
Ac-SDKP 抑制YEATS4 拮抗实验性矽肺的机制
2021-06-17 02:07蔡文臣张诗汇高一冰牛婉丽
中国比较医学杂志 2021年5期
蔡文臣,张诗汇,高一冰,牛婉丽,徐 洪,3,杨 方*
(1.华北理工大学公共卫生学院,河北唐山 063210; 2.华北理工大学临床医学院,河北唐山 063210;3.河北省器官纤维化重点实验室,河北唐山 063210)
矽肺纤维化主要是由于长期吸入游离二氧化硅(SiO2)粉尘引起的,以矽结节形成和弥漫性肺间质纤维化为主要病理变化的一种职业性肺病,发生机制尚未完全阐明[1]。 课题组前期通过高通量转录组测序技术,发现含 YEATS(Yaf9、ENL、AF9、SAS5、TAF14) 结 构 域 蛋 白 4 ( YEATS domain containing protein 4, YEATS4)可能是实验性矽肺模型进展的重要靶基因[2]。 YEATS4 是一种高度保守的,具有YEATS 家族结构域的核蛋白[3]。 作为转录激活因子,YEATS4 参与了多种肿瘤的发生发展[4-5]。 课题组多年来对N-乙酰基-丝氨酰-天门冬酰-赖氨酰-脯氨酰(N-acetyl-seranyl-aspartate-lysylproline, Ac-SDKP)的抗矽肺纤维化作用机制进行了系列研究[6-7]。 因此,本研究拟观察Ac-SDKP 能否通过对YEATS4 的调节,从而抑制巨噬细胞活化,发挥拮抗矽肺纤维化的作用。
1 材料和方法
1.1 实验材料
1.1.1 实验动物
SPF 级 3 周龄 30 只雄性 Wistar 大鼠(180±10)g 购自北京维通利华动物技术有限公司[SCXY(京)2016-0008]。 于华北理工大学医学实验中心洁净级动物房饲养[SYXK(冀)2020-0007], 室温(22±2)℃,相对湿度40%~60%,自由饮水、进食,分笼喂养。 所有操作均符合实验动物伦理学要求(LX2019035),并按照实验动物使用的3R 原则给予人道的关怀。
