
贮藏年限对裸燕麦种带真菌和真菌毒素的影响
2021-06-28 11:01聂秀美慕平赵桂琴何海鹏吴文斌蔺豆豆苏伟娟张丽睿
草业学报 2021年6期
聂秀美,慕平,赵桂琴,何海鹏,吴文斌,蔺豆豆,苏伟娟,张丽睿
(1.甘肃农业大学草业学院,甘肃 兰州730070;2.甘肃农业大学农学院,甘肃 兰州730070)
裸燕麦(Avena nuda)是禾本科燕麦属一年生粮饲兼用作物,广泛种植于我国华北和西北等半干旱地区,每年种植面积约60万hm2[1-2]。研究表明,裸燕麦籽粒含有大量易消化和高热量的营养物质,如粗蛋白(12%~18%)、粗脂肪(4%~6%)、矿物质和可溶性膳食纤维(11%~18%)等,能够促进人体和畜禽肠道系统中有益菌的繁殖,调节血脂和血糖,改善血液循环等[3-4]。此外,燕麦籽粒因其粗蛋白、粗脂肪和可溶性糖含量高,难以消化的粗纤维含量低,是牛、马等家畜补饲肥膘的优质饲料[5]。
随着人们对裸燕麦种子保健功能认识的逐步提高,以及国家“粮改饲”“引草入田”和“草牧业”等重大政策的推行实施,燕麦种植面积和种子需求量不断增加。种子在生产和贮藏过程中不可避免地会携带大量的真菌,这些真菌可通过自身的分解作用对种子中的有机物质进行消化、吸收和利用,导致种子劣变、缩短种子贮藏年限、增加病原菌传播风险,甚至影响种子安全贮藏和幼苗健康[6-7]。此外,种子在贮藏过程中,大多数霉菌在适宜的生境下会代谢产生真菌毒素,长期摄入低含量毒素会引起代谢紊乱、免疫力下降以及皮肤、肝脏和胃肠道等的病变;短期内摄入毒素含量过高,可引起生殖毒性、细胞毒性、癌变和畸变等,对人畜健康具有极大威胁[8-9]。
据联合国统计,全世界每年被真菌毒素污染的谷物占25%左右,即使某些种子外表未发现霉变,却已被霉菌毒素侵染[10]。因此,种子带菌是危害人畜健康和安全的一大因素,对种子携带的真菌和真菌毒素进行早期检测至关重要。但截至目前,国内对裸燕麦种子的带菌检测仅见荆卓琼等[11]通过形态学观察,从2个裸燕麦品种中分离鉴定出11个属的真菌;马从[12]对甘肃省的2个裸燕麦品种通过形态学观察检出6属14种真菌;聂秀美等[13]通过形态学和分子生物学鉴定,从不同产地的4份裸燕麦材料中分离出真菌13属30种。而有关贮藏年限对裸燕麦种子在贮藏过程中受真菌污染、带菌及产毒情况等方面的研究还尚未见报道。因此,本研究拟以3个主栽品种为研究对象,对其在贮藏期间所携带的真菌和其产生的毒性较强的真菌毒素进行检测,以明确贮藏年限对裸燕麦种子带菌情况和真菌毒素含量的影响,为裸燕麦种子的安全贮藏和生产利用提供科学参考。
1 材料与方法
1.1 试验材料
供试裸燕麦品种为白燕2号、坝莜9号和坝莜14号,于2017、2016、2015和2013年在甘肃省定西市通渭县华家岭镇收获,收获的种子贮藏于甘肃农业大学牧草实训基地种子室,种子贮藏期间的月均温和月均相对湿度(用HC-520温湿度测量仪记录)(图1)。试验于2018年6月在甘肃农业大学草业学院实验室进行,供试材料分别贮藏了1、2、3和5年。
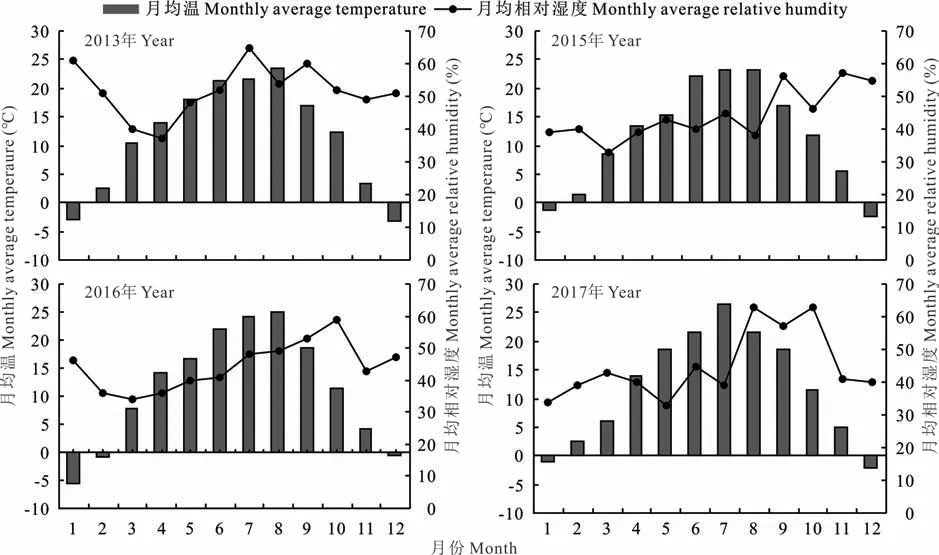
图1 种子贮藏期间的月均温和月均相对湿度Fig.1 Monthly average temperature and relative humidity during seed storage
1.2 种子带菌检测
1.2.1 种子外部带菌检测 参照耿丽华等[7]和高苇等[14]的方法稍做改动后进行种子带菌检测。每个品种随机取200粒完整种子分别放入50 mL灭菌三角瓶,加无菌水20 mL后于180 r·min-1的摇床上充分振荡1 h。在无菌条件下吸取1 mL菌悬液均匀涂布到直径90 mm的PDA(potato dextrose agar,马铃薯葡萄糖琼脂)平板上(内加硫酸链霉素和卡那霉素各100 mg·L-1),每处理3个重复,涂布好的平板置于25℃恒温培养箱中黑暗培养。从第3天起进行观察,及时挑取菌落至新鲜PDA培养基上进行分离,对分离出的真菌进行纯化和保存。待不再有新的菌落出现时,按稀释倍数和检测种子数计算孢子负荷量和检出真菌的分离率。

