
Tagitinin F诱导MDA-MB-231细胞凋亡作用机制研究
2021-07-03 06:37王娟杨旭赵越勤丁骁夏海滨赵逾涵陕西师范大学生命科学学院基因治疗重点实验室西安710119中国科学院昆明植物研究所植物化学与西部植物资源持续利用国家重点实验室昆明65001
中南药学 2021年5期
王娟,杨旭,赵越勤,丁骁,夏海滨,赵逾涵*(1.陕西师范大学生命科学学院 基因治疗重点实验室,西安 710119;.中国科学院昆明植物研究所 植物化学与西部植物资源持续利用国家重点实验室,昆明 65001)
乳腺癌(breast cancer)是女性中常见的恶性肿瘤,是女性癌症死亡的第二大主要原因[1]。目前乳腺癌的治疗方法主要包括显微外科切除术,放射疗法和化学疗法[2]。在乳腺癌中,三阴性乳腺癌(triple negative breast cancer,TNBC)通常是指缺乏雌激素受体(estrogen receptor,ER),孕激素受体(progesterone receptor,PR)和人表皮生长因子受体2(human epidermal growth factor receptor-2,HER2)表达的乳腺癌,占所有乳腺癌的15%~20%,更容易转移和浸润,恶性程度更高,容易产生化学耐药性,因此目前TNBC的标准治疗方法主要是化学疗法和放射疗法。但是,尽管经过标准化治疗,患者的平均总生存期仅为12~18个月[3-4]。因此寻找更多对TNBC有效的药物一直是世界范围内学者研究的热点。
Tagitinin F(结构式见图1)属于倍半萜内酯类天然小分子化合物,在菊科植物肿柄菊中大量存在[5],其具有抗炎、抗微生物、抗寄生虫、抗肿瘤[5-6]等活性,其类似物tagitinin C能够诱导肿瘤细胞caspase依赖性凋亡,甚至导致肿瘤细胞自噬死亡,而且tagitinin C衍生物对治疗癌症具有巨大潜力[7]。但是对tagitinin F的抗肿瘤活性的具体作用机制研究较少,故本研究拟对其机制进行探讨。
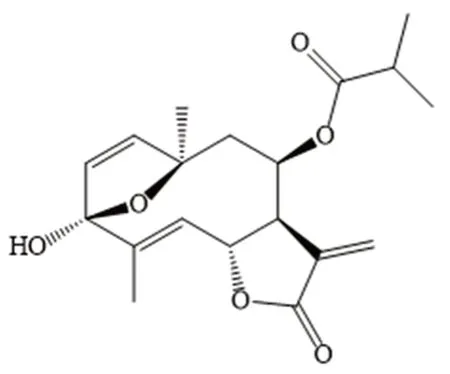
图1 Tagitinin F的化学结构Fig 1 Chemical structure of tagitinin F
1 材料
1.1 细胞来源
人乳腺癌MDA-MB-231细胞株由中国科学院昆明植物研究所活性筛选中心孔清华老师惠赠。
1.2 仪器
成像型微孔板检测仪(BioTek Cytationl),全自动化学发光图像分析系统(tanon,5200),流式细胞仪(BD FACSCalibur Flow Cytometer),CO2培养箱(Thermo Scientific),生物显微镜(Olympus,CKX31),血球计数板(上海市求精生化试剂仪器有限公司),电泳仪(Bio-Rad),冷冻离心机(Eppendorf,5415R)等。
