4、A5、A6,进行样本采集。选取生长发育状态比较一致的3条二级枝蔓,每个处理随机选取生长比较一致的植株3 株,采集倒数展开完整叶1~3 叶,重复3次,液氨速冻后用于生理生化指标测定。
1.3 测定项目与方法
蔗糖含量采用 Ros 比色法测定[13],单位为mg·g-1FW;可溶性蛋白质含量测定采用考马斯亮蓝 G-250 染色法[14],单位为 mg·g-1FW;可溶性淀粉含量用碘显色法[15],单位为 mg·g-1FW;丙二醛(MDA)含量测定采用硫代巴比妥酸显色法[16],单位为nmol·g-1FW。
过氧化物酶(POD)、超氧化物歧化酶(SOD)、a-淀粉酶、蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS 合成方向)、硝酸还原酶(NR)等酶活性均采用苏州格锐思生物有限公司的微板法测试盒测定。POD 酶活性以每克组织每分钟在反应体系中使470 nm 处吸光值增加1 为一个酶活单位U,单位为△OD470·min-1·g-1FW;SOD 酶活性以在黄嘌呤氧化酶藕联反应体系中抑制百分率为50%时,反应体系中的SOD 酶活力定义为一个酶活力单位(U/mL),单位为U·g-1FW;a-淀粉酶活性单位为mg·min-1·g-1FW;SPS 活性单位为 mg·min-1·g-1FW;SS(合成方向)活性单位为mg·min-1·g-1FW;NR 活性单位为nmol·h-1·g-1FW。每个处理测定重复3次,取平均值。
1.4 数据统计分析
数据统计与处理采用DPS v14.10 软件进行统计分析,以Duncan 新复极差法进行多重比较检验差异显著性;采用Microsoft Excel 2007 进行图表绘制。
2 结果与分析
2.1 西番莲枝蔓生长及开花坐果进程
西番莲7 月份二、三级蔓基本已经长成并全部上架,8 月中旬开始出现花芽分化,9 月份开始进入坐果期。根据田间调查数据(图1),由于8 月份温度较高,全田花芽分化率为13.33%,9 月份才达到29.33%,而其坐果率也较低,8 月份才有8%,9 月也仅为23.33%,花量与坐果量也不高,而且出现大量枯花、落花现象。而在10-11 月才达到盛花期,其坐果率才有所提高。

图1 田间西番莲成花与坐果情况
2.2 西番莲枝蔓叶片丙二醛(MDA)和可溶性淀粉含量变化
MDA 是许多植物细胞膜内不饱和脂肪酸发生过氧化作用的终产物,其含量的高低可用于评判植物细胞膜质损伤程度。从图2可 见 ,MDA 含 量 在 8 月 19 日 达 到高峰期,之后持续保持高含量,与7 月份均达到极显著差异。对于淀粉含量,也在同样的反应趋势,均在(8 月19 日)达到高峰期,之后持续保持高含量,与7 月份均达到极显著差异。

图2 西番莲枝蔓叶片MDA和可溶性淀粉含量
2.3 西番莲枝蔓叶片可溶性蛋白质和蔗糖含量变化

图3 西番莲枝蔓叶片可溶性蛋白质和蔗糖含量
从图3 可见,可溶性蛋白与蔗糖含量则随着时间进程有所下降趋势。可溶性蛋白含量在7 月30日开始下降,8 月份则处于低水平时期,9 月份稍有回升,但在11 月份植株坐果期则处于最低含量水平,各时期均达到极显著差异。蔗糖含量也是在8 月份处于低水平时期,各时期均也达到极显著差异。
2.4 西番莲枝蔓叶片蔗糖磷酸合成酶(SPS)与蔗糖合成酶(SS-合成方向)活性
由图4 可知,SPS 活性在7 月底降至最低水平,8 月份稍有提升,之后9 月的花芽分化期与11月的坐果期,其活性稍有下降,结合当时气象信息,高光强与高温天气抑制了西番莲植株叶片光合作用。合成方向的SS 活性则也是在7 月底开始下降,8 月底与11 月则降至最低水平。

