
雅鲁藏布江异齿裂腹鱼不同群体的形态差异
2021-06-08 03:41魏源博张桂蓉李雷马波马徐发魏开建
华中农业大学学报(自然科学版) 2021年3期
魏源博,张桂蓉,李雷,马波,马徐发,魏开建
1.华中农业大学水产学院/水产养殖国家级实验教学示范中心/农业农村部淡水生物繁育重点实验室,武汉430070; 2. 中国水产科学研究院黑龙江水产研究所,哈尔滨 150070
异齿裂腹鱼(Schizothoraxo’connori)隶属鲤形目(Cypriniformes)、鲤科(Cyprinidae)、裂腹鱼亚科(Schizothoracinae)、裂腹鱼属(Schizothorax),是我国西藏特有的高原冷水性鱼类,分布于雅鲁藏布江中上游干支流及其附属水体中[1]。异齿裂腹鱼是随着青藏高原的急剧隆升而特化的裂腹鱼类群之一[2],是雅鲁藏布江的主要优势鱼类和珍稀物种,具有重要经济和生态价值,在鱼类进化和动物地理学等研究中具有重要科学价值。近年来,由于过度捕捞、外来物种入侵、涉水工程建设等人类活动的影响,其自然种群数量急剧下降,个体小型化明显[3-4],急需加强其现有种群资源保护的研究。
目前,国内学者对异齿裂腹鱼生物学特征[5-6]、资源状况[3]等作了研究,并开展了种群资源评估[7]、增殖放流试验与幼鱼标记方法[8-9]等研究,以保护其种群资源。探明珍稀鱼类不同群体的潜在遗传结构、遗传分化、形态差异等种质特征,是科学开展亲本遗传管理、人工繁育、增殖放流等种质资源养护的重要前提之一。武云飞等[1]首次报道了雅鲁藏布大峡谷(后文简称大峡谷)上游流域和下游支流中异齿裂腹鱼在须长、下咽骨形态等少量形态特征上存在明显差异。随后国内学者基于单个mtDNA序列[10]或将mtDNA序列与微卫星标记相结合[11-12],对雅鲁藏布江异齿裂腹鱼的亲缘地理学和群体遗传学进行了研究,揭示异齿裂腹鱼群体存在显著遗传结构,大峡谷上游流域和下游支流等群体间存在明显遗传分化。而关于异齿裂腹鱼不同地理群体的形态差异还未见研究报道。为此,笔者通过测量鱼体常规和框架形态性状,运用主成分分析、聚类分析、判别分析等方法比较了雅鲁藏布江异齿裂腹鱼10个群体的形态差异,建立了不同群体的形态判别函数,旨在为雅鲁藏布江异齿裂腹鱼群体识别、种质资源保护和管理提供基础资料。