1.3 试药
Tagitinin F(云南西力生物技术有限公司,纯度:98%,货号:BBP00166),紫杉醇(Meilunbio,货号:MB1178),衣霉素(Abcam,货号:ab120296),碘化丙啶(PI,BioFroxx公司,货号:1246),3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴盐(MTT,BioFroxx公司,货号:3580),Annexin V-FITC/PI双染细胞凋亡检测试剂盒(四正柏生物,货号:FXP018),胰蛋白酶(Biological Industries,货号:03-050-1A),100×青链霉素混合液(Biosharp,青霉素10 000 U·mL-1、链霉素10 000 μg·mL-1,货号:P1400),RNase A溶液(Solarbio)无水乙醇,Bip、PDI、Calnexin、Ero1-Lα、IRE1α抗体(Cell Signaling Technology,货号:9956T),β-actin(Cell Signaling Technology,货号:3700S)。
2 方法
2.1 细胞培养
将细胞培养于含10% FBS(胎牛血清),1%双抗的DMEM高糖培养基中,置于37℃、5%CO2细胞培养箱中,取处于对数生长期且状态良好的细胞用于后续实验。
2.2 MTT法检测人乳腺癌MDA-MB-231细胞活力
取对数生长期的MDA-MB-231细胞接种于96孔板中。待细胞贴壁后,加入不同浓度的用DMSO配制的tagitinin F(1.25、2.5、5、10、20 μmol·L-1)或TAXOL(0.05 μmol·L-1)继续培养72 h,弃上清液,每孔加入100 μL的MTT溶液(1 mg·mL-1),孵育4 h,每孔加入100 μL DMSO,用成像型微孔板检测仪在490 nm波长下测量各孔的吸光度(OD)值,计算细胞存活率。
2.3 Tagitinin F对细胞形态影响的观察
将细胞消化后接种于6孔板中,待细胞贴壁后,加入不同浓度的tagitinin F(5、10、20 μmol·L-1),设DMSO(0.1%)对照组,随后将6孔板放置于Incucyte S3自带培养箱中继续培养48 h,每隔12 h拍照记录细胞的形态。
2.4 克隆形成实验