1.2.2 种子内部带菌检测 每个品种随机选取200粒完整种子,于超净工作台经75%酒精浸泡2 min,1%次氯酸钠消毒5 min,无菌水漂洗4次,灭菌滤纸吸干种子表面的水分后,均匀摆放在90 mm的PDA培养基上(内加硫酸链霉素和卡那霉素各100 mg·L-1),每皿10粒,5皿为一重复,4次重复。置于25℃恒温培养箱中黑暗培养。从第3天起进行观察,及时挑取菌落至新鲜PDA培养基上进行分离、纯化和保存。待不再有新的菌落出现时,统计带菌种子数和带某种真菌的种子数,并计算种子带菌率和某菌分离率。

1.3 形态学鉴定
将分离纯化出的真菌在25℃黑暗培养箱再次纯化培养7 d后,参考《植物病原真菌学》[15]和《真菌分类学》[16],用Revolve RVL-100-G显微镜观察菌落和分生孢子的颜色、形状和大小等形态学特征,进行初步鉴定。
1.4 分子生物学鉴定
采用Omega真菌基因组试剂盒提取菌株DNA。
利用真菌通用引物对ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)进行扩增。其中PCR反应体系为50μL:2×Taq PCR Master Mix 25μL,模板5μL,正反向引物各1μL,补充ddH2O至50μL。扩增程序为:94℃预变性5 min;94℃变性30 s,52℃退火30 s,72℃延伸1 min,34个循环;72℃下延伸7 min。反应完成后,用1%琼脂糖凝胶进行电泳检测和紫外凝胶成像仪观察,然后送样进行测序及比对分析后,用Mega 7.0软件中的邻接法(neighbor joining,NJ)构建系统发育树。
1.5 真菌毒素含量的检测
1.5.1 标准溶液的配制 细交链孢酮酸(tenuazonic acid,TeA)、黄曲霉毒素B1(aflatoxin B1,AFB1)和桔青霉素(citrinin,CIT)3种真菌毒素(规格均为1 mg,纯度>99%)均用纯甲醇溶解,配制成浓度均为100μg·mL-1的母液。吸取TeA、AFB1和CIT母液各500μL,配成浓度为500 ng·mL-1的TeA标准溶液、AFB1和CIT混合标准溶液,并用甲醇依次稀释成浓度为100、50、20、8和0.4 ng·mL-1的标准溶液,存贮于-20℃冰箱中备用。
1.5.2 样品处理 参照王芹等[17]和任贝贝[18]的方法,做适当改进后进行样品中毒素含量的检测。每份种子样品称取20.00 g(精确到0.01 g),经小型谷物粉碎机粉碎后各取5.00 g(精确到0.01g)于50 mL离心管,重复3次。各样品加入30 mL乙腈-水溶液(84∶16,V∶V)经QL-866涡旋振荡仪涡旋混匀后于室温下浸泡1 h,30℃恒温超声30 min,10000 r·min-1离心10 min,取上清液25 mL于-20℃冰箱静置3 h后,重新吸取上清液20 mL在50℃下经Christ RVC 2-33 IR真空离心浓缩仪浓缩至近干,残渣加入乙腈-水(40∶60,V∶V)混合溶液1 mL,经涡旋振荡仪混匀溶解后超声5 min,经0.22μm有机滤膜过滤后用Agilent 1100高效液相色谱质谱仪(high performance liquid chromatography mass spectrometry,HPLC-MS)检测。
1.6 数据分析
采用Microsoft Excel 2010进行数据整理,用DNAStar软件进行序列拼接、校对,在NCBI核酸数据库中进行BLAST序列同源性比对分析,用Mega 7.0软件构建系统发育树,用SPSS 19.0进行方差分析和相关性分析。
2 结果与分析
2.1 贮藏年限对裸燕麦种子带菌量的影响
由表1可知,随贮藏年限的变化,各供试种子外部的孢子负荷量差异较大,为0.60~8.40 cfu。其中白燕2号在贮藏5年时的孢子负荷量(1.57 cfu)最高,贮藏3年时仅有0.60 cfu,显著低于其他年限(P<0.05)。坝莜9号贮藏3年时的负荷量最高(4.17 cfu,P<0.05),贮藏2年时的负荷量最低(1.73 cfu),较前者降低了58.51%。坝莜14号贮藏1年时的孢子负荷量为8.40 cfu,显著高于其他年限(P<0.05),且高于其他参试材料。
