
NaCl 胁迫对转BADH- 反义4CL 基因二色胡枝子盆栽苗的影响
2021-06-29 01:55解小娟
花卉 2021年12期
解小娟
(山西省投资咨询和发展规划院,山西 太原 030009)
二色胡枝子(Lespedeza bicolor)是豆科胡枝子属(Lespedeza)多年生落叶灌木,具有生物量大、枝叶营养丰富,耐旱、耐寒、耐瘠薄、萌蘖能力强等优良特性,是荒坡绿化、矿山生态修复的先锋树种,同时也是优良的木本饲料作物、园林观赏植物和药用植物。尽管二色胡枝子具有较多优点,但仍然存在抗盐碱能力差、木质素含量高等缺点。目前,应用基因工程技术提高植物的抗盐碱能力、控制木质素的合成已在多种植物中取得成功[1]。甜菜碱被认为是最好的渗透调物质,甜菜碱醛脱氢酶(BADH)是甜菜碱合成的关键酶之一,转入BADH 的美丽胡枝子耐盐能力提高[2-3]。4-香豆酸辅酶A 连接酶(4CL)是控制木质素合成的关键性酶,可以利用不同底物来调控木质素的合成,转入反义4CL 基因的杨树木质素含量降低。本试验以转入BADH、反义4CL 基因的二色胡枝子为材料,研究不同浓度的氯化钠处理对其生理指标的影响,以探讨外源基因的导入对植物渗透调节的影响。
1 材料与方法
1.1 试验材料
试验材料为同时炼苗出瓶的非转基因二色胡枝子,以及分子检测呈阳性的转BADH-反义4CL 基因二色胡枝子的2 个株系(67号、85号)。
所有试验材料经12个月盆栽,苗木高达80~90cm时进行NaCl 胁迫。NaCl 处理浓度分别为0、0.5g/L、1.0g/L、2.0g/L,每种材料处理8 盆,每盆浇灌处理液300mL,隔4d浇灌一次,共3 次,12d后每种材料每个梯度挑选生长一致的叶片进行各项生理指标的测定。
1.2 试验方法
BADH 活性和甜菜碱含量的测定方法:BADH 活性测定参照罗晓丽等的方法。甜菜碱含量测定采用雷氏盐紫外分光光度计法[4]。
相关生理指标测定方法:细胞膜透性采用电导仪法[5]。丙二醛(MDA)含量采用硫代巴比妥酸法[6]。脯氨酸含量测定采用的酸性茚三酮比色法。可溶性蛋白含量测定采用考马斯亮蓝法[6]。抗氧化酶活性的测定根据李合生法[6]。
每种指标重复测定3 次,试验结果用DPS 软件进行分析。
2 结果与分析
2.1 盐胁迫对BADH 活性和甜菜碱含量的影响