1.1.2 实验细胞
小鼠单核巨噬细胞RAW264.7(中国科学院上海细胞库)。
1.2 主要试剂与仪器
动式染尘控制系统(天津开发区合普工贸有限公司);兔抗I 型胶原(Collagen type I,COLI)(美国Abcom 公司);鼠抗YEATS4(美国Santa Cruz);鼠抗精氨酸酶-1(Arginase-1)(美国BD 公司);兔抗单核细胞趋化蛋白-1(Monocyte chemoattractant protein-1,MCP-1)(美国 Affinity 公司);α-微管蛋白(Tubulin alpha, Tublin α)(美国 Affinity 公司);二氧化硅(silicon dioxide,SiO2)(美国 Sigma 公司);DMEM 培养基(以色列BI 公司);Ac-SDKP(瑞士Bachem AG公司);PV6000 通用型两步法免疫组化试剂(北京中杉公司);二抗山羊抗兔,抗鼠IgG(美国KPL 公司);ECL 显色试剂盒(美国GE 公司);苏木精染色液(珠海贝索公司)。
1.3 实验方法
1.3.1 动物分组
采用HOPE-MED8050 动式染尘控制系统制备矽肺大鼠模型[2]。 实验动物分组:①对照24 周组(Control 24 week, C24):吸入纯净气至16 周后,腹腔埋入含有生理盐水的微量缓释胶囊;②矽肺24 周组(silicosis 24 week, S24):动式染尘以(50±10)μg/m3的浓度h/d,染尘至16 周后,腹腔内埋入含有生理盐水的微量缓释胶囊;③Ac-SDKP 治疗组(Ac-SDKP treatment 24 week, Ac24):动式染尘至16 周后,在大鼠腹腔内埋入含有Ac-SDKP[800 μg/(kg·d)]的微量缓释胶囊,均于24 周处理动物。 取右肺下叶组织置于4%多聚甲醛中固定,常规石蜡