1 材料与方法
1.1 材 料
异齿裂腹鱼10个群体样本于2018年4-6月采自雅鲁藏布江流域拉孜县、日喀则市、桑日县、加查县、朗县、米林县及派镇的干流江段,中游支流拉萨河的墨竹工卡县江段,以及大峡谷下游支流帕隆藏布江的通麦镇、卡达桥江段,样本量和鱼体规格等见表1。
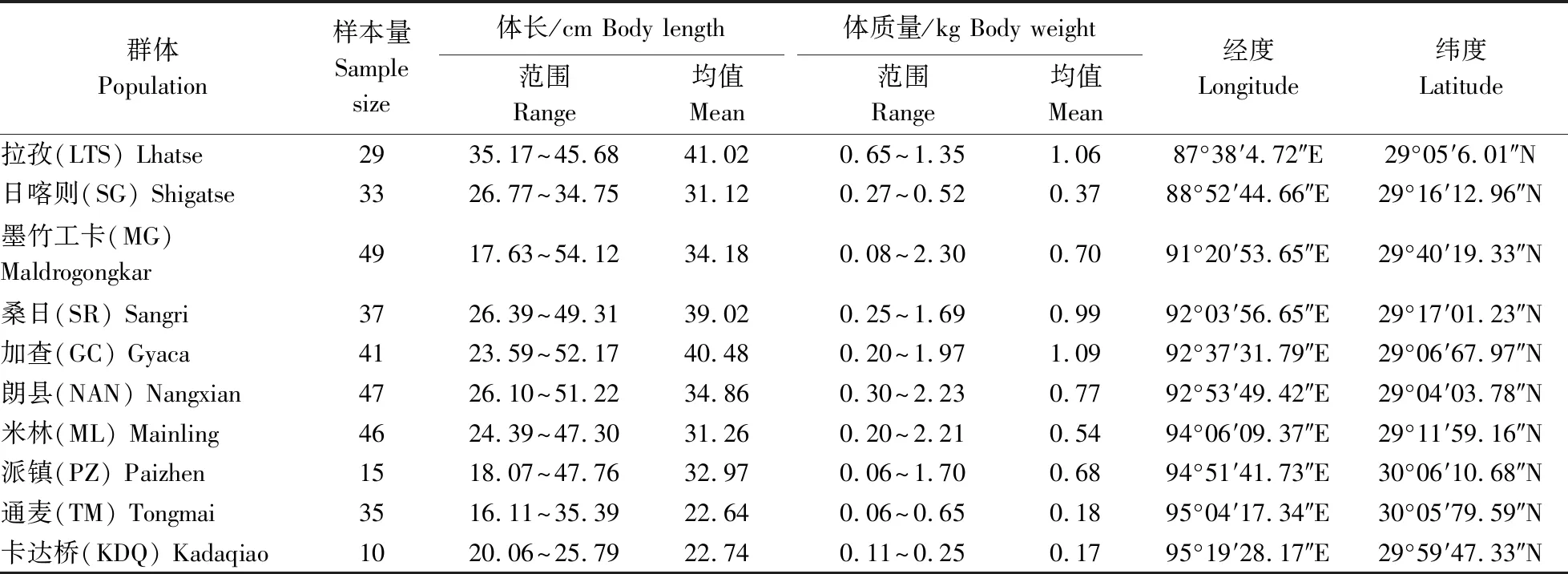
表1 异齿裂腹鱼10个群体的样本量、体长、体质量和采样地点Table 1 Sample size,body length,body weight and sampling locations of ten populations of S. o’connori
1.2 形态性状测量
对鱼体的两类形态性状进行测量。一类是常规形态性状[13-14],先用数显游标卡尺(精确度0.01 mm)测量鱼体的眼径(ED)、眼间距(ID)、口裂宽(MB)、体宽(BW)、胸鳍起点间距(DOP)、腹鳍起点间距(DOV),然后以直尺为参照拍摄鱼体左侧数码照片,采用Photoshop软件度量工具测量鱼体的全长(TL)、体长(BL)、头长(HL)、吻长(SL)、体高(BD)、尾柄长(CPL)、尾柄高(CPD)等13个常规形态性状。另一类是框架形态性状[13,15-16],采用11个解剖学坐标点进行框架测量(图1),坐标点之间的直线距离用D1-2、D1-3、…、D8-10等表示,采用Photoshop软件共测量了20个框架形态性状(精确度0.1 mm)。
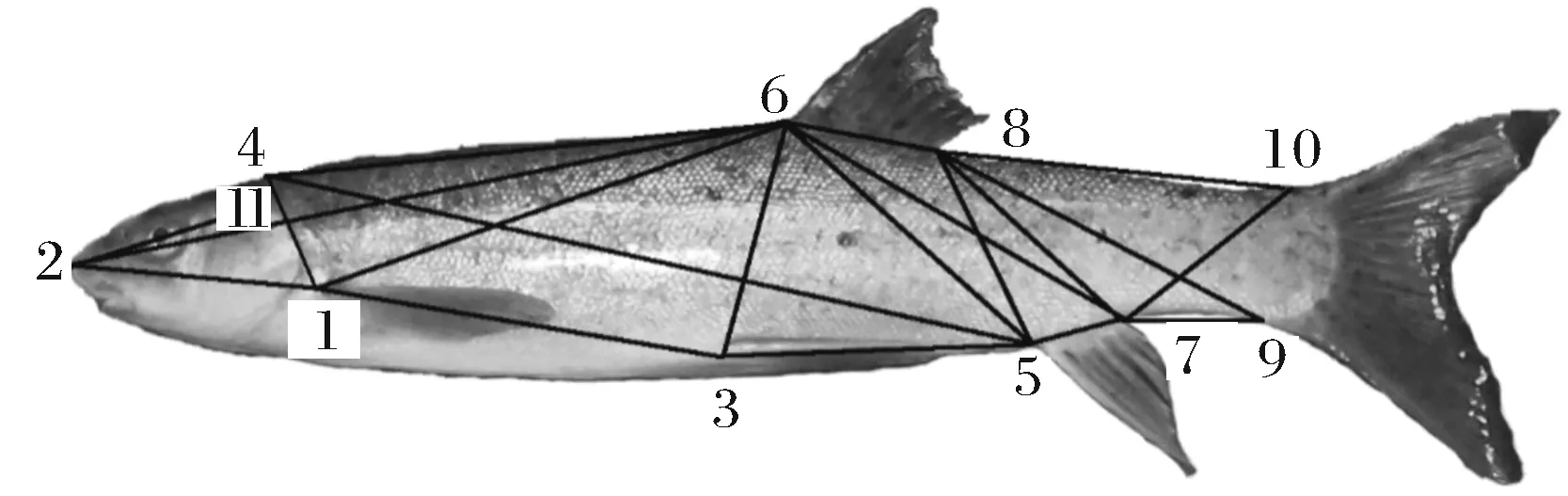
11个解剖学坐标位点 Eleven anatomical landmark points. 1. 胸鳍起点 Origin of pectoral fin; 2. 吻端 Tip of snout; 3.腹鳍起点 Origin of ventral fin; 4. 额部有鳞部最前端 Most anterior end of scales on skull; 5. 