取处于对数生长期的MDA-MB-231细胞接种于12孔板中。待细胞贴壁后,加入不同浓度的tagitinin F(1、2 μmol·L-1),紫杉醇(0.02 μmol·L-1)以及0.1%的DMSO处理细胞,继续培养12 d,弃去培养基,每孔加入500 μL考马斯亮蓝R250(2.5 mg·mL-1)染色2 h,弃去染色液,PBS清洗3遍,拍照。
2.5 流式细胞仪检测细胞周期
将细胞消化后接种于6孔板中,待细胞贴壁后,加入不同浓度的tagitinin F(5、10 μmol·L-1)、紫杉醇(0.05 μmol·L-1)以及0.1%的DMSO处理24 h。收集细胞,加入70%乙醇在-20℃过夜固定。加入2.5 μL RNase A(10 mg·mL-1),使其终质量浓度为50 μg·mL-1,37℃孵育30 min,再加入25 μL PI在室温下孵育30 min后过滤,流式细胞仪检测细胞周期。
2.6 流式细胞仪检测细胞凋亡
将细胞消化后接种于6孔板中,待细胞贴壁后,加 入20 μmol·L-1的tagitinin F以 及0.1%的DMSO分别处理不同时间(24、48、72 h),收集细胞,按凋亡试剂盒说明书分别加入试剂,流式细胞仪检测细胞凋亡率。
2.7 DCFH-DA探针检测细胞内活性氧变化
将细胞消化后接种于6孔板中,待细胞贴壁后,加入不同浓度的tagitinin F(5、10、20 μmol·L-1)、H2O2(2 mmol·L-1)以及0.1%的DMSO处理12 h,弃培养基,加入无血清培养基配制的DCFH-DA(20 μmol·L-1),室温孵育15 min,弃去培养基,PBS清洗3遍,拍照,收集细胞用流式细胞仪检测荧光强度。
2.8 Western blot检测内质网应激相关蛋白表达
将细胞消化后接种于6孔板中,待细胞贴壁后,加入5 μmol·L-1的tagitinin F、2 μmol·L-1的衣霉素以及0.1%DMSO分别处理细胞24 h,收集细胞制备蛋白样品,10%聚丙烯酰胺-SDS凝胶电泳后转至PVDF膜上,5%脱脂奶粉室温封闭1 h,加入一抗4℃孵育过夜,PBST洗3次,加入二抗室温孵育1 h,PBST洗3次。加入ECL化学发光液后放入全自动化学发光图像分析系统中曝光。
2.9 统计学分析
每组实验独立重复3次,用Graph-Pad Prism 8软件进行统计分析处理,结果以均值±标准差表示,用t检验进行组间差异分析,P<0.05表示差异具有统计学意义。
3 结果与分析
3.1 Tagitinin F对人乳腺癌MDA-MB-231细胞活力的影响
如图2所示,将DMSO组的细胞存活率定义为100%,与DMSO组相比,tagitinin F不同浓度处理组细胞存活率均降低(P<0.05,P<0.001),说明tagitinin F对MDA-MB-231细胞的活力有明显的抑制作用,IC50为(5.07±0.19)μmol·L-1。