1.4 统计学方法
采用SPSS 20.0 软件进行统计学分析。 数据以平均数±标准差()表示。 采用独立样本t检验或单因素方差分析评估组间差异,进一步两两比较采用SNK-q检验。P<0.05 为差异有统计学意义。
2 结果
2.1 沉默 YEATS4 可抑制 SiO2 诱导的RAW264.7 活化
如图1、表1 所示,Western blot 结果显示,与siC组比较,siY-3 明显减弱了 YEATS4 的表达(P<0.05)。 如图 2、表 2 所示,与 siC 组比较,沉默YEATS4 后,COL I、MCP-1、Arginase-1 表达明显降低,差异均具有统计学意义(P<0.05)。 给予SiO2刺激 的 同 时 沉 默 YEATS4 后, COL I、 MCP-1、Arginase-1 表达也明显降低(表3),差异均具有统计学意义(P<0.05)。
2.2 Ac-SDKP 对 SiO2 诱导的 RAW264.7 细胞中YEATS4 表达的调节作用
免疫细胞化学染色结果显示,与对照组比较,SiO2刺激组中 YEATS4 高表达;与 SiO2刺激组比较, Ac-SDKP 处理组中,YEATS4 阳性表达明显降低(图 3)。 如图4、表 4 所示,与对照组比较,SiO2刺激组中 COL I、MCP-1、Arginase-1 和 YEATS4 蛋白表达均升高,差异均具有统计学意义(P<0.05);与SiO2刺激组比较, Ac-SDKP 处理组中,COL I、MCP-1、Arginase-1 和 YEATS4 蛋白表达均降低,差异均具有统计学意义(P<0.05)。
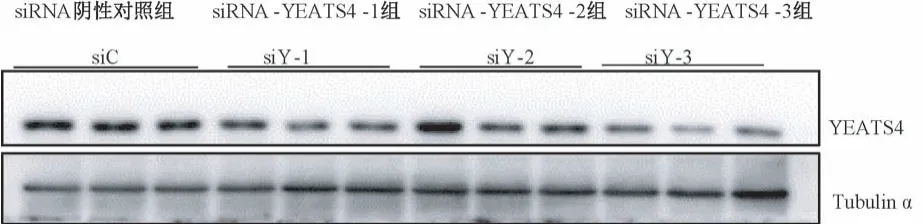
图1 YEATS4 沉默序列筛选Figure 1 YEATS4 silencing sequence screening