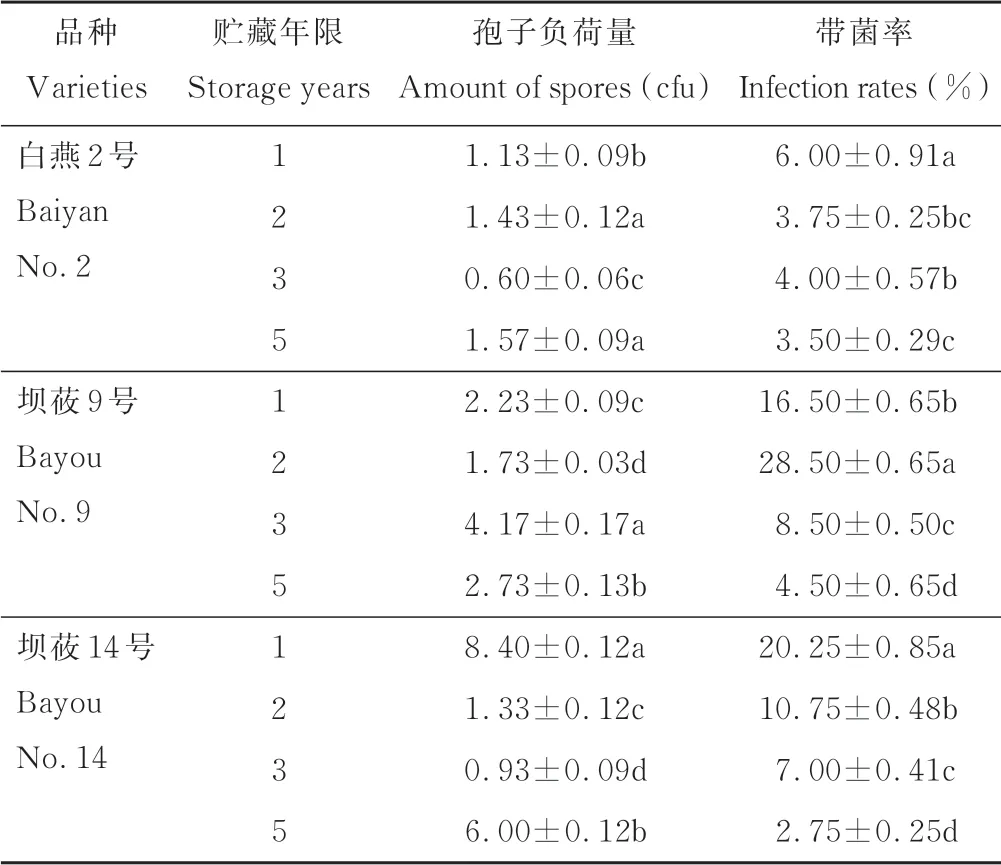
表1 不同贮藏年限裸燕麦种子的带菌量Table 1 The amount of fungi in naked oat seeds with different stor age year s
各参试材料在不同贮藏年限的内部带菌率存在明显差异,随贮藏年限的增加总体表现为下降趋势。带菌率变幅为2.75%~28.50%,其中白燕2号在贮藏1年时带菌率最高,为6.00%(P<0.05)。贮藏2年时坝莜9号带菌率为28.50%,较贮藏1、3和5年分别增加了42.11%、70.18%和84.21%(P<0.05),且高于其他材料。坝莜14号在贮藏1年时带菌率最高,为20.25%,随后显著下降,贮藏5年时带菌率最低(2.75%),较贮藏1年时降低了86.42%。
2.2 裸燕麦种带真菌的鉴定
部分分离真菌的菌落、孢子形态特征及分子生物学鉴定结果如下:
AS2在PDA培养基上生长较快,直径约62 mm,菌落隆起,呈粗绒状,正面中央褐色至灰褐色,边缘呈白色,背面黑褐色,边缘褐色,具同心轮纹。分生孢子单生或串生,倒棍棒状、倒梨形或椭圆形,淡黄褐色至褐色,具横隔1~6个,纵隔0~3个,部分含有短喙,大小为(9.1~24.2)μm×(3.4~8.5)μm,根据形态特征可初步鉴定为细交链孢(Alternaria alternate,图2)。进一步通过序列比对和构建系统进化树发现,AS2与A.alternate(MK656442)聚在同一分支,说明二者亲缘关系很近,由此鉴定为A.alternate。
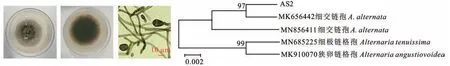
图2 细交链孢的形态特征及系统进化树Fig.2 Morphological characteristics and phylogenetic tree of A.alternate
AS6在PDA培养基上生长迅速,3~4 d即可长满整个平板,菌落呈绒毛状或棉絮状,正面初期白色,后中央为白色,边缘呈淡黄色,背面呈黄褐色,具同心环。分生孢子单生,呈球形或近球形,褐色至深褐色,无隔膜,大小(11.6~14.6)μm×(10.6~13.8)μm,根据形态特征可初步鉴定为节菱孢菌(Arthrinium rasikravindrae,图3)。进一步通过序列比对和构建系统进化树发现,AS6与A.rasikravindrae(MF326617)聚在同一分支,说明二者亲缘关系很近,由此鉴定为A.rasikravindrae。

图3 节菱孢菌的形态特征及系统进化树Fig.3 Morphological characteristics and phylogenetic tree of A.rasikravindrae
AS9在PDA培养基上生长较快,直径约84 mm,菌落平坦干燥,呈粗粉状,正面黄绿色,背面呈焦黄色。分生孢子单生,呈球形,黄绿色,无隔膜,大小为2.3~5.0μm,根据形态特征可初步鉴定为黄曲霉(Aspergillus flavus,图4)。进一步通过序列比对和构建系统进化树发现,AS9与A.flavus(MN095161)聚在同一分支,说明二者亲缘关系很近,由此鉴定为A.flavus。
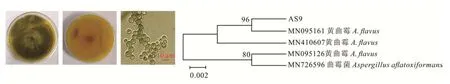
图4 黄曲霉的形态特征及系统进化树Fig.4 Morphological char acter istics and phylogenetic tr ee of A.flavus
AS17在PDA培养基上生长缓慢,直径约15 mm,菌落稍隆起,中央有裂纹,呈粉状,正面橄榄绿色,背面橄榄褐色。分生孢子单生,近球形或卵形,大小为(3.2~6.7)μm×(1.8~2.9)μm,根据形态特征可初步鉴定为枝细枝孢(Cladosporium ramotenellum,图5)。进一步通过序列比对和构建系统进化树发现,AS17与C.ramotenellum(MG548565)和C.ramotenellum(MH 383201)聚在同一分支,说明三者亲缘关系很近,由此鉴定为C.ramotenellum。