为了进一步分析外源基因(BADH)的功能,对转基因二色胡枝子2 个阳性株系进行不同梯度的盐胁迫处理,同时以非转基因二色胡枝子为对照,测定盐胁迫条件下植株的BADH 活性和甜菜碱含量(见图1、图2)。结果表明:随着盐浓度的提高,BADH活性和甜菜碱含量都有所提高。无盐胁迫时,转基因株系与对照植株BADH 活性和甜菜碱含量含量较低,且不存在显著性差异;NaCl 浓度为2g/L 时,转基因株系67号BADH 活性和甜菜碱含量是对照植株的2.5 倍和1.61 倍,转基因株系85号BADH 活性和甜菜碱含量是对照植株的3 倍和1.66 倍,且转基因株系与非转基因植株存在显著性差异。同时BADH 活性增加的幅度高于甜菜碱含量增加幅度。因而,盐胁迫条件下转基因二色胡枝子具有较强的渗透调节能力,降低了盐胁迫对叶片细胞造成的伤害,抗性增强。

图1 不同NaCl 浓度下BADH 的活性变化
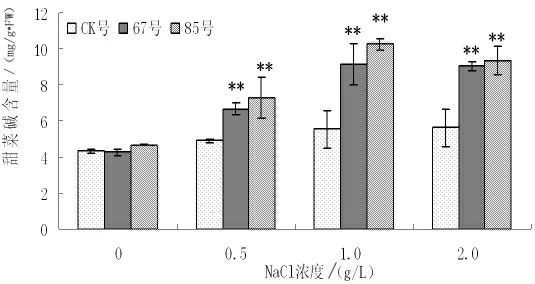
图2 不同NaCl 浓度下甜菜碱含量的变化
2.2 盐胁迫对细胞膜透性和丙二醛含量的影响
盐胁迫对非转基因和转基因二色胡枝子膜透性的影响如图3所示,随着盐浓度的增加,非转基因和转基因二色胡枝子相对电解质渗透率(EL)呈上升趋势。无盐胁迫条件下,非转基因和转基因二色胡枝子相对电解质渗透率含量较低,且不存在显著性差异;NaCl 浓度为0.5g/L 时,与无盐胁迫时相比,非转基因植株的EL 增加1.67 倍,转基因株系67号和85号分别增加0.76 倍和1.08 倍,且转基因株系和非转基因植株存在显著性差异;NaCl 浓度为1.0g/L 和2.0g/L,转基因株系的EL 增加的幅度均小于非转基因植株。结果表明盐胁迫条件下转基因二色胡枝子EL 比非转基因植株低,说明转基因植株膜结构完整性较好,膜受到伤害较小,抗盐性强,说明外源基因(BADH)导入提高了植株的抗盐性。盐胁迫对非转基因和转基因二色胡枝子丙二醛(MDA)含量的影响如图4所示,随着盐浓度的增加,非转基因和转基因二色胡枝子MDA 含量与电导率趋势的一致。无盐胁迫条件下,转基因株系和非转基因植株MDA 含量较低,且不存在显著差异;NaCl 浓度为0.5g/L 时,与无盐胁迫时相比,非转基因植株的MDA 含量增加45%,转基因株系67号和85号分别增加13%和9%,且转基因株系和非转基因植株存在显著性差异;NaCl 浓度为1.0g/L和2.0g/L,转基因株系的MDA 含量增加的幅度均小于非转基因植株。结果表明盐胁迫条件下转基因二色胡枝子MDA 含量比非转基因植株低,说明转基因植株膜脂过氧化程度较低,膜结构伤害的程度较小,进一步说明外源基因(BADH)导入提高了植株的抗盐性。

图3 不同NaCl 浓度下电导率的变化

图4 不同NaCl 浓度下丙二醛的变化
2.3 盐胁迫对脯氨酸含量的影响