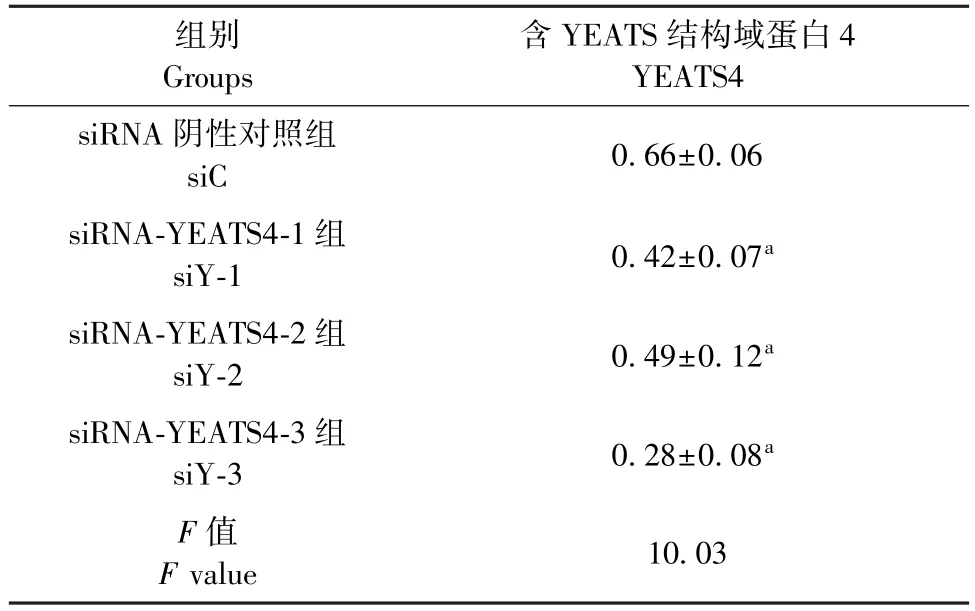
表1 YEATS4 沉默序列筛选Table 1 YEATS4 silencing sequence screening

图 2 COL I、MCP-1、Arginase-1 蛋白在沉默YEATS4 的 RAW264.7 中的表达Figure 2 The levels of COL I, MCP-1 and Arginase-1 in RAW264.7 regulated by silencing YEATS4
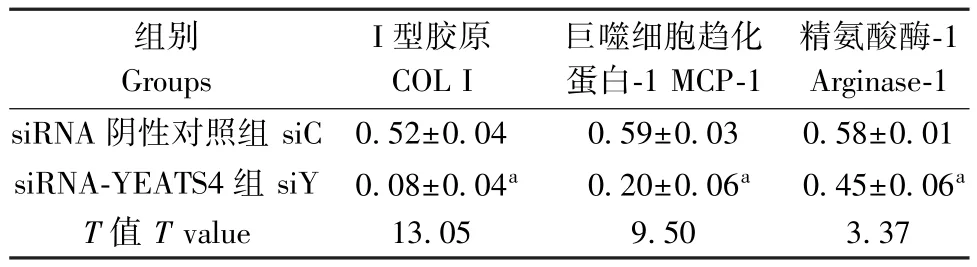
表 2 COL I、MCP-1、Arginase-1 蛋白在沉默YEATS4 的 RAW264.7 中的表达Table 2 The levels of COL I, MCP-1 and Arginase-1 in RAW264.7 regulated by silencing YEATS4
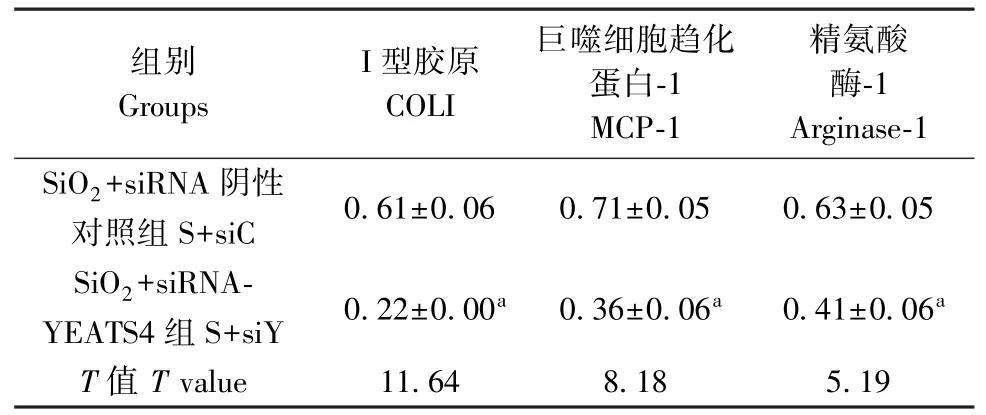
表 3 COL I、MCP-1、Arginase-1 蛋白在 SiO2 诱导的沉默YEATS4 的 RAW264.7 中的表达Table 3 The levels of COL I, MCP-1 and Arginase-1 in silica-treated RAW264.7 regulated by siRNA-YEATS4
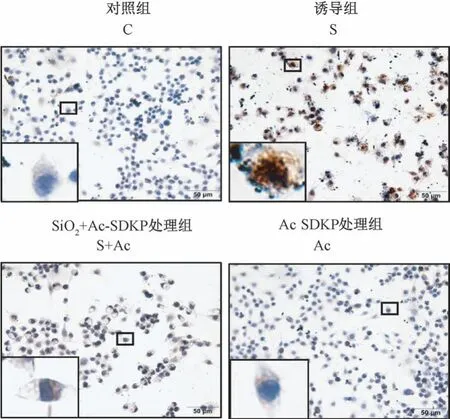
图3 Ac-SDKP 对SiO2 诱导的RAW264.7 细胞中YEATS4 表达的调节作用Figure 3 Ac-SDKP inhibits the high-expression of YEATS4 in RAW264.7 cells induced by silica