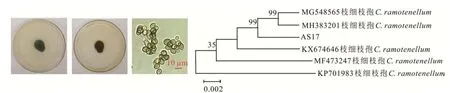
图5 枝细枝孢的形态特征及系统进化树Fig.5 Morphological char acter istics and phylogenetic tr ee of C.ramotenellum
AS29在PDA培养基上生长缓慢,直径约21 mm,菌落呈粉状兼绒状,正面初期蓝绿色,具脐状突起,背面中央淡红褐色,边缘奶黄色。分生孢子单生,呈球形,蓝绿色,无隔膜,大小1.9~3.4μm,根据形态特征可初步鉴定为鲜绿青霉(Penicillium verrucosum,图6)。进一步通过序列比对和构建系统进化树发现,AS29与P.verrucosum(MH 860078)聚在同一分支,说明二者亲缘关系很近,由此鉴定为P.verrucosum(图6)。
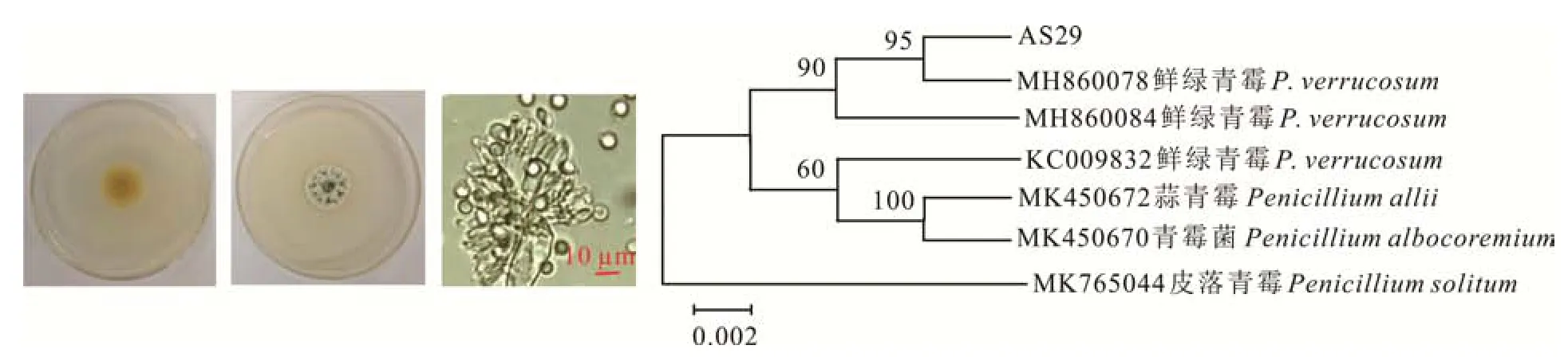
图6 鲜绿青霉的形态特征及系统进化树Fig.6 Morphological characteristics and phylogenetic tree of P.verrucosum
AS32在PDA培养基上生长迅速,2~3 d即可长满整个平板,菌落呈丝状,正面初期乳白色,后期灰褐色。分生孢子单生,呈近球形或卵圆形,淡褐色,无隔膜,大小为(4.6~12.5)μm×(3.9~11.3)μm,根据形态特征可初步鉴定为米根霉(Rhizopus oryzae,图7)。进一步通过序列比对和构建系统进化树发现,AS32与R.oryzae(NR103595)聚在同一分支,说明二者亲缘关系很近,由此鉴定为R.oryzae。

图7 米根霉的形态特征及系统进化树Fig.7 Morphological characteristics and phylogenetic tree of R.oryzae
2.3 贮藏年限对裸燕麦种子带菌种类和分离率的影响
2.3.1 贮藏年限对裸燕麦种子外部带菌种类和分离率的影响 从各供试裸燕麦种子表面共检出真菌14属29种(表2),其中细交链孢在11份样品中检出,检出率为91.67%;其次是黄曲霉,检出率为66.67%。带菌种类最多的是贮藏1和5年的坝莜14号,均携带11种真菌;贮藏1年的白燕2号和贮藏3年的坝莜9号中真菌种类检出数最少,均仅有4种。

表2 裸燕麦种子表面的带菌种类和分离率Table 2 Fungal species and isolation rates of naked oat seeds surface(%)
白燕2号在贮藏2、3和5年时均能检出的真菌有细交链孢、塔宾曲霉(Aspergillustubingensis)和米根霉,而雅致放射毛霉(Actinomucor elegans,19.92%)、枝状枝孢菌(Cladosporium cladosporioides,57.93%)和疣孢漆斑菌(Myrothecium verrucaria,11.64%)仅存在于贮藏1年的种子中;烟曲霉(Aspergillus fumigatu,4.86%)和黑附球菌(Epicoccum nigrum,4.31%)仅在贮藏2年时发现;芸薹链格孢(Alternaria brassicae,13.33%)和黑曲霉(Aspergillusniger,12.04%)仅存在于贮藏3年的种子中;节菱孢菌(1.85%)仅在贮藏5年时检出。葱链格孢(Alternaria porri)、黄曲霉和拟孢镰孢菌(Fusarium sporotrichioid es)仅在贮藏2和3年时分离出,产黄青霉(Penicillium chrysogenum)存在于贮藏3和5年的种子中。