盐胁迫对非转基因和转基因二色胡枝子脯氨酸(Pro)含量的影响如图5所示,随着盐浓度的增加,非转基因和转基因二色胡枝子Pro 含量呈现先上升后下降的趋势。无盐胁迫时,非转基因植株和转基因各株系间的脯氨酸含量较低,且不存在差异显著;NaCl 浓度为0.5g/L 时,与无盐胁迫时相比,非转基因植株的Pro含量增加0.48 倍,转基因株系67号和85号分别增加2.97 倍和2.47 倍,且转基因株系和非转基因植株存在显著性差异;NaCl 浓度为1.0g/L 时,转基因株系的Pro 含量增加的幅度仍大于非转基因植株;非NaCl 浓度为2.0g/L 时,脯氨酸含量下降,但转基因株系的Pro 含量仍高于非转基因植株。结果表明,盐胁迫条件下转基因二色胡枝子的Pro 含量高于非转基因植株,说明盐胁迫下转BADH 二色胡枝子渗透调节能力较强。
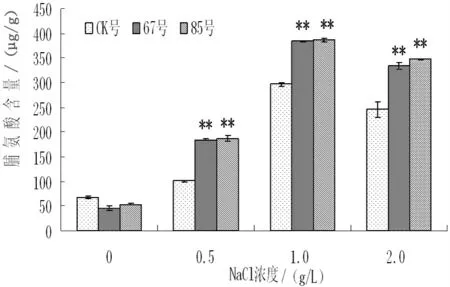
图5 不同NaCl 浓度下脯氨酸含量的变化
2.4 盐胁迫对可溶性蛋白含量的影响
盐胁迫对非转基因和转基因二色胡枝子可溶性蛋白含量的影响如图6所示,随着盐浓度的增加,非转基因和转基因二色胡枝子可溶性蛋白含量呈现下降的趋势。无盐胁迫时,非转基因植株和转基因各株系间的可溶性蛋白含量较高,且不存在差异显著;NaCl 浓度为0.5g/L 时,与无盐胁迫时相比,非转基因植株的可溶性蛋白含量降低了30.4%,转基因株系67号和85号分别降低了17.9%和14.4%,且转基因株系和非转基因植株存在显著性差异;NaCl 浓度为1.0g/L 和2.0g/L 时,转基因株系可溶性蛋白的含量降低的幅度仍小于非转基因植株。结果表明,盐胁迫条件转BADH 二色胡枝子可溶性蛋白含量高于非转基因植株,说明在盐胁迫下转基因二色胡枝子比非转基因植株蛋白质分解速度慢、抗性强。
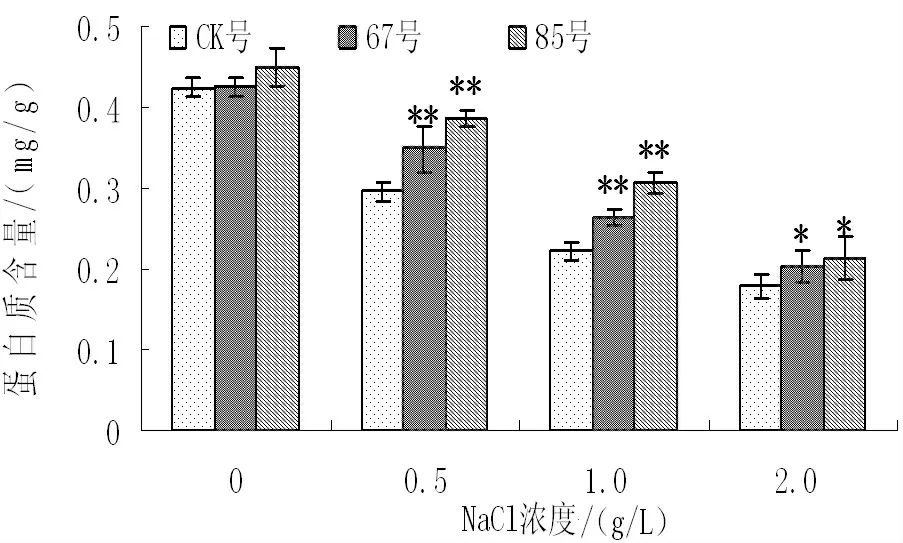
图6 不同NaCl 浓度下可溶性蛋白的变化
2.5 盐胁迫对抗氧化酶活性的影响
盐胁迫对非转基因和转基因二色胡枝子过氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性的影响如图7~图9 所示,随着盐浓度的增加,非转基因和转基因二色胡枝子SOd活性呈现上升的趋势,POD、CAT 活性呈现先上升后下降的趋势。无盐胁迫时,非转基因植株和转基因株系的SOD、POD、CAT 活性较低,且不存在显著性差异;NaCl 浓度为0.5g/L 时,非转基因植株和转基因株系的SOd活性增强幅度较小,67号与非转基因植株不存在显著性差异,85号与非转基因植株存在显著差异,非转基因植株和转基因株系的POd活性也增强幅度较小,且转基因株系和非转基因植株不存在显著性差异,而非转基因植株和转基因株系的CAT 活性也增强幅度较大,转基因株系与非转基因株系存在显著性差异;NaCl 浓度为1.0g/L 时,与非转基因植株相比,非转基因植株的SOd活性增强0.56 