臀鳍起点 Origin of anal fin; 6. 背鳍起点 Origin of dorsal fin; 7. 臀鳍基部末端 Terminus of anal fin base; 8.背鳍基部末端 Terminus of dorsal fin base; 9. 尾鳍腹部起点 Ventral origin of caudal fin; 10.尾鳍背部起点 Dorsal origin of caudal fin; 11. 鳃盖基部 Operculum base.
1.3 数据分析
为了消除鱼体大小差异对形态性状观测值的影响,将每一个体的形态测量数据与其体长(BL)的比值作为形态度量分析的性状[14-17],共有32个比例性状(体长除外)用于分析。采用Statistica 7.0软件对32个比例性状数据进行主成分分析、聚类分析和判别分析,其中聚类方法为欧氏距离的最短距离法,判别分析采用逐步判别法[18]。采用单因素方差分析和Newman-Keuls检验比较不同群体之间形态比例性状的差异。
2 结果与分析
2.1 主成分分析
主成分分析提取了6个主成分(PC1~PC6),其方差累积贡献率为74.23%(表2)。从32个比例性状对6个主成分的负荷值可以看出,PC1贡献率最高(28.22%),主要与全长、吻长、眼径、D1-2、D1-3、D1-6、D2-11、D3-5、D4-5、D4-6、D7-9、D7-10、D8-9、D8-10对体长(BL)的14个比例性状有关,主要反映吻端至尾柄端鱼体水平轴向的形态变化;PC2贡献率次之(19.09%),主要与体高、尾柄高、体宽、胸鳍起点间距、腹鳍起点间距、D3-6、D5-6、D5-8、D6-7、D7-8对BL的10个比例性状有关,主要反映鱼体垂直轴向躯干部高度、宽度以及尾柄高度的形态变化;PC3和PC4的贡献率分别为9.02%和8.59%,各比例性状的负荷值均小于0.600;PC5(贡献率5.25%)主要与D6-8/BL有关,主要反映背鳍基部长度的变化;PC6(贡献率4.06%)主要与ID/BL有关,主要反映头部眼间距的变化。