2.3 Ac-SDKP 对实验性矽肺大鼠肺组织中YEATS4 的表达的调节作用
IHC 结果显示,与 C24 周组比较,S24 周组中YEATS4 阳性表达主要定位于矽结节中的巨噬细胞;与S24 周组比较,Ac-SDKP 处理组中YEATS4 阳性表达细胞数量减少(图5)。 如图6、表5 所示,与C24 周组比较,S24 周组中 COL I、MCP-1、Arginase-1和YEATS4 表达均升高,差异均具有统计学意义(P<0.05)。 与 S24 周组比较,Ac24 周组中,COL I、MCP-1、Arginase-1 和 YEATS4 蛋白表达明显降低,差异均具有统计学意义(P<0.05)。
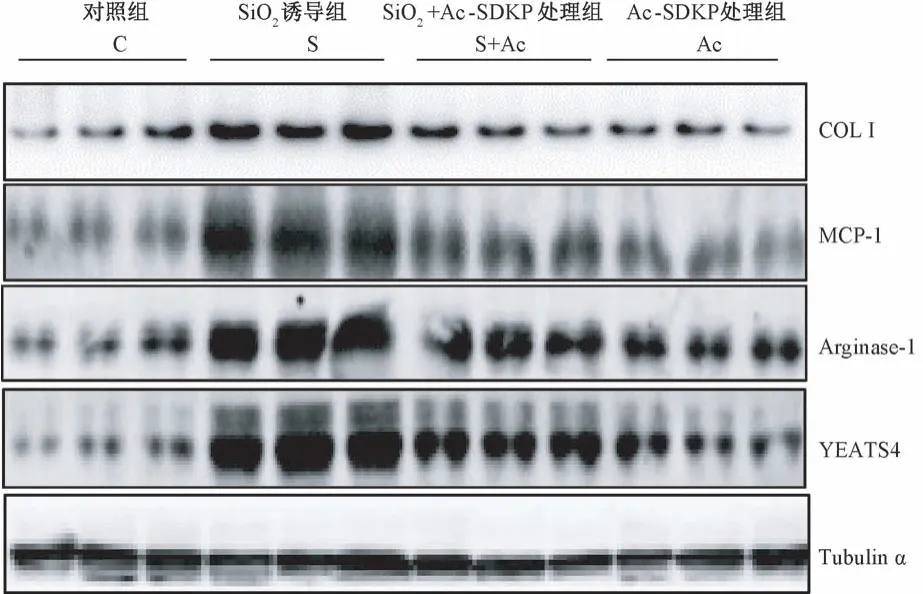
图4 Ac-SDKP 对 SiO2 诱导的 RAW264.7细胞中YEATS4 表达的调节作用Figure 4 Ac-SDKP inhibits the high-expression of YEATS4 in RAW264.7 cells induced by silica
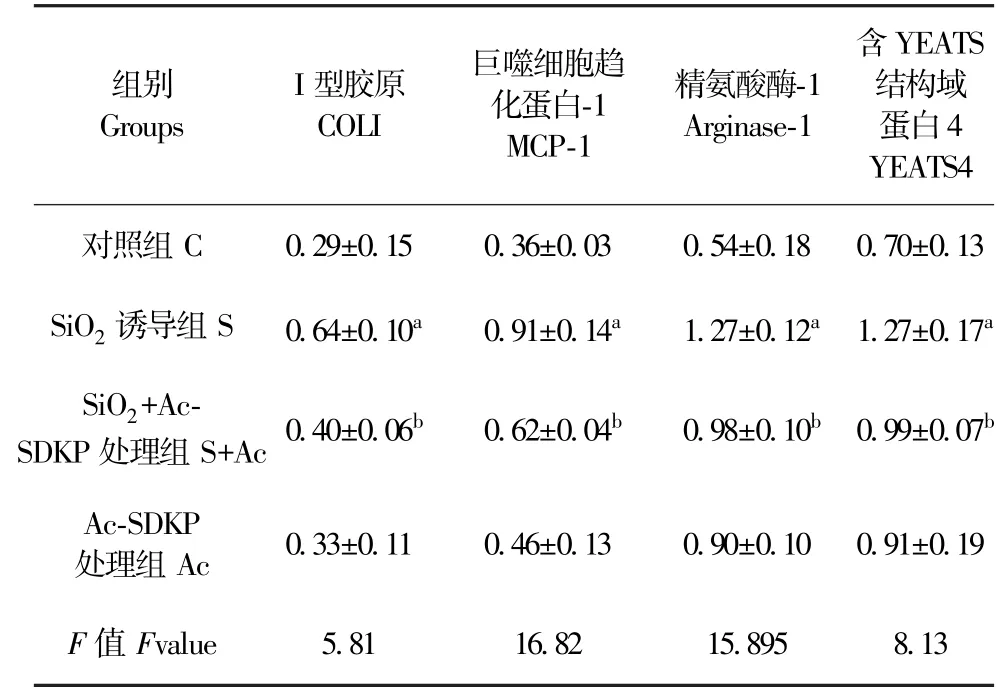
表4 Ac-SDKP 对 SiO2 诱导的 RAW264.7细胞中YEATS4 表达的调节作用Table 4 Ac-SDKP inhibits the high-expression of YEATS4 in RAW264.7 cells induced by silica
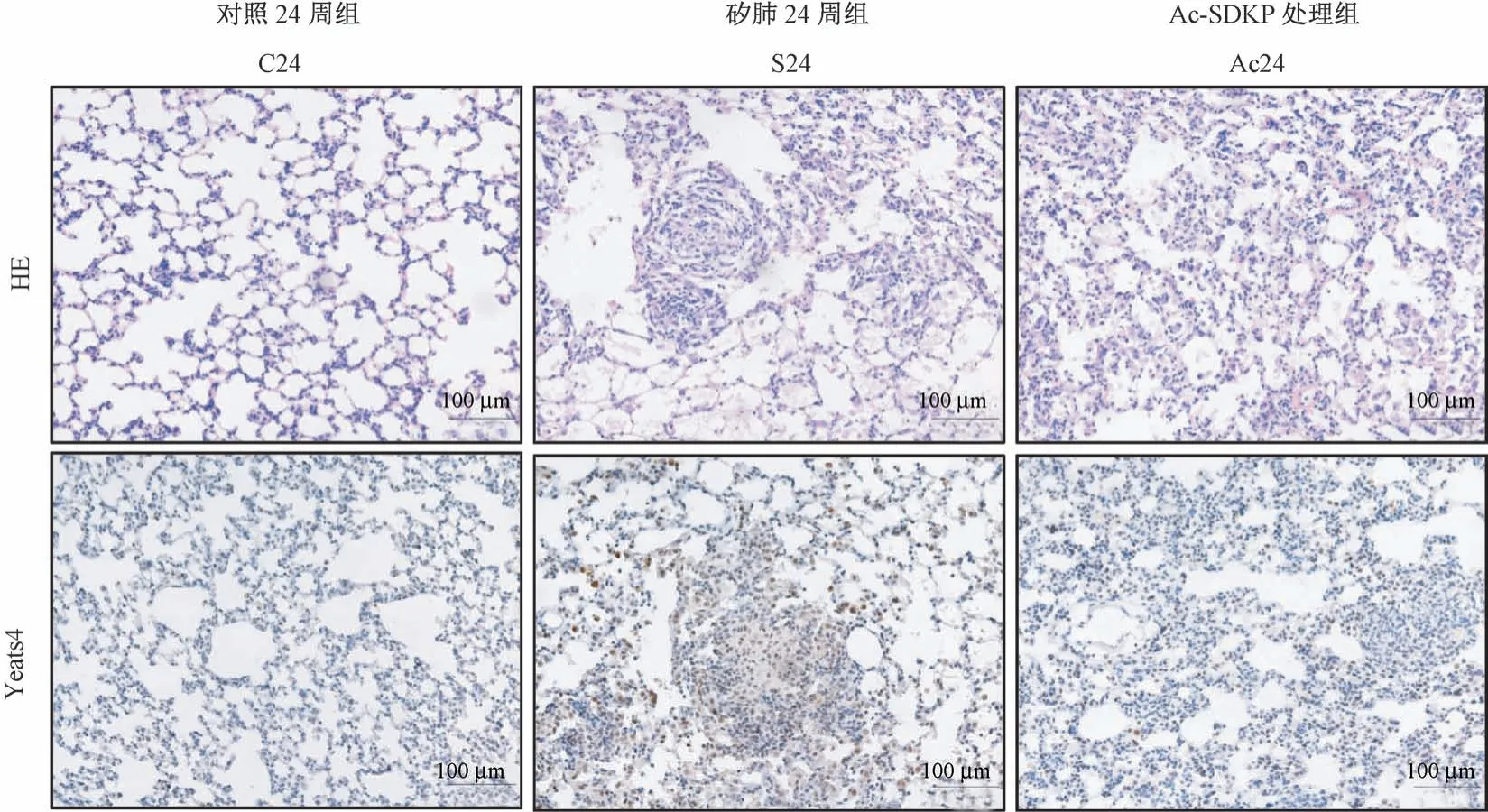
图5 Ac-SDKP 抑制染尘大鼠肺组织中YEATS4 的高表达Figure 5 Ac-SDKP inhibits the increasing level of YEATS4 in rats exposed to silica
