
ARR21基因在拟南芥生长发育和离体再生过程中的功能分析
2021-06-16 07:09卫云丰张树伟程金金王兴春
山西农业科学 2021年6期
卫云丰,张树伟,程金金,王兴春
(1.山西农业大学生命科学学院,山西太谷030801;2.山西农业大学农学院,山西太谷030801)
细胞分裂素是五大经典激素之一,在植物生长发育、胁迫响应和离体再生等诸多过程中起着极其重要的调控作用[1-8]。在过去的20多年中,科学家利用模式植物拟南芥系统研究了细胞分裂素的生物合成、转运及信号转导过程。在拟南芥中发现了3种组蛋白激酶(Arabidopsis histidine kinases,AHKs)为细胞分裂素受体,分别为AHK2、AHK3和AHK4[3,9],这些受体通过磷酸化的方式将细胞分裂素信号经组氨酸磷酸转移蛋白(Arabidopsis histidine-phosphot transfer proteins,AHPs)传递至B型应答调节因子(Arabidopsis response regulators,ARRs)[4-6]。B-ARRs作为一型MYB蛋白,其N端信号区含有天冬氨酸(D)、天冬氨酸(D)和赖氨酸(K)3个保守的氨基酸残基,该结构也被称为DDK结构域,该结构域的第2个天冬氨酸具有接受磷酸基团的功能[3,9-10]。B-ARRs的DDK结构域接收到上游AHP传递来的磷酸基团后,被磷酸化激活,从而启动包括A-ARRs在内的下游靶基因的转录,进而调节植物的生长发育[3,9-12]。
拟南芥中共编码12个B-ARR,N端均有1个保 守 的 信 号 接 收 域(Receiver domain,RD),除ARR23外C端均含有典型的MYB类转录因子结构域(GARP)[13]。有研究发现,单突变体arr1、arr2、arr10、arr11和arr18等与野生型均无明显的表型差异,而arr10/12双突变体对细胞分裂素的响应变弱,arr1/10/12三突变体完全丧失了对细胞分裂素的响应且植株矮小[14],表明B-ARRs基因家族存在功能冗余[13]。进一步研究发现,B-ARRs的N端信号区的DDK结构域对C端转录激活区具有抑制作用,而过表达DDK结构域缺失突变基因(ΔDDK)会解除转录抑制,导致B-ARRs介导的细胞分裂素信号途径组成型激活,从而使转基因植物呈现典型的细胞分裂素表型[10,15-17]。
ARR21作为B型ARR家族中的一员,与ARR13具有较高的相似性[16],但其功能尚待深入研究。本试验利用雌激素诱导表达系统和T-DNA插入突变体研究了ARR21基因表达对拟南芥幼苗生长发育和离体再生的影响,旨在为深入解析ARR21基因的功能奠定基础,为利用ARR21基因提高植物离体再生和遗传转化效率奠定基础。
1 材料和方法
1.1 试验材料
研究用的野生型拟南芥(Arabidopsis thaliana)为Columbia-0(Col-0)。载体为pER10和pER8[18]。T-DNA插入突变体arr13(SALK_042719c)和arr21(SALK_005772c)由陈受宜研究员馈赠。arr13/21双突变体由arr13和arr21单突变体杂交获得。拟南芥播种在1/2 MS培养基上,培养条件为:光周期16 h光/8 h暗,光照强度80~120μmol/(m2·s),温度22℃。
1/2 MS培养基:2.3 g/LMurashige and Skoog Basal Medium w/Vitamins(PhytoTechnology Laboratories,货号M519)、2%蔗糖和0.8%的琼脂粉,pH值5.8。
雌激素诱导培养基为1/2 MS培养基添加10μmol/L的17-β雌二醇(Sigma,货号E8875)。
1.2 试验方法
1.2.1 拟南芥B-ARRs进化关系分析 从拟南芥的基因组数据库TAIR(www.arabidopsis.org)中获得B型ARR家族12个成员序列,利用MEGA 7.0软件构建系统进化树,Bootstrap参数设置为1000。