表2 异齿裂腹鱼10个群体32个比例性状对6个主成分的负荷值及主成分的贡献率Table 2 Loading of six principal components for thirty-two characters of ten S. o’connori populations and their contribution rate
以贡献率最高的PC1和PC2构建因子得分散布图(图2)。通麦(TM)和卡达桥(KDQ)2个群体彼此重叠,其散布区位于PC1轴上因子得分>0.75、PC2轴上因子得分<0.25的区域,并与大峡谷上游8个群体几乎完全分开,存在明显差异。在大峡谷上游8个群体中,日喀则(SG)与米林(ML)、墨竹工卡(MG)群体的散布区相邻且有较多重叠。拉孜(LTS)、桑日(SR)、加查(GC)、朗县(NAN)4个群体之间彼此重叠,它们与SG群体散布区相邻但重叠很少。MG、ML和派镇(PZ)3个群体的散布区较分散。
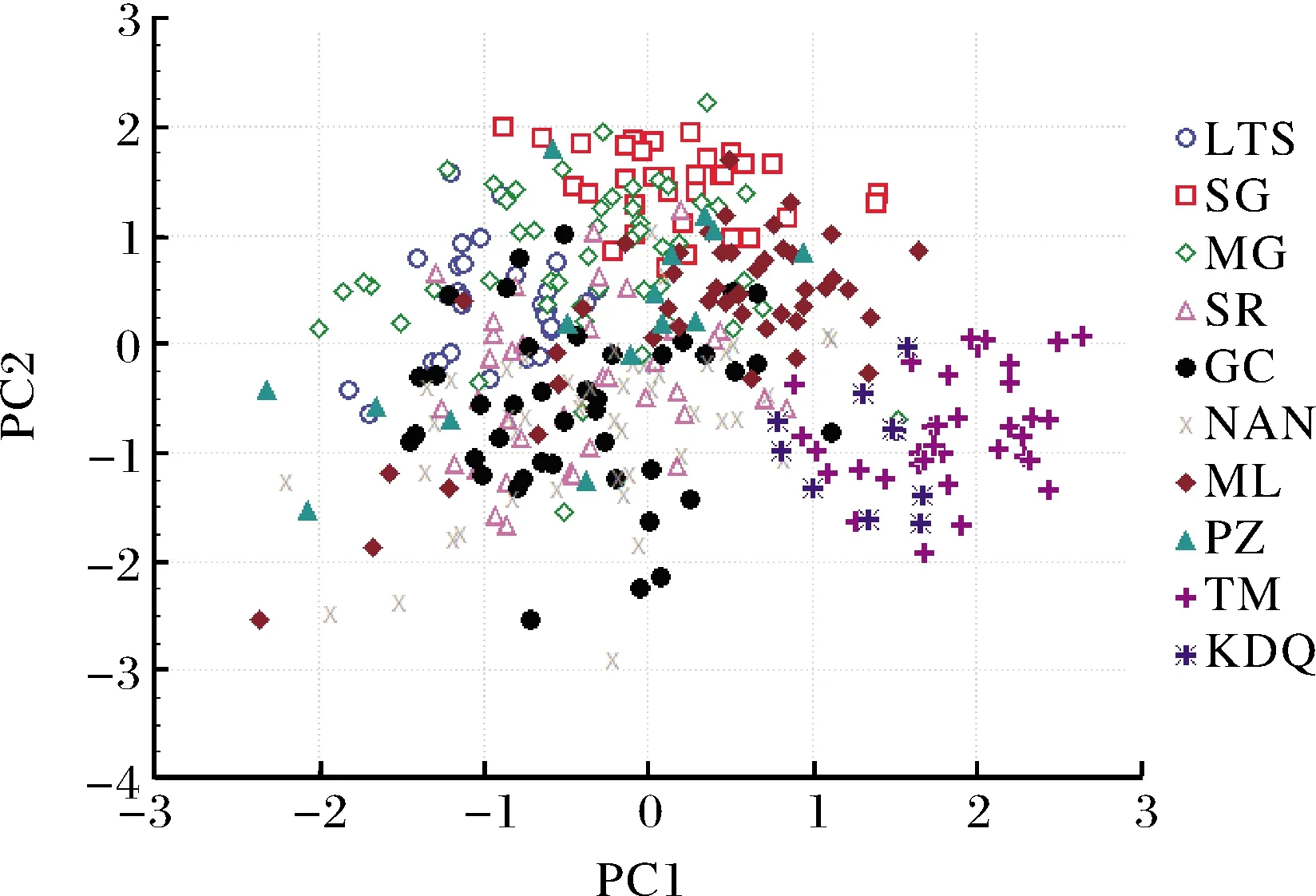
图2 异齿裂腹鱼10个群体的第一、第二主成分散布图Fig.2 Scatter diagram for the first principalcomponent (PC1) and the second principalcomponents (PC2) of ten S. o’connori populations
2.2 群体间形态差异的方差分析
单因素方差分析表明,10个群体之间32个比例性状均存在显著性差异(P<0.05),群体之间各比例性状平均数的多重比较结果见表3。
从表3可见,在12个常规形态性状中,TM群体TL/BL和ED/BL均值最大,KDQ群体次之,这2个群体的TL/BL和ED/BL都显著大于大峡谷上游8个群体(P<0.05)。同时,TM群体的HL/BL、SL/BL、CPD/BL均值也最大,显著大于其余9个群体(P<0.05);KDQ群体ID/BL均值最小,显著小于其余9个群体(P<0.05)。TM和KDQ 2个群体的其他6个常规性状与大峡谷上游一部分群体间无显著差异。