图2 Tagitinin F对MDA-MB-231细胞活力的抑制作用Fig 2 Inhibit viability on MDA-MB-231 cells of tagitinin F
3.2 Tagitinin F对人乳腺癌MDA-MB-231细胞形态的影响
如图3所示,对照组DMSO处理的细胞贴壁良好,连接紧密,细胞呈梭形且边缘清晰,间质饱满。而tagitinin F处理后,随着药物浓度以及处理时间的增加,细胞贴壁能力变弱,体积变小,细胞之间的间隙逐渐增大,细胞固缩变圆,漂浮细胞增多,失去正常细胞的形态。说明tagitinin F能够改变细胞形态并促进细胞凋亡。

图3 Tagitinin F对MDA-MB-231细胞形态的影响Fig 3 Effect of tagitinin F on the morphology of MDA-MB-231 cells
3.3 Tagitinin F对人乳腺癌MDA-MB-231细胞体外克隆形成能力的影响
如图4A所示,与DMSO组相比,不同浓度的tagitinin F(1.2 μmol·L-1)以及紫杉醇(0.02 μmol·L-1)作用于MDA-MB-231细胞12 d后,克隆球数量以及大小都明显降低(见图4B),说明tagitinin F能够浓度依赖性地抑制MDAMB-231细胞的体外克隆形成能力。
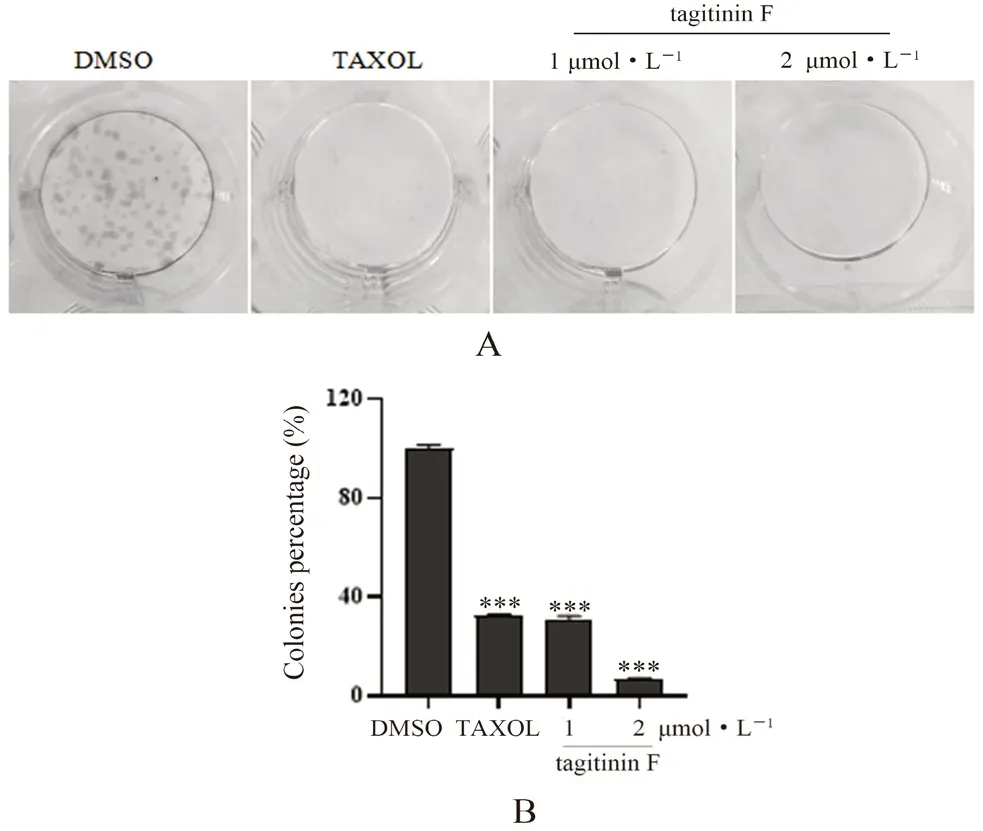
图4 Tagitinin F对MDA-MB-231细胞体外克隆形成能力的抑制作用Fig 4 Inhibition of tagitinin F on the colony formation of MDAMB-231 cells
3.4 Tagitinin F对人乳腺癌MDA-MB-231细胞周期的影响
如图5A所示,随着tagitinin F处理浓度的增加,细胞的G0/G1期比例由43.2%下降到34.7%,S期比例由37.3%下降到6.01%,G2/M期比例由19.5%升高到59.2%。随着处理浓度的增加,处于G2/M期的细胞也随之增多,与DMSO组(19.68%)相比,差异有统计学意义(见图5B),说明tagitinin F能够浓度依赖性地将MDA-MB-231细胞周期阻滞在G2/M期。

图5 Tagitinin F对MDA-MB-231细胞G2/M期的影响Fig 5 Effect of tagitinin F on cell cycle of MDA-MB-231 cells