图4 西番莲枝蔓叶片SPS和SS(合成方向)活性

图5 西番莲枝蔓叶片POD和SOD活性
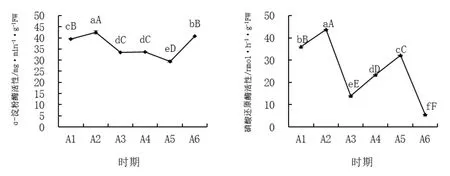
图6 西番莲枝蔓叶片ɑ-淀粉酶和硝酸还原酶活性
2.5 西番莲枝蔓叶片过氧化物酶(POD)与超氧化物歧化酶(SOD)活性
由图5 可知,在高温诱导下,西番莲植株本能启动自身的抗氧化系统,从而达到提高自身抵御高温能力。从8 月开始,其POD 和SOD 活性开始提高,尤其是SOD 活性在8 月25 日显著提升至高水平。而后9 月11 日两者则有所下降,11 月4 日再次提高,POD 活性则是达到高水平。
2.6 西番莲枝蔓叶片a-淀粉酶与硝酸还原酶活性
从图6 可知,ɑ-淀粉酶活性在8、9 月份处于低谷,而后在11 月结果期才有所上升。而硝酸还原酶活性呈Z 型变化规律,在8 月19 日和11 月4日均处于低水平,尤其以坐果期11 月4 日最低,各时期达到极显著差异。
3 讨论
苹果、梨等一些落叶果树,营养物质的积累是花芽分化的物质基础,营养物质是否充足决定了花芽分化质量的优劣[17]。然而,百香果的花芽形成和枝条生长是同时进行的,花芽形成和茎叶生长同步必然存在养分的竞争,其花芽形成有其自身的生物学特性和规律,但也受器官间营养分配和外界环境的影响[12]。许多果树 植 物 如 柑 橘[18]、苹 果[19]、葡萄[20]、枣树[21]、梨树[22]等在受到高温胁迫时,其内部生理生化反应和相关基因、蛋白表达都会发生一系列变化:光合作用下降,呼吸作用减弱,细胞膜透性增大,活性氧含量增加,启动抗氧化物质与抗氧化酶等发生应答性变化。
可溶性淀粉作为植物体内重要的贮存物质,本研究中的西番莲植株叶片淀粉含量在8 月份后均能保持较高水平,其大量贮存为西番莲花芽分化等提供了所需要的能量与结构物质。而其α-淀粉酶活性在8-9 月高温期间则是保持较低活性,到11 月4 日的坐果期才提高至高水平,这说明了α-淀粉酶在高温季节保持低活性有利于淀粉的积累,而在后期其活性提高将有利于淀粉的分解成为花芽生长发育所需的营养物质,有利于西番莲植株花芽分化与坐果。可溶性蛋白质是植物生长发育所需的重要有机营养物,其含量的高低反映着植物生理生化代谢能力的强弱[23]。本研究中,西番莲植株叶片中可溶性蛋白含量随时间进程有所下降,其可能是植株随着花芽分化与坐果过程中过度消耗营养物质,这为加强西番莲花芽分化坐果期田间管理措施提供一定的参考。
蔗糖是光合产物代谢最直接的物质。蔗糖磷酸合成酶(sucrose phosphatesynthase, SPS)是以UDPG 为供体,以6-磷酸果糖为受体的蔗糖合成调节关键酶,催化UDP-葡萄糖(UDPG)和果糖-6-磷酸(F6P)转化为UDP 和蔗糖-6-磷酸(S6P),随后通过蔗糖磷酸酶(SPP)水解形成蔗糖[24,25]。SS存在于细胞质中,SS 催化UDP-葡萄糖和果糖生成蔗糖和二磷酸尿苷(UDP),催化的反应是可逆的,当SS 分解方向起作用时,分解蔗糖生成UDPG和果糖,SS 合成方向有利于蔗糖的形成[25-26]。高温明显降低 SPS 和 SS 合成方向活性[27]。本研究中,高温季节均降低了蔗糖合成相关酶活性,从而降低蔗糖含量,而随着花芽分化和坐果进程,其SPS 和SS 活性均有所下降,这是否与植株营养分配均衡性有关,有待于后续研究。而对于硝酸还原酶活性呈Z 型变化规律,在8 月19 日和11 月4 日均处于低水平,尤其以坐果期11 月4 日最低,这可能与植株在后期氮利用效率有一定关系,高温对其影响呈无规律变化。硝酸还原酶作为植物利用硝酸盐的关键酶之一,硝酸盐还原在植株根内和枝叶中均可以完成,NR 活性常用来表示氮代谢的水平[28-29]。
热胁迫会引起植物叶片产生的大量活性氧从而引起膜脂过氧化,导致大量过氧化产物MDA 累积,对植物光合系统和抗氧化系统造成损伤[30-31]。POD、SOD 是清除自由基、防止膜脂过氧化的重要酶类,可以保持自由基产生和消除之间平衡[32-36]。徐超等[34]研究发现,高温处理后草莓幼苗具有较高的MDA 含量,其抗氧化酶活性增加,但随着时间的延长,抗氧化酶活性又显著降低。本研究中,西番莲植株叶片中MDA 含量在8月19 日达到高峰期,之后持续保持高含量;其POD 和 SOD 活性自 8 月开始提高,尤其是 SOD 活性,这可能是在高温诱导下西番莲植株本能启动自身的抗氧化系统,从而达到提高自身抵御高温能力。
4 结论
西番莲品种‘满天星’容易受到高温等因素影响,花芽分化期延迟,直至10-11 月份才进入盛花期和盛果期。在高温季节,西番莲植株通过启动POD 和SOD 活性等自身抗氧化系统,及提高可溶性淀粉贮藏为其花芽分化等提供了所需要的能量与结构物质,而蔗糖与可溶性蛋白质等则有所下降;随着花芽分化和坐果进程,其SPS 和SS(合成方向)活性均下降,氮代谢也受到不同程度上的抑制。下一步将对其它西番莲品种开展相关研究,进一步分析其避热和耐热反应机制。
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23
中国果业信息(2022年4期)2022-11-22
农产品加工(2022年14期)2022-08-18
热带作物学报(2022年5期)2022-06-01
种子科技(2021年11期)2021-08-19
VOGUE服饰与美容(2021年5期)2021-05-13
果农之友(2020年5期)2020-06-21
农民致富之友(2020年6期)2020-04-08
农业与技术(2019年5期)2019-05-29
农民致富之友(2018年21期)2018-12-27