倍,转基因株系67号和85号分别增强0.99 倍和0.83 倍,非转基因植株的POd活性增强0.89 倍,转基因株系67号和85号分别增强2.21倍和2.06 倍,非转基因植株的CAT 活性增强1.35 倍,转基因株系67号和85号分别增强1.61 倍和1.74 倍;NaCl 浓度为2.0g/L时,非转基因植株和转基因株系的SOd活性仍呈增强趋势,且转基因株系SOd活性增加的幅度大于非转基因植株,而非转基因植株和转基因株系的POd和CAT 活性却降低,但转基因株系的POd和CAT 活性仍高于非转基因植株。结果表明,盐胁迫条件下转BADH 基因二色胡枝子的抗氧化酶活性高于非转基因植株,说明盐胁迫下转基因二色胡枝子抗氧化酶活性维持在较高水平,其抗性较强。
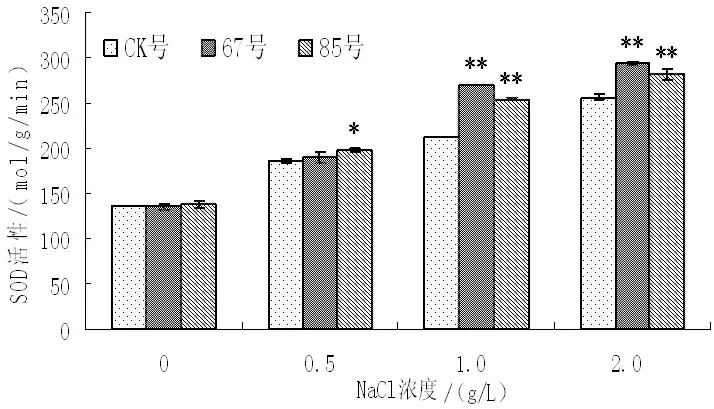
图7 不同NaCl 浓度下SOd活性的变化
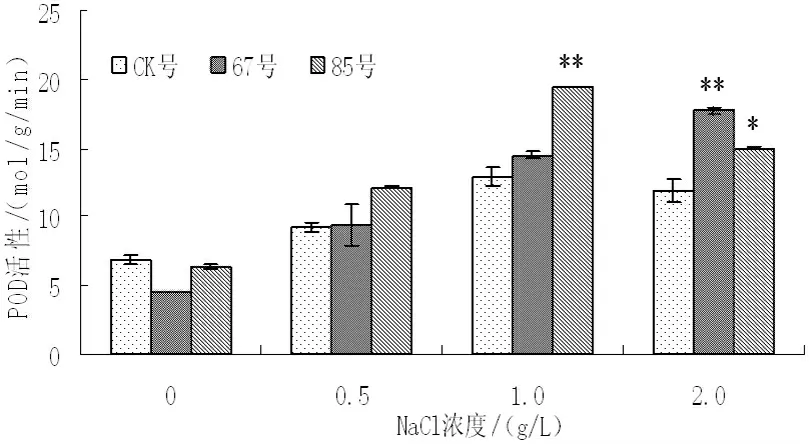
图8 不同NaCl 浓度下POd活性的变化

图9 不同NaCl 浓度下CAT 活性的变化
3 讨论
甜菜碱作为一种非毒性渗透调节物质,在植物受到环境胁迫时在细胞内积累并降低渗透势,还能作为一种保护物质具有极为重要的“非渗透调节”功能,维持生物大分子的结构和完整性,维持其正常的生理功能,解除高浓度盐对酶活性的毒害和保护呼吸酶及能量代谢的过程[7]。大量研究结果表明,本试验采用NaCl 对转基因二色胡子BADH 基因进行诱导。Ishitani 等在300mmol·L-1NaCl 胁迫条件下的大麦叶片中,BADH 的mRNA 含量比对照增加了8 倍,在无盐条件下,BADHmRNA 的量就恢复到接近正常的状态。McCue 等发现当NaCl 的浓度从零逐渐增加到500mmol·L-1时,菠菜叶片中的BADH 的mRNA 含量增加3~4倍。本试验结果也证实了这点,盐胁迫下转基因二色胡枝子BADH 表达明显增加,NaCl 浓度为2%时,转基因株系67号BADH 活性和甜菜碱含量是对照植株的2.5 倍和1.61 倍,转基因株系85号BADH 活性和甜菜碱含量是对照植株的3 倍和1.66倍,这与其他盐胁迫条件下诱导BADH 基因表达的相关报道一致[8]。上述研究结果表明,甜菜碱含量的积累与盐胁迫诱导密切相关,甜菜碱在合成的过程中相关基因的表达可能受盐浓度调控;此外不同株系间BADH 活性和甜菜碱含量也存在一定差异,可能与外源BADH 基因插入的位点及甲基化程度有关。