坝莜9号在所有贮藏年限中均能检出的真菌仅有细交链孢,而芸薹链格孢(2.98%)、藜生链格孢(Alternaria chenopodiicola,19.16%)、葱链格孢(12.01%)和楸子茎点霉(Didymella pomorum,1.39%)仅在贮藏1年时检出;寄生曲霉(Aspergillus parasiticus,3.44%)、枝细枝孢(3.70%)和产黄青霉(21.18%)仅在贮藏2年的种子中分离出;阿姆斯特丹曲霉(Aspergillus amstelodami,0.90%)和烟曲霉(4.51%)仅存在于贮藏3年的种子中;黑曲霉(2.08%)、拟孢镰孢菌(6.61%)和米根霉(1.04%)仅在贮藏5年时检出;塔宾曲霉仅在贮藏1和2年时检测出,谢瓦氏曲霉(Aspergilluschevalieri)在贮藏3和5年时检出。
细交链孢在坝莜14号所有贮藏年限的样品中均能检出,而拟孢镰孢菌(1.20%)和鲜绿青霉(20.58%)仅在贮藏1年时分离出;塔宾曲霉(5.25%)、黑附球菌(2.22%)和德氏霉菌(Pyrenophora chaetomioides,32.76%)仅存在于贮藏2年的种子中;黑曲霉(7.41%)和裂褶菌(Schizophyllum commune,3.70%)在贮藏3年时分离出;红曲霉(Aspergillusruber,4.32%)、肉桂紫青霉(Penicillium cinnamopurpureum,1.08%)和米根霉(2.82%)仅在贮藏5年的样品中检出。葱链格孢和木贼镰刀菌(Fusarium equiseti)在贮藏1和2年时都有检出,藜生链格孢和产黄青霉仅存在于贮藏2和3年的种子中,谢瓦氏曲霉和烟曲霉仅在贮藏3和5年时检出。
2.3.2 贮藏年限对裸燕麦种子内部带菌种类和分离率的影响 从不同贮藏年限的裸燕麦种子内部共检出12属23种真菌(表3),其中最多的是细交链孢(100.00%),次高的是芸薹链格孢和黄曲霉,均为66.67%。贮藏1年的坝莜14号所携带的真菌种类最多,为10种;贮藏5年的坝莜14号仅有4种。
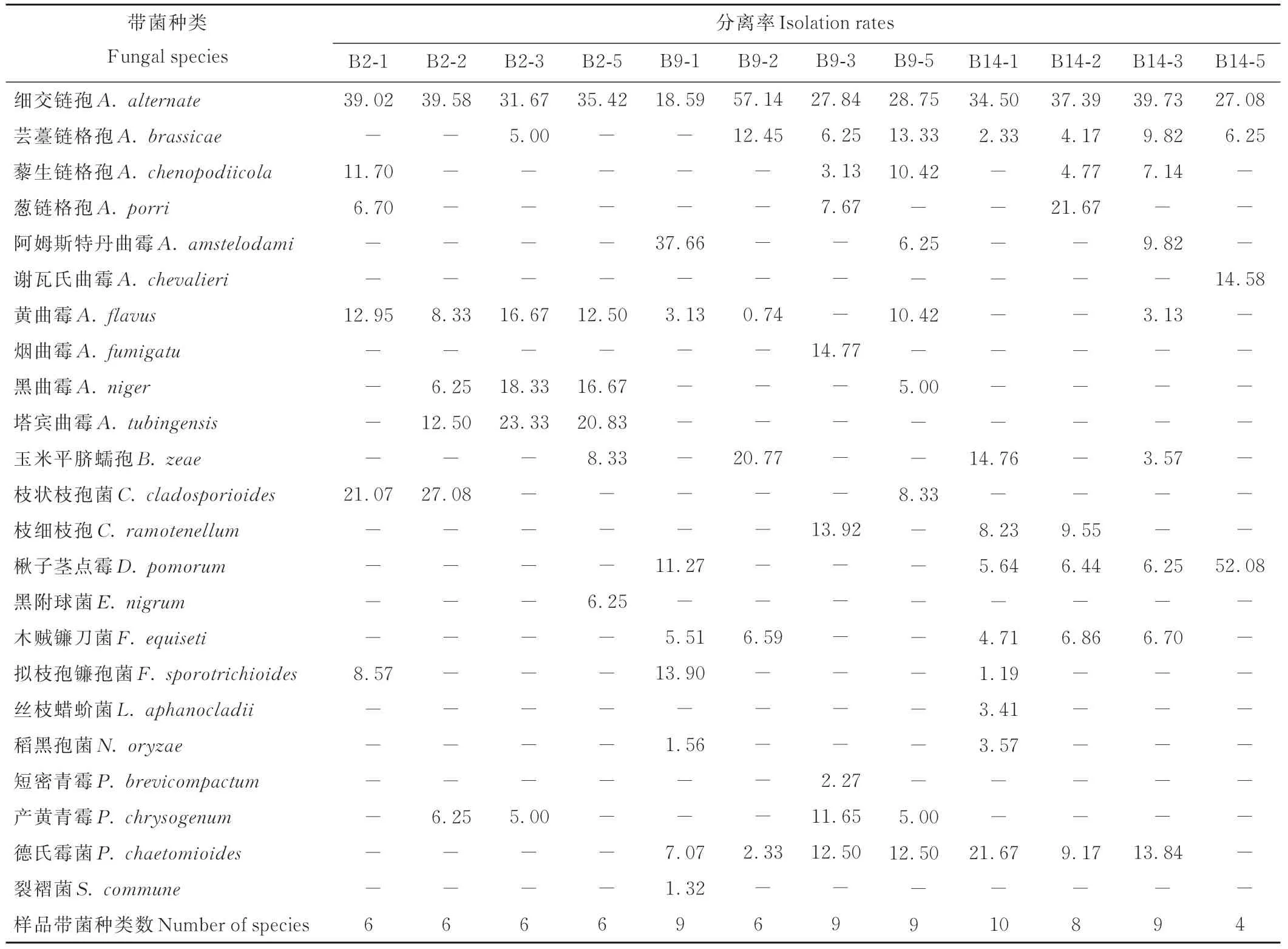
表3 裸燕麦种子内部的带菌种类和分离率Table 3 Internal fungal species and isolation rates of naked oat seeds(%)
在不同贮藏年限的白燕2号种子内部都分离出了细交链孢和黄曲霉。黑曲霉和塔宾曲霉在贮藏2~5年时均可检出,且其分离率在第3年时最大;而藜生链格孢(11.70%)、葱链格孢(6.70%)和拟枝孢镰孢菌(8.57%)仅在贮藏1年时检测出;芸薹链格孢(5.00%)仅从贮藏3年的种子中分离出,玉米平脐蠕孢(Bipolaris zeae,8.33%)和黑附球菌(6.25%)仅在贮藏5年时发现。在贮藏1和2年的种子中均可检出的仅有枝状枝孢菌。产黄青霉仅从贮藏2和3年时的种子中检出。
坝莜9号在贮藏1年时的优势菌为阿姆斯特丹曲霉(37.66%);其他年限优势菌均为细交链孢,分离率为18.59%~57.14%。所有样品中均可检出的真菌有细交链孢和德氏霉菌。贮藏2~5年时可检出的仅有芸薹链格孢。仅从贮藏1年的种子中检测到的是拟枝孢镰孢菌(13.90%)、楸子茎点霉(11.27%)、稻黑孢菌(Nigrospora oryzae,1.56%)和裂褶菌(1.32%)。仅在贮藏2年时检测出的是玉米平脐蠕孢(20.77%)。葱链格孢(7.67%)、烟曲霉(14.77%)、枝细枝孢(13.92%)和短密青霉(Penicillium brevicompactum,2.27%)仅从贮藏3年的种子中分离出,黑曲霉(5.00%)和枝状枝孢菌(8.33%)仅在贮藏5年时发现。木贼镰刀菌仅在贮藏1和2年时检测出,藜生链格孢和产黄青霉仅在贮藏3和5年检出。
坝莜14号在贮藏5年时种子内部分离率最高的是楸子茎点霉,为52.08%;其他年限分离率最高的均为细交链孢。不同贮藏年限共有的真菌有细交链孢、芸薹链格孢和楸子茎点霉。贮藏1~3年的种子内部可检出的仅有木贼镰刀菌和德氏霉菌,而拟枝孢镰孢菌(1.19%)、丝枝蜡蚧菌(Lecanicillium aphanocladii,3.41%)和稻黑孢菌(3.57%)仅在贮藏1年时检测出;葱链格孢(21.67%)仅在贮藏2年时检测出;阿姆斯特丹曲霉(9.82%)和黄曲霉(1.13%)仅在贮藏3年的种子中分离出;谢瓦氏曲霉(14.58%)仅在贮藏5年时发现。在贮藏1和2年时均可分离出枝细枝孢,而贮藏2和3年时均可检出的仅有藜生链格孢。