3.5 Tagitinin F对人乳腺癌MDA-MB-231细胞凋亡的影响
如图6A所示,随着tagitinin F处理时间的增加,细胞凋亡率(早期凋亡+晚期凋亡)增加,分别为7.53%、23.86%、57.2%,与DMSO组(4.81%)相比,差异有统计学意义(见图6B)。说明tagitinin F诱导人乳腺癌MDA-MB-231细胞凋亡具有时间依赖性。

图6 Tagitinin F对MDA-MB-231细胞凋亡的影响Fig 6 Effect of tagitinin F on the apoptosis of MDA-MB-231 cells
3.6 Tagitinin F对人乳腺癌MDA-MB-231细胞内活性氧的影响
如图7所示,H2O2作为阳性对照,与DMSO组相比,随着Tagitinin F处理浓度的增加,成像型微孔板检测仪检测荧光强度(见图7A)以及流式细胞仪检测平均荧光强度(见图7B)均逐渐增强;当tagitinin F与活性氧清除剂NAC联用处理后,荧光强度以及平均荧光强度均减弱。说明tagitinin F能显著引起细胞内源性活性氧的积累。
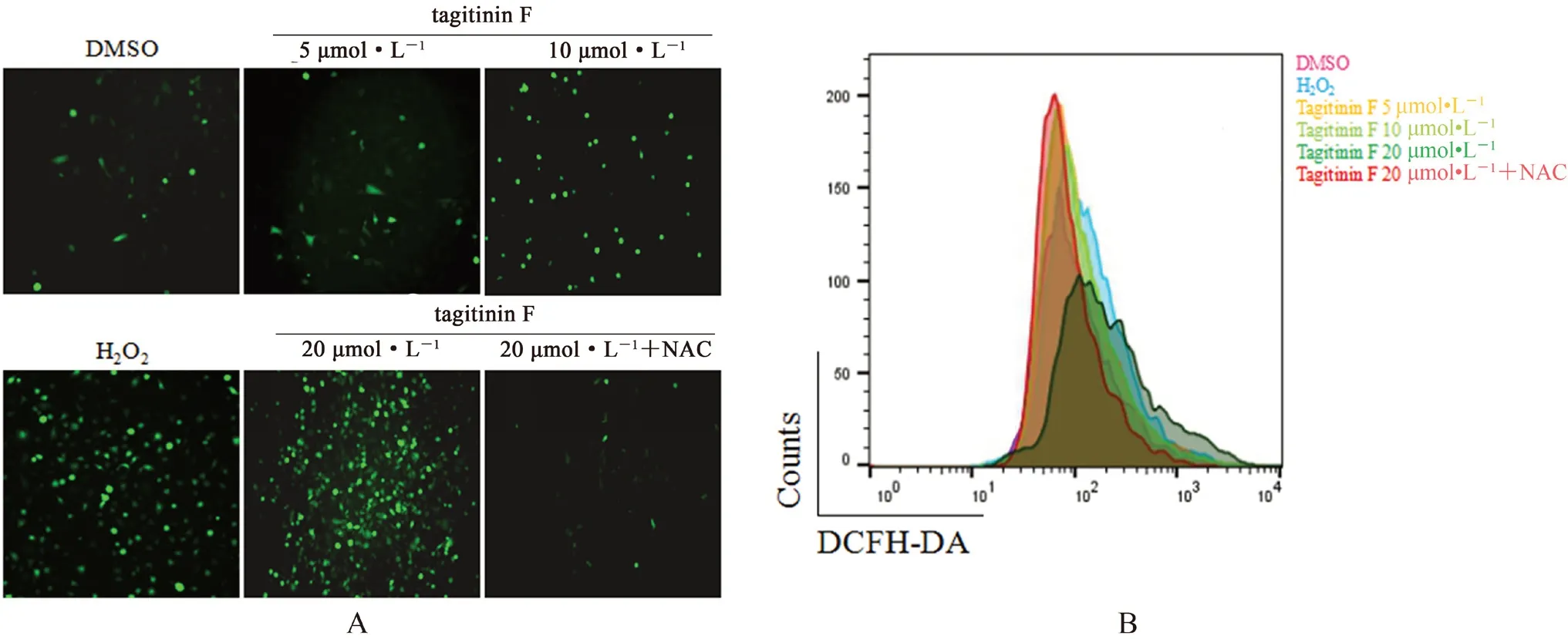
图7 Tagitinin F对MDA-MB-231细胞内活性氧的影响Fig 7 Effect of tagitinin F on the endogenous ROS levels in MDA-MB-231 cells
3.7 Tagitinin F对人乳腺癌MDA-MB-231细胞相关内质网应激信号通路蛋白的影响
如图8所示,衣霉素作为阳性对照化合物,当tagitinin F处理浓度为5 μmol·L-1时,随着处理时间的增加,内质网应激相关蛋白Bip、PDI、Calnexin、Ero1-Lα、IRE1α的表达量升高。说明tagitinin F能够激活内质网应激信号通路,而当内质网应激持续激活时,细胞程序性凋亡。
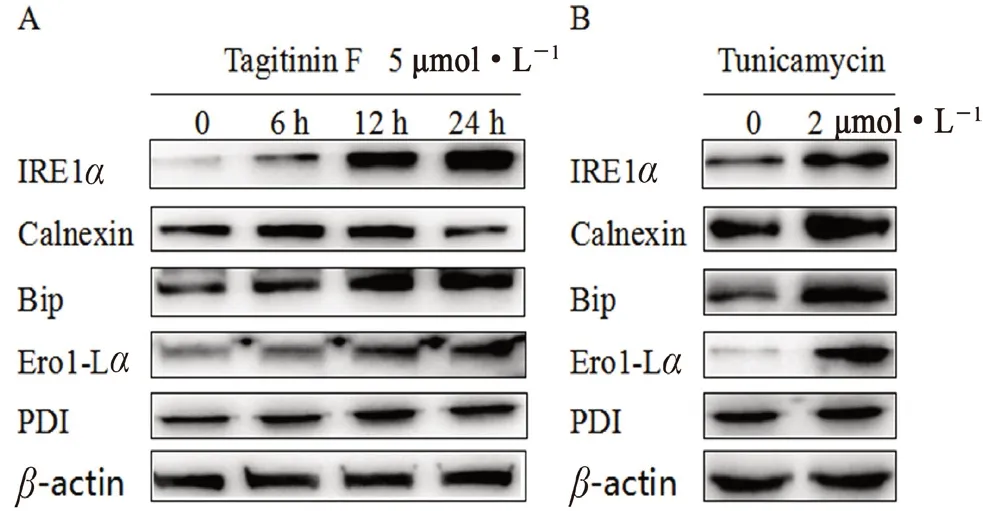
图8 Tagitinin F(A)及衣霉素对内质网应激(B)的影响Fig 8 Effect of tagitinin F(A)and tunicamycin(B)on endoplasmic reticulum stress
4 结论与讨论