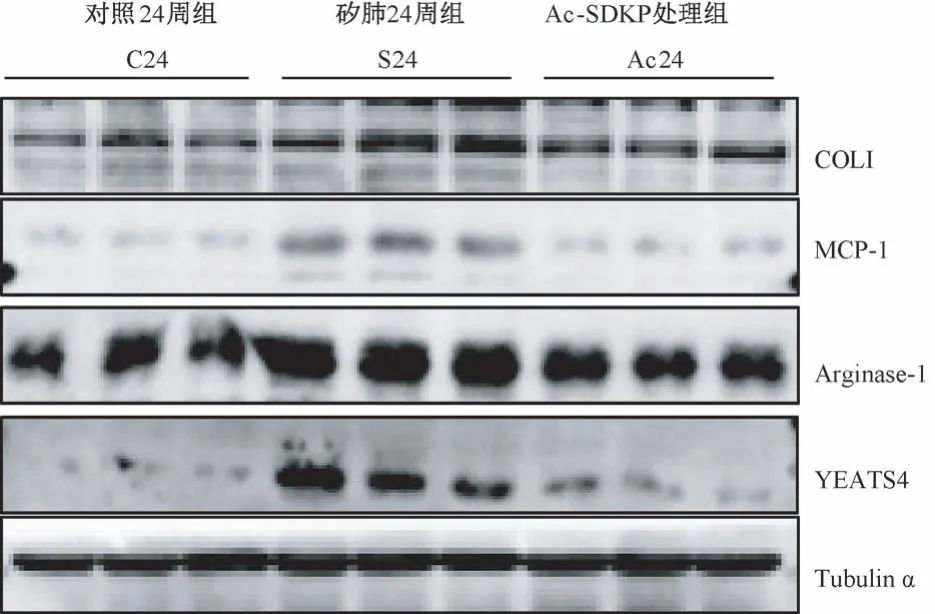
图6 Ac-SDKP 抑制染尘大鼠肺组织中YEATS4 的高表达Figure 6 Ac-SDKP inhibits the increasing level of YEATS4 in rats exposed to silica
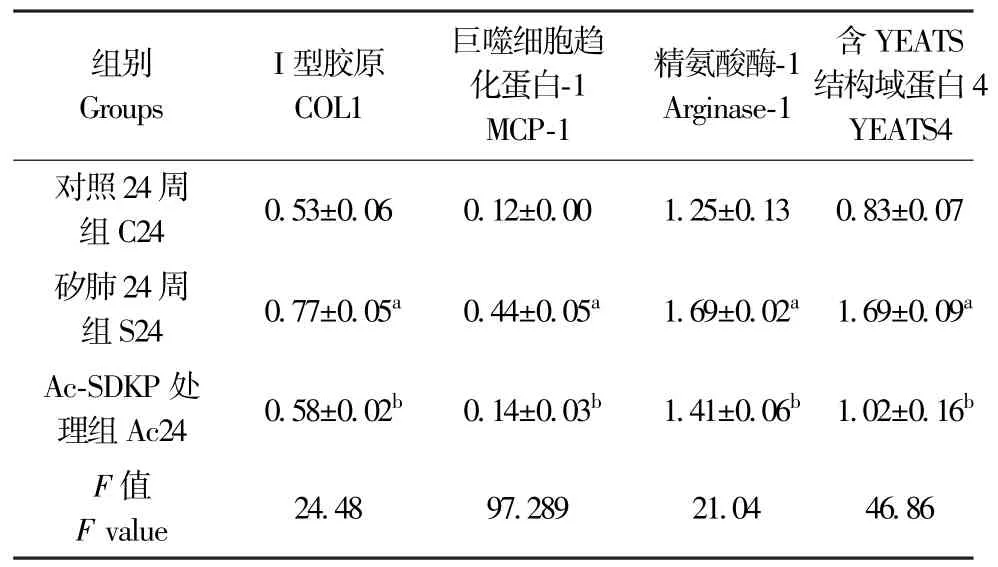
表5 Ac-SDKP 抑制染尘大鼠肺组织中YEATS4 的高表达Table 5 Ac-SDKP inhibits the increasing level of YEATS4 in rats exposed to silica
3 讨论
课题组前期通过高通量测序技术,在实验性矽肺大鼠模型中筛选了与矽肺纤维化发生、发展有关的长链非编码RNA,而YEATS4 是其重要的靶基因之一[2]。 研究表明,YEATS 家族成员能够选择性识别组蛋白赖氨酸乙酰化位点,可促进赖氨酸巴豆酰化,在转录后修饰方面起到了重要的调节作用,主要参与染色质修饰、转录调控和DNA 修复[9-13]。 在本研究中,以实验性矽肺大鼠模型和体外SiO2诱导的RAW264.7 细胞作为研究对象,发现了YEATS4主要定位于巨噬细胞,且其高表达伴随着巨噬细胞活化指标Arginase-1、MCP-1 和纤维化指标COL I 表达的上调,提示YEATS4 可能参与了实验性矽肺进程,其主要作用可能与矽尘导致的巨噬细胞活化有关。
诸多研究显示,YEATS4 作为一种致癌基因,在肿瘤发生、发展中起到了重要的调节作用。 在非小细胞肺癌中,YEATS4 能够激活P53 信号通路促进肿瘤细胞的增殖和迁移,采用基因沉默技术则能够抑制该变化[14]。 其还可通过激活Wnt/β-连环蛋白信号促进胰腺癌和胃癌细胞迁移、增殖和侵袭[5,15]。