1.2.2ARR21基因过量表达载体的构建和拟南芥遗传转化 以Col-0基因组DNA作为模板,利用东洋纺(上海)生物科技有限公司的高保真DNA聚合酶KOD-plus v2以及引物ARR21XhoI和ARR21XbaI(表1)扩增ARR21,经XhoⅠ和XbaⅠ双酶切后连接pER8载体,构建雌激素诱导的pER8-ARR21过量表达载体。PCR扩增体系为DNA模板3μL,dNTP 5μL,MgSO43μL,上下游引物各1.5μL,DNA聚合酶5μL,10×Buffer 5μL。扩增程序为:94℃2 min;94℃10 s,55℃30 s,68℃1 min,30个循环;68℃10 min,4℃保存。
利用ARR21ΔDDKXhoⅠ和ARR21XbaⅠ引物扩增缺失5′端438 bp(146个氨基酸)的ARR21基因,即ARR21ΔDDK片段。将该片段利用XhoⅠ和XbaⅠ双酶切后连接雌激素诱导表达载体pER10,构建成pER10-ARR21ΔDDK载体。PCR扩增体系和扩增程序同上。

表1 引物序列
将pER8-ARR21载体及pER10-ARR21ΔDDK载体通过电激法转入农杆菌GV3101中,并利用农杆菌介导的花序浸染法[19]将其转入Col-0野生型中;进行转化后,将获得的拟南芥种子种植于筛选培养基中,并施加筛选压力,筛选符合3∶1分离比的抗性拟南芥植株,获得转化成功突变体。
1.2.3 RNA提取和实时定量PCR反应 拟南芥总RNA的提取、cDNA第1链合成和实时荧光定量PCR(real-time quantitative PCR,RT-qPCR)均采用宝日医生物技术(北京)有限公司的试剂盒。其中,RNA提取采用柱式法植物RNA小量提取试剂盒(货号9769),cDNA第1链合成采用PrimeScriptTMRT reagent Kit(货号RR047A),RT-qPCR采用TB GreenRPremix Ex TaqTMⅡ试剂盒(货号RR820A),所有操作均严格按照试剂盒说明书进行。RT-qPCR所用的ARR21基因特异引物为ARR21qF和ARR21qR,内参基因ACTIN7(AT5G09810)的引物为ACT7qF和ACT7qR。采用2-ΔΔCt法计算相对表达量。
1.2.4 拟南芥离体再生 拟南芥离体再生试验以弱光下培养的下胚轴为外植体,采用两步法进行[20]。
2 结果与分析
2.1 ARR21基因的系统进化分析

拟南芥基因组中共有12个B-ARRs基因,分别为ARR1、ARR2、ARR10、ARR11、ARR12、ARR13、ARR14、ARR18、ARR19、ARR20、ARR21和ARR23。系统进化树结果表明,12个B-ARRs聚为3个亚类,其 中,ARR1、ARR2、ARR10、ARR11、ARR12、ARR14和ARR18为第Ⅰ亚类,ARR13、ARR21和ARR23为第Ⅱ亚类,ARR19和ARR20为第Ⅲ亚类。其中,ARR23序列较短,仅有信号接收域RD,无MYB结合域(GRAP)(图1-A),同一亚类中的ARR13和ARR21亲缘关系较近(图1-B),预测其可能具有相同或相似的功能。
2.2 过量表达拟南芥ARR21基因对幼苗生长发育的影响
为了研究ARR21基因在拟南芥生长和发育过程中的功能,构建了ARR21基因过量表达拟南芥pER8-ARR21(图2-A),在MS培养基和雌激素诱导培养基生长情况结果显示,过表达植株在无诱导剂的培养基上均生长良好,而在雌激素诱导培养基上生长发育严重受阻。其中,24号株系长出真叶,生长发育受阻较轻(图2-C);14号和31号株系生长发育受阻较重,且31号受阻表型最为严重,根系较短,且没有真叶长出(图2-B、D)。RT-qPCR结果表明,野生型拟南芥中ARR21基因几乎不表达,而在过表达的转基因拟南芥中,24号植株的表达量最低,31号株系中表达量最高,这与其生长发育受阻表型是一致的(图2-E)。由此预测,ARR21基因过量表达抑制了拟南芥的生长发育。

2.3 ARR21信号接收域缺失对其功能的影响