2.4 裸燕麦种带真菌的系统发育树
通过对裸燕麦种子上分离获得的不同菌株的基因组DNA进行测序和同源性比对分析,发现各被鉴定菌株与其对应的比对菌株相似度均在97.00%以上。在NCBI数据库提交被鉴定菌株AS1~AS33,获得的菌株序列号为MT 503287~MT 503319。进一步构建系统发育树(图8),发现同一个属的菌株基本都聚在一个较大的分支,不同属的菌株单独形成一支。共鉴定出17属33种真菌,其中曲霉菌属8种,青霉菌属5种,链格孢属4种,节凌孢属(Arthrinium)1种、枝孢菌属(Cladosporium)和镰刀菌属(Fusarium)各2种,德氏霉属(Drechslera)、毛霉属(Actinomucor)、蠕孢属(Bipolaris)、亚隔孢壳属(Didymella)、附球菌属(Epicoccum)、裂褶菌属(Schizophyllum)、蜡蚧菌属(Lecanicillium)、漆斑菌属(Myrothecium)、黑孢霉属(Nigrospora)、茎点霉属(Phoma)和根霉属(Rhizopus)各1种。
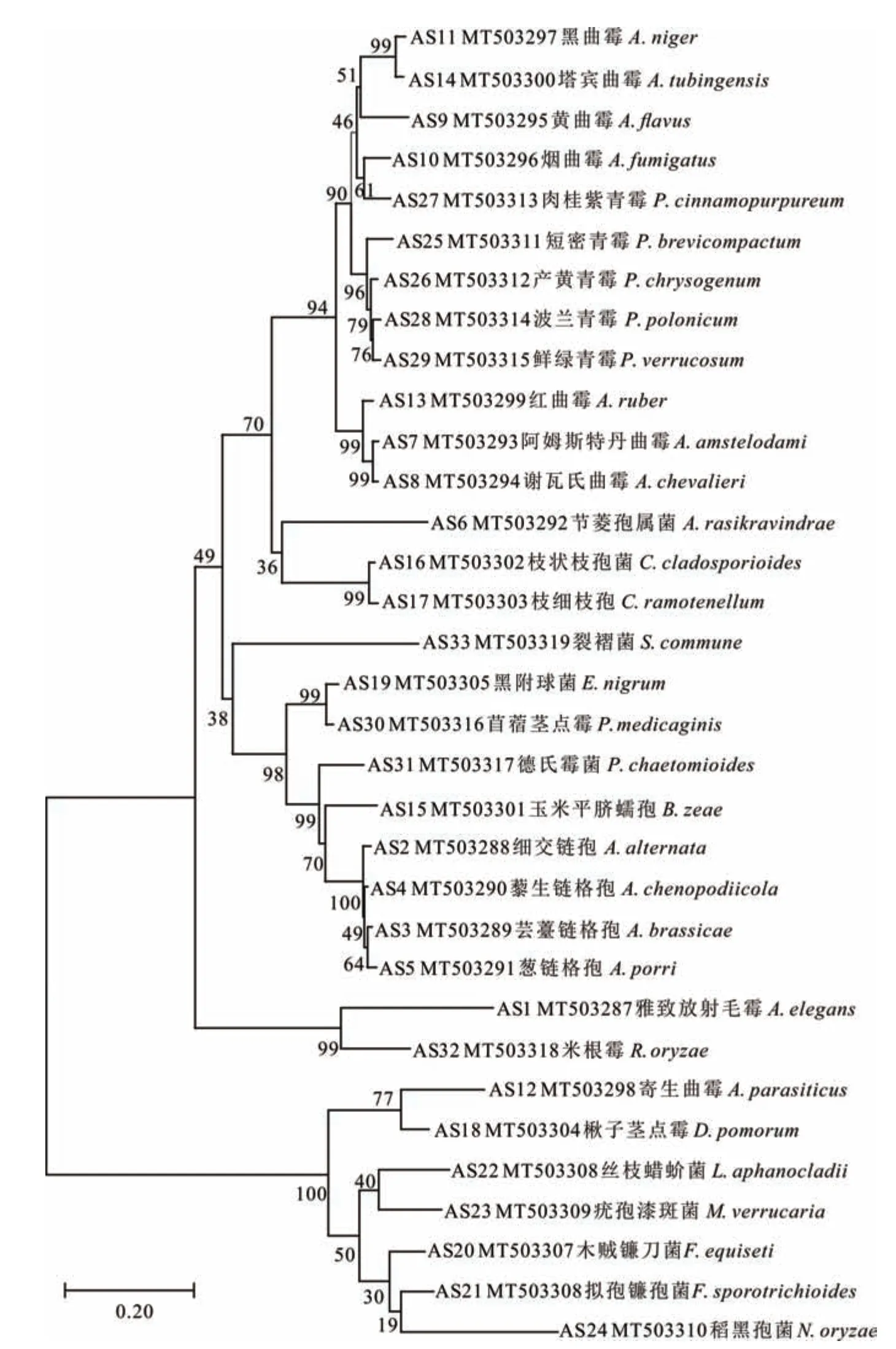
图8 裸燕麦种带真菌的系统发育树Fig.8 Phylogenetic tree of seed borne fungiof naked oat seeds
2.5 真菌毒素的线性方程、线性范围和相关系数
通过对3种真菌毒素的标准样品按500、100、50、20、8和0.4 ng·mL-1的浓度进行检测表明,其线性关系良好,相关系数均在0.9995以上,各标样的线性方程、线性范围和相关系数见表4。
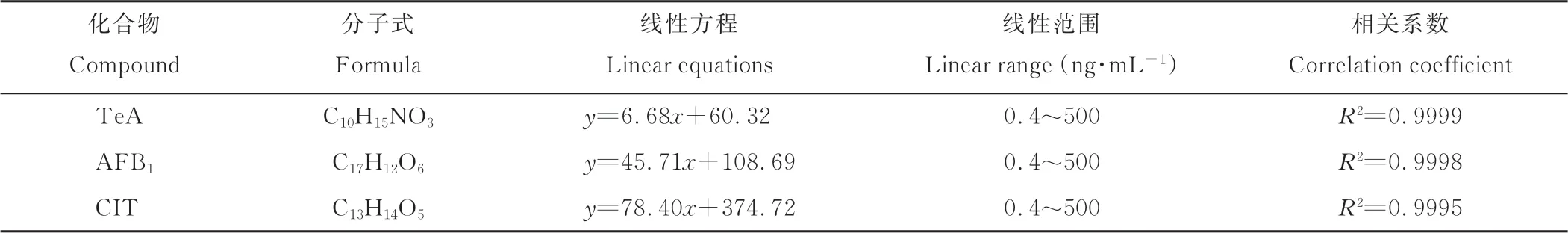
表4 真菌毒素的线性方程、线性范围和相关系数Table 4 Linear equations,linear range and correlation coefficient of mycotoxin
2.6 贮藏年限对裸燕麦种子真菌毒素含量的影响
通过对不同贮藏年限裸燕麦种子的TeA、AFB1和CIT进行检测表明,不同品种的毒素含量差异明显,同一品种不同贮藏年限的毒素含量也有显著差异(表5)。
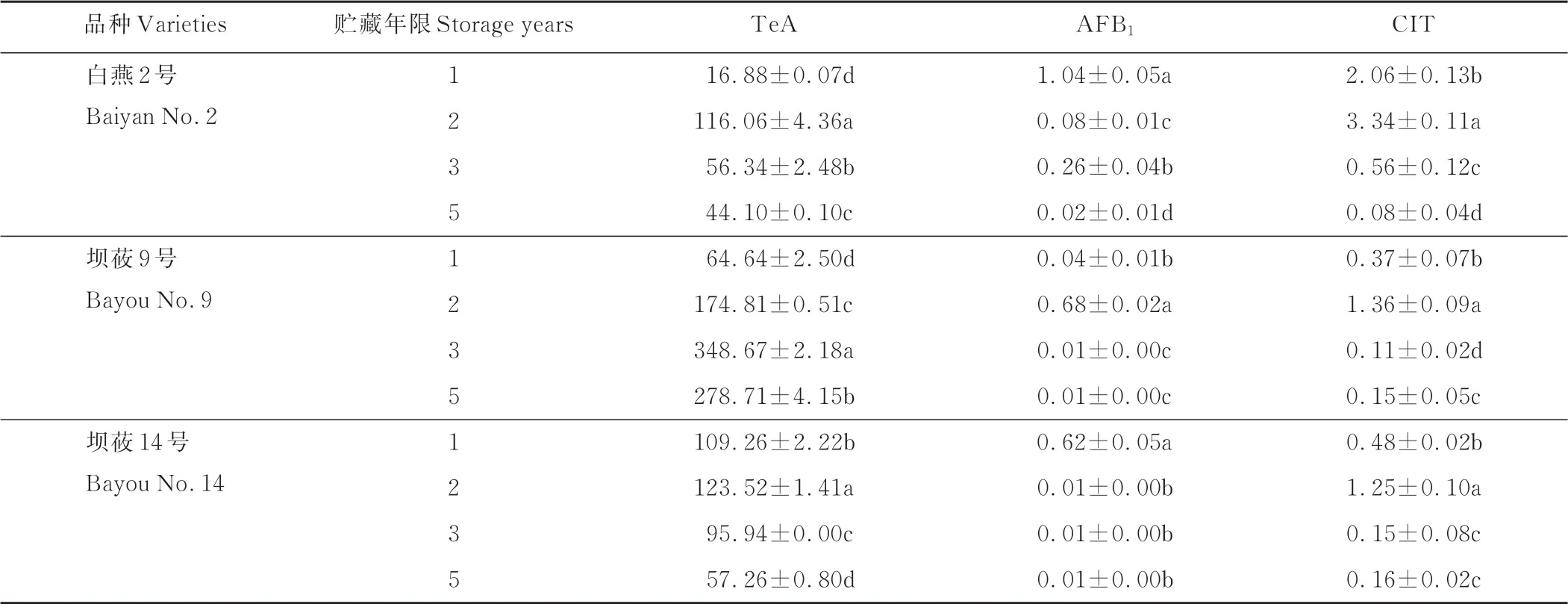
表5 贮藏年限对裸燕麦种子真菌毒素含量的影响Table 5 Effect of stor age year s on mycotoxin content of naked oat seeds(μg·kg-1)
TeA含量随贮藏年限的延长总体呈先升后降的趋势,3个品种中坝莜9号的毒素含量相对最高。白燕2号贮藏2年时的TeA含量最高,为116.06μg·kg-1,显著高于其他3个贮藏年限(P<0.05);贮藏1年时该毒素的检出量最低(16.88μg·kg-1),且明显低于其他材料。坝莜9号贮藏3年时该毒素的含量高达348.67μg·kg-1,是所有处理中最高的;但其在贮藏1年时检出量(64.64μg·kg-1)最低,较贮藏3年时减少了81.46%。坝莜14号在贮藏2年时TeA的检出量最高(123.52μg·kg-1),贮藏5年时其检出量最低,为57.26μg·kg-1,较贮藏2年时下降了53.64%。
AFB1的变化因品种和贮藏年限而异。白燕2号贮藏1年时含量最高,为1.04μg·kg-1,是所有处理中最高的;随后显著下降,至贮藏5年时,该毒素的检出量仅有0.02μg·kg-1,较贮藏1年时下降了98.08%。坝莜9号随贮藏年限的延长其AFB1含量呈先升后降趋势,贮藏2年时检出量最高,为0.68μg·kg-1,随后显著下降,贮藏5年时降至最低(0.01μg·kg-1)。坝莜14号的AFB1含量随贮藏年限的延长而下降,贮藏1年时的含量最高,为0.62 μg·kg-1,随后急剧下降。
CIT含量随贮藏年限的增加总体呈先升后降的趋势,3个品种贮藏2年的CIT含量都显著高于其他年限(P<0.05)。白燕2号的CIT含量在贮藏2年后显著下降,至贮藏5年时,该毒素的检出量(0.08μg·kg-1)为所有处理中最低。坝莜9号在贮藏3年时的含量较贮藏2年时下降了91.91%。坝莜14号在贮藏3年时的含量最低(0.15 μg·kg-1),除与贮藏5年时该毒素的检出量无明显差异外,显著低于其他年限(P<0.05)。