另有研究显示,YEATS4 在固有淋巴细胞及其祖细胞中高表达,并促进向辅助固有淋巴祖细胞分化,敲除YEATS4 小鼠肺泡灌洗液中嗜酸性粒细胞浸润显著减少[16]。 在本研究中,采用基因沉默技术,下调YEATS4 后,在基础水平和SiO2诱导水平均显著下调了 Arginase-1、MCP-1 和 COL I 的表达水平,进一步提示沉默YEATS 能够抑制巨噬细胞的活化。课题组前期研究表明,Ac-SDKP 可通过抑制巨噬细胞活化、肌成纤维细胞分化和上皮间质转化进程,从而发挥拮抗矽肺纤维化的作用[17-18]。 体内外实验结果显示,予以Ac-SDKP 能够抑制矽肺大鼠和RAW264.7 细胞中 YEATS4 的表达, 同时下调Arginase-1、MCP-1 和 COL I 的表达水平,提示 Ac-SDKP 可能通过对YEATS4 的阻断,从而发挥拮抗矽肺纤维化的作用。
总之,本研究发现YEATS4 可在SiO2诱导的巨噬细胞活化中起到了重要调节作用,其可能是矽肺纤维化进展的重要环节,采用基因沉默技术或给予Ac-SDKP,均能够抑制YEATS4 表达的上调,提示其可作为抗矽肺纤维化进展的一个有效靶点。 在今后的研究工作中,将深入研究矽肺纤维化进展中YEATS4 参与组蛋白乙酰化/巴豆酰化的具体机制,为明确其功能提供进一步的理论和实验依据。
猜你喜欢
艺术启蒙(2022年11期)2022-12-06
法律方法(2022年1期)2022-07-21
基层中医药(2021年6期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
国际呼吸杂志(2020年5期)2020-03-02
老年医学与保健(2017年6期)2017-02-06
当代临床医刊(2015年6期)2015-12-16
西安交通大学学报(医学版)(2015年2期)2015-02-28
卫生职业教育(2014年14期)2014-05-16
安徽医药(2014年9期)2014-03-20