乳腺癌是女性中普遍存在的癌症类型,并且是女性癌症死亡的第二大主要原因[1]。而其中TNBC又非常容易转移并产生化学耐药性,所以其临床预后通常较差[4]。由于抗肿瘤药物的局限性,迫切需要从不同的方向寻找更多的化疗药物。从天然植物中提取分离得到的小分子化合物因其来源丰富,结构种类繁多,具有更多的生物活性,毒副作用较小,是抗肿瘤新药研发的重要来源。本文主要从细胞活力、细胞体外克隆形成能力,诱导细胞周期阻滞和凋亡以及相关内质网应激信号通路的影响来研究天然小分子化合物tagitinin F对人乳腺癌MDA-MB-231细胞增殖、诱导凋亡作用及其机制。
本研究中,首先采用了MTT法检测tagitinin F对人乳腺癌MDA-MB-231细胞活力的影响,发现与DMSO组相比,随着tagitinin F浓度的增加,细胞活力也随之下降;细胞形态表明,tagitinin F能够改变细胞形态并杀死细胞;通过克隆形成实验发现,与DMSO组相比,随着处理浓度的增加,克隆球的大小及数量减少;流式细胞仪检测表明,与DMSO组相比,处理组的G2/M期细胞比例增加,细胞的早期凋亡以及晚期凋亡数目都增加,以上结果表明tagitinin F能显著抑制MDA-MB-231细胞增殖,并且诱导其细胞周期阻滞和凋亡。
内质网是一种多功能细胞器,参与许多包括新生的蛋白质折叠和修饰,钙储存,液体生物合成和排毒等细胞生物学过程。细胞在诸如缺氧、营养缺乏、药物诱导的毒性,遗传突变等各种细胞内外的刺激下,会导致内质网内腔中蛋白质的积累或者折叠错误,此时,肿瘤细胞会触发内质网应激来重建细胞内稳态并促进细胞存活[8]。内质网应激与活性氧的产生有关,而且一旦这种应激状态时间过长会激活内质网凋亡信号诱发细胞凋亡[9]。而在本文中,我们用 DCFH-DA探针检测活性氧的变化,经过tagitinin F处理后,细胞内活性氧增加;Western blot结果显示,内质网应激相关蛋白Bip、PDI、Calnexin、Ero1-Lα、IRE1α的表达量升高。说明tagitinin F能够通过使活性氧累积激活内质网应激信号通路发挥作用。
综上所述,tagitinin F能够抑制人TNBC MDA-MB-231细胞的增殖以及体外克隆形成能力,改变其正常的细胞形态;诱导细胞周期阻滞在G2/M期并诱导细胞凋亡;同时使细胞内活性氧水平增加来激活内质网应激信号通路发挥抑肿瘤作用。
猜你喜欢
核科学与工程(2022年3期)2022-10-18
热力发电(2022年3期)2022-03-25
数学物理学报(2021年6期)2021-12-21
能源工程(2021年2期)2021-07-21
服饰导报·鞋世界(2021年4期)2021-05-17
广州化工(2020年6期)2020-04-18
中国药理学通报(2019年5期)2019-01-11
现代矿业(2018年9期)2018-10-16
西南军医(2015年4期)2015-01-23
天然气与石油(2014年2期)2014-07-16