3 讨论
植物种子在生长、成熟、收获和贮藏过程中,难以避免真菌入侵。本试验中,不同贮藏年限参试样品的带菌量存在明显差异,其带菌量随贮藏年限的延长总体呈降低趋势,贮藏时间和品种对燕麦种子的带菌量有显著影响。这与陈焘等[19]对老芒麦(Elymus sibiricu)种子、雷玉明等[20]对河西走廊玉米(Zea mays)种子带菌量的研究结果类似。金柳艳等[21]研究表明,玉米种子的孢子负荷量与带菌率不具有明显关联性,乳熟期涿州种样的孢子负荷量为1448 cfu,带菌率仅有2.5%;完熟期蚌埠种样的孢子负荷量高达1458 cfu,其带菌率却仅有10.0%;新郑种样的负荷量为275 cfu,其带菌率却达到了100.0%。本研究中,燕麦种子的孢子负荷量和带菌率的表现趋势也不一致,如白燕2号在贮藏5年时的孢子负荷量最高,但其带菌率却低于其他年限。贮藏2年的坝莜9号种子表面的负荷量在所有贮藏年限中最低,但其带菌率却最高。这可能跟贮藏时间、品种以及真菌寄生能力密切相关。刚收获贮藏的种子很容易携带其母体及生长环境中的真菌体系,随着贮藏时间的增加,带菌量会因真菌的适应性而发生变化[22];经收获贮藏的种子,通常会处于休眠状态而不能规避真菌的侵染,但随着贮藏时间的延长,种子自身会逐渐解除休眠以抵御真菌的进一步污染[23]。岳晓禹等[24]对不同贮藏年限玉米样品在贮藏过程中的霉菌群落研究表明,其真菌群落的变化与贮藏时间的相关性较强,贮藏1和3年的样品中真菌群落差异明显。前人对芝麻(Sesamum indicum)[25]和棉花(Gossypium hirsutum)种子[26]进行真菌检测表明,种子外部和内部携带的真菌种类差异较大。本研究中,裸燕麦种子在不同贮藏年限下内部和外部的带菌量和带菌种类检出情况差异显著。所有参试品种中,白燕2号种子外部的孢子负荷量相对最少,但其带菌种类却较多;坝莜9号的种子内部带菌率相对最高,但其带菌种类却较少。燕麦种子随贮藏年限的变化其带菌种类和分离率差异也很大,如坝莜9号在贮藏1年时,种子内部分离率最高的真菌为阿姆斯特丹曲霉(37.66%),其他年限最高的均为细交链孢,这跟杨颜霞等[27]对结缕草(Zoysia japonica)种子的研究结果类似。这可能跟种子在贮藏期间的自身防御能力和环境条件有关,不同真菌的侵染强度有所差异,而且同一品种在不同贮藏年份气候因子间的差异也会影响真菌的存活力和繁殖率,进而影响对种子的侵染[11]。
不同品种在不同贮藏年限的种子外部和内部携带的优势真菌种类存在明显差异。如白燕2号贮藏1年时种子外部的优势种为枝状枝孢菌,2年时为拟孢镰孢菌,在3和5年时均为细交链孢。坝莜9号贮藏1年时种子内部的优势菌为阿姆斯特丹曲霉,其他贮藏年限却是细交链孢。尽管优势种不同,但优势属均为链格孢属、曲霉属和青霉属。这和前人对三七(Panax notoginseng)[28]和红豆草(Onobrychis viciaefolia)等[29]的种带真菌研究结果类似。这主要是由于链格孢菌、青霉菌和曲霉菌等腐生菌的寄主范围通常较广,可通过侵染其母本植株、种子及田间杂草进行寄生,其孢子萌发和菌丝生长也相对较快,较有利于其在植物组织或器官侵染和传播[30-31]。本研究发现,燕麦种子外部和内部的真菌情况有所不同,雅致放射毛霉、寄生曲霉、苜蓿茎点霉和米根霉等10种真菌仅在外部检出,而玉米平脐蠕孢、丝枝蜡蚧菌、稻黑孢菌和短密青霉4种仅在内部检出。共同携带的真菌有11种,但其在不同品种或在同一品种不同贮藏年限的分离情况差异也较大。这与赵辉等[25]对芝麻种子和先米西努尔·肉孜[32]对苜蓿(Medicago sativa)种子的研究结果类似,种子经消毒前后的真菌检出情况存在明显差异。这可能是由于某些真菌对次氯酸钠的敏感性较强,外部消毒处理会破坏其生长环境(主要是适宜真菌生长的酸性环境)和降低活性,不利于其孢子的萌发和菌丝的生长发育,从而抑制真菌的生长繁殖。