在大峡谷上游8个群体中,LTS群体有4个性状(TL/BL、HL/BL、SL/BL、CPD/BL)均值最小;SG群体有2个性状(ID/BL、CPL/BL)均值最大,5个性状(MB/BL、BD/BL、BW/BL、DOP/BL、DOV/BL)均值最小;MG和SR群体分别在CPL/BL、ED/BL性状的均值最小;SR和GC群体分别在BW/BL、MB/BL性状的均值最大;NAN群体在3个性状(BD/BL、DOP/BL、DOV/BL)的均值最大。在大峡谷上游8个群体之间,12个常规形态性状分别呈现不同程度的差异(表3)。
在20个框架形态性状中,TM与KDG 2个群体之间仅有3个性状(D1-2/BL、D2-11/BL和D3-6/BL)存在显著性差异(P<0.05)。TM群体3个性状(D1-2/BL、D1-4/BL、D2-11/BL)均值最大,都显著大于大峡谷上游8个群体(P<0.05)。TM(或KDG)群体5个性状(D1-6/BL、D2-6/BL、D3-5/BL、D4-5/BL、D4-6/BL)均值最小,2个群体的5个性状都显著小于大峡谷上游8个群体(P<0.05)。TM(或KDG)群体6个性状(D5-6/BL、D5-8/BL、D6-7/BL、D7-8/BL、D8-9/BL、D8-10/BL)均值最大,2个群体的6个性状都显著大于大峡谷上游8个群体(P<0.05)。在大峡谷上游8个群体中,LTS群体有4个性状(D1-6/BL、D2-6/BL、D4-5/BL、D4-6/BL)均值最大,D2-11/BL均值最小;SG群体有7个性状(D1-3/BL、D3-6/BL、D5-6/BL、D5-8/BL、D6-7/BL、D6-8/BL、D7-8/BL)均值最小;MG群体的D6-8/BL均值最大,2个性状(D8-9/BL、D8-10/BL)均值最小;SR群体有2个性状(D1-4/BL、D7-10/BL)均值最小;GC群体有2个性状(D3-5/BL、D5-7/BL)均值最大,D7-9/BL均值最小;NAN群体的D1-2/BL均值最小,在2个性状(D1-3/BL、D3-6/BL)均值最大。大峡谷上游8个群体之间20个框架形态性状表现出不同程度的差异(表3)。