植物受到盐胁迫情况下,将发生离子区域化作用,即植物细胞将吸收的大量盐离子贮存于液泡从而降低细胞质中的盐浓度,减少盐离子对细胞的毒害,但随着液泡和细胞质之间的渗透势差增大,最终会导致细胞质失水,造成机械损伤;同时植物在抗逆胁迫下细胞内自由基如羟自由基、超氧自由基、单线态氧等会引发膜脂过氧化,造成细胞膜系统的损伤,膜透性增加,使细胞内的大量的无机离子和氨基酸、可溶性糖等小分子外渗[9]。本试验证明,转BADH-反义4CL 基因二色胡枝子在盐胁迫下膜结构较稳定,膜透性变化较小,MDA 积累较少,膜脂过氧化程度较低,受到的伤害较小,因而抗盐性较强。
脯氨酸是一种很好的相容性溶质,作为渗透调节剂和渗透保护剂的效应十分广泛和显著,不仅起着降低细胞内水势的作用,而且能有效地保护和稳定各种酶系以及复合体四级蛋白的四级结构,并能维持细胞膜系统在逆境中的稳定性,充当活性氧的清除剂,降低膜脂过氧化等。在正常环境中脯氨酸含量较低,只有在盐胁迫等逆境条件下脯氨酸的合成反应才被激活。本试验研究结果表明,转基因二色胡枝子叶片中脯氨酸含量显著高于对照,这与耐盐程度的高低与脯氨酸含量呈正相关的报道相一致。但有些研究却表明,脯氨酸的积累与耐盐程度成负相关[10]。也有些人则认为脯氨酸的积累是伤害的结果,不能作为抗性筛选的指标,更适宜作为一个胁迫伤害指标[11]。
植物在逆境胁迫条件下,可溶性蛋白分解速度加快,合成减少,因而蛋白质含量下降,氨基酸含量增多。刘世琦曾报道植物可溶性蛋白中50%以上是酶蛋白,其含量的高低可间接反映代谢活动的强弱[12]。本试验研究结果表明,在盐胁迫条件下转基因植株可溶性蛋白的含量高于非转基因植株,间接说明转基因植株代谢活动比非转基因植株强。
CAT、POd和SOd均为植物内源的自由基清除剂,属于保护酶系统。植物在正常条件下,自由基水平较低不会引起伤害,此时自由基的产生与清除处于一种动态平衡之中,但在高盐浓度胁迫条件下,这种动态平衡受到破坏,自由基产生积累,膜脂过氧化。因此,盐胁迫下只有保护酶活性增强或维持较高水平,才能清除活性氧自由基使其保持较低水平,防止生物膜的破坏,从而保持一定的耐盐性。本研究结果表明,盐胁迫条件下,转基因株系CAT、POd和SOd活性均高于非转基因植株,但盐浓度增加到2.0%时,保护酶的活性有所下降,这可能与高盐浓度下植物的保护酶系统受到破坏有关。
猜你喜欢
养猪(2022年1期)2022-11-21
中国畜禽种业(2022年8期)2022-09-17
草业学报(2022年5期)2022-07-08
中国现代中药(2022年1期)2022-03-03
江西农业学报(2021年8期)2021-09-08
上海师范大学学报·自然科学版(2020年5期)2020-12-18
植物研究(2020年6期)2020-03-05
农家科技下旬刊(2017年6期)2017-07-05
发明与创新·中学生(2016年5期)2016-05-14
天津农业科学(2016年4期)2016-04-20