B-ARRN端含有负调控功能的DDK结构域。为研究该结构域的功能,构建了删除N端160个氨基酸残基的雌激素诱导表达载体pER10-ARR21ΔDDK,结果显示,与pER8-ARR21转基因幼苗表现一致,过量表达缺失信号接收域的ARR21ΔDDK基因抑制了拟南芥幼苗的生长发育(图3-A、B),且这种生长发育抑制表型与ARR21ΔDDK基因的表达量相关(图3-C)。
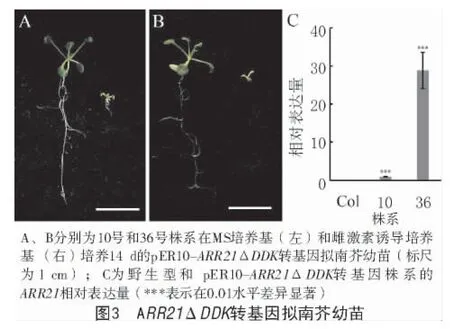
2.4 ARR21基因功能缺失对拟南芥生长发育的影响

为了进一步了解ARR21基因在拟南芥生长发育过程中的功能,研究了ARR21功能缺失突变体arr21的表型,在arr21突变体中,T-DNA插入在第5个外显子中,导致该基因功能完全丧失(图4-A)。在本试验条件下,arr21突变体与Col-0野生型无明显差异,表明ARR21基因功能缺失对拟南芥生长发育无影响(图4-B、C)。由于ARR21基因与ARR13基因有较高的相似性(82%),可能存在功能冗余。为此,进一步分析了arr13及arr13/21双突变体的表型,结果表明,arr13和arr13/21突变体幼苗与野生型Col-0也无明显差异(图4-D、E)。
2.5 拟南芥ARR21基因过表达和缺失对离体再生的影响
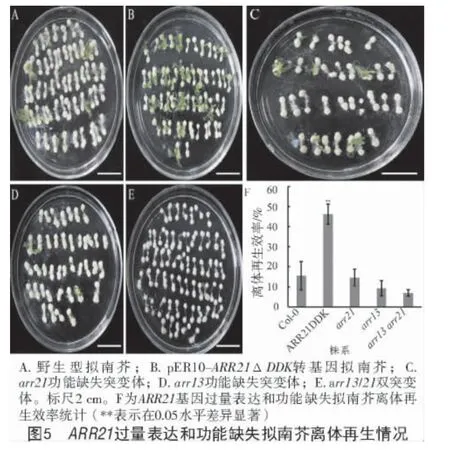
细胞分裂素在植物离体器官再生过程中起着极其重要的作用,进一步分析了ARR21基因过表达和功能缺失对拟南芥离体再生的影响,结果显示(图5),过表达ARR21基因显著提高了离体再生能力,但arr21功能缺失对离体再生无显著影响;此外,arr13单突变体和arr13/21双突变体离体再生效率均低于野生型。
3 结论与讨论
B-ARR为细胞分裂素信号途径的关键枢纽,探究其生物学功能对于阐明细胞分裂素途径的调控机制有重要意义。拟南芥基因组中共编码12个B-ARRs,其中,ARR1、ARR10、ARR12等的功能已有了清晰的认识[5]。本研究表明,B型ARR基因ARR21过量表达抑制了幼苗的生长发育,促进了离体再生;虽然arr21单突变体对离体再生无显著影响,但arr13/21双突变体离体再生能力低于野生型。
有研究发现,由于DDK结构域对C端转录激活域的抑制,组成型表达DDK结构域缺失的B型ARR基因会导致细胞分裂素信号激活,抑制幼苗生长发育,导致难以获得转基因后代[10,15-18,21]。本研究发现,过表达DDK缺失突变体在雌激素诱导培养基上,幼苗生长发育异常,无法收获转基因种子,与前人对组成型表达ARR21ΔDDK的研究结果完全一致。值得注意的是,该结果与过量表达ARR21全长基因的研究结果相似,二者突变体幼苗均出现典型的细胞分裂素表型,即幼苗生长发育严重受阻[15];尽管如此,arr21单突变体和arr13/21双突变体幼苗生长发育完全正常,推测ARR21基因可能不参与营养生长阶段的调控,这与其表达谱一致[13]。
细胞分裂素是植物离体器官再生的关键决定因素。本研究发现,arr21单基因突变对拟南芥离体再生无显著影响,而arr13/21双基因突变后离体再生效率显著降低,表明B型ARR参与调控拟南芥离体再生,且存在明显的功能冗余;相反,结构缺失导致的ARR21基因组成型表达可以显著提高离体再生效率。因此,ARR21基因有望用于难以组培的植物,以提高其遗传转化效率。
本研究初步解析了ARR21基因在植物生长发育和离体再生过程中的功能,但其具体调控机制有待进一步研究。随着高通量测序技术的发展,未来可以利用基于高通量测序的ChIP-Seq等技术,筛选和鉴定ARR21的下游靶基因,阐明其转录调控机制。
猜你喜欢
作物学报(2022年2期)2022-11-06
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
江苏农业科学(2017年19期)2017-11-22
中国医学创新(2017年5期)2017-03-16
奥秘(2016年10期)2016-12-17
考试周刊(2016年82期)2016-11-01
中学教学参考·理科版(2016年2期)2016-04-05
中国青年(1991年4期)1991-09-27