对燕麦种子携带的优势菌属代谢产生的危害较大的3种毒素含量进行分析,发现各供试样品均不同程度地受到3种毒素的污染,其中污染最重的是TeA,其次是CIT和AFB1。这主要是因为链格孢菌种类繁多、适应性强、繁殖力高、寄主范围广,95%以上的种都能在植物上寄生,特别是禾谷类农作物[33]。王晓红等[34]对薏苡(Coix lacryma-jobi)仁在贮藏过程中的真菌毒素检测表明,在贮藏0~30 d时AFB1含量很低,但在贮藏90 d时其含量明显增加。罗晓杨[35]发现大多数材料的脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)和伏马毒素(fumonisin,FB)含量随贮藏时间的变化而变化,在贮藏前期呈降低趋势,后期会增加或先增加后急剧下降,甚至反复反弹。本研究中,燕麦种子中真菌毒素检出量差异显著,随贮藏年限的变化大多表现为先升后降的趋势。真菌毒素含量在某一贮藏年份会显著高于其他年份,很可能是由于这种毒素的大多数产毒真菌在这一贮藏时间达到了优良生长期,代谢量增大。不同燕麦品种种子的毒素含量差异显著,同一品种在不同贮藏年限的毒素含量也有明显差异。如白燕2号在贮藏1年时的AFB1含量最高,为1.04μg·kg-1,4年后下降至0.02μg·kg-1;白燕2号和坝莜9号在贮藏2年时的CIT含量显著高于其他年限,但白燕2号在贮藏5年时该毒素的检出量最低(0.08μg·kg-1),而坝莜9号在贮藏3年时的含量最低。这与刘冬梅等[36]对远杂9102和豫花15花生(Arachis hypogaea)中毒素含量的研究结果类似,在相同的贮藏条件下,不同品种或同一品种的毒素检出情况会因贮藏时间的长短而存在差异。本试验中,燕麦种子在贮藏期间毒素含量的波动除与真菌自身的代谢状况有关外,还可能与其所处的贮藏环境或其他微生物的分解作用有关。
李俭等[37]研究表明,玉米种子中的AFB1含量随着生霉粒数量的增加呈现上升趋势;而DON和玉米赤霉烯酮(zearalenone,ZEN)含量则上下波动。卢洋等[38]也发现,玉米种子带菌量的多少和毒素检出量不相关,不能通过霉变量直接判断种子中毒素含量的高低。本研究中,裸燕麦种子中真菌毒素含量的多少与带菌量未表现出线性关系,如白燕2号在贮藏1年时的TeA含量显著低于其他3个年限,仅有16.88μg·kg-1;但其带菌率显著高于其他年限,孢子负荷量也较高。贮藏3年的坝莜9号AFB1和CIT含量(分别为0.01μg·kg-1和0.11μg·kg-1)都低于其他处理,但其带菌率和孢子负荷量(分别为8.50%和4.17 cfu)都较高。种子在贮藏过程中受真菌侵染的程度与贮藏环境、真菌自身的代谢状况、真菌间的分解作用以及种子自身的生理机制等都有关系,且其产毒性能不具有均一性[39-40]。本研究中3种真菌毒素含量并未出现同时增加或降低的情况,这在一定程度上说明其产毒真菌并不是同时生长繁殖的,这很可能跟燕麦种子在贮藏期间携带真菌的多样性较高,相互之间存在竞争性有关;而且除真菌外的其他微生物(细菌和放线菌等)及其代谢产物之间都可能发生拮抗作用而抑制某些真菌的产毒代谢活动。
4 结论
1)贮藏年限对裸燕麦种子的带菌量、带菌种类和分离率以及毒素含量都有显著影响。带菌量随贮藏年限的延长逐渐下降,毒素含量呈先增后减的趋势,带菌种类和分离率因贮藏时间的不同差异明显。
2)裸燕麦种子携带的优势真菌属为链格孢菌属、曲霉菌属和青霉菌属,优势种因贮藏时间和品种不同而异。
3)裸燕麦种子的带菌量与带菌种类、真菌毒素含量的多少无明显相关性。
猜你喜欢
甘肃教育(2021年10期)2021-11-02
干旱地区农业研究(2021年4期)2021-08-11
东北农业大学学报(2020年3期)2020-05-14
生物技术通报(2019年2期)2019-03-15
四川农业大学学报(2018年5期)2018-11-09
天然产物研究与开发(2018年7期)2018-08-21
新农村(2018年2期)2018-08-18
种子科技(2017年12期)2017-01-12
天津科技大学学报(2016年1期)2016-02-28
西藏大学学报(自然科学版)(2015年1期)2015-12-29