表3 异齿裂腹鱼10个群体间32个比例性状平均数的多重比较Table 3 Multiple comparison for averages of thirty-two proportional characters among ten populations of S. o’connori
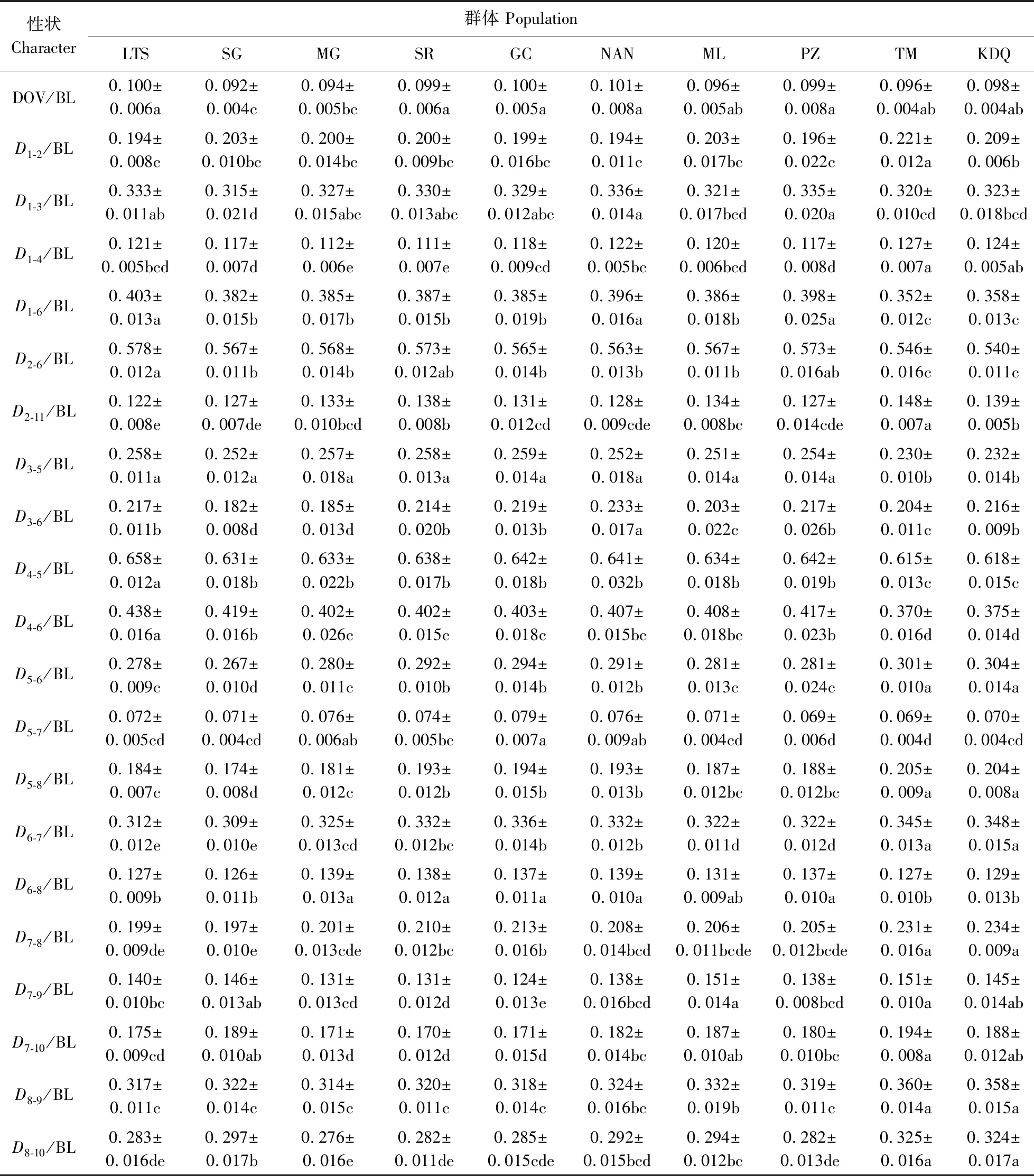
续表3 ContinuedTable 3
总体上,在32个比例性状中,TM和KDQ 2个群体与大峡谷上游8个群体之间的显著差异性状数(ND)分别为25~30和22~25(TM、KDQ群体与ML之间的ND=19)(P<0.05)。在大峡谷上游8个群体中,SR与GC之间的ND最少(4);LTS群体与ML之间的ND最多(22),与SG、MG之间的ND次之(17),与其余4个群体之间的ND相对较少;SG群体与ML、MG之间的ND较少(13~15),与其余5个群体之间的ND较多(17~21)。
2.3 聚类分析
基于32个比例性状对10个群体进行聚类分析,获得群体之间的平均欧氏距离范围为0.087(TM/KDQ)~0.172(TM/LTS)。其中,TM和KDQ 2个群体与大峡谷上游8个群体之间的欧氏距离较大(0.126~0.172);NAN群体与SG和MG之间的距离(0.133和0.132)、GC与SG群体之间的距离(0.128)也较大,其余群体之间的距离较小(0.102~0.124)。基于欧氏距离构建了10个群体的聚类图(图3),大峡谷下游支流TM和KDQ 2个群体形态相近而聚为一支,而大峡谷上游8个群体聚为另一支。在大峡谷上游8个群体这一分支中,SR与GC群体形态相似、先聚在一起,再依次与LTS、PZ、NAN群体聚为一个组群;而SG、ML、MG 3个群体聚为另一个组群。
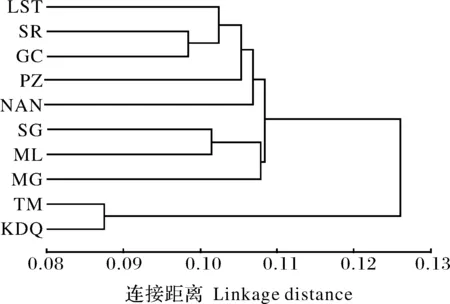
图3 异齿裂腹鱼10个群体的最短距离聚类图Fig.3 Single-linkage clustering dendrogram for ten populations of S. o’connori
2.4 判别分析
以32个比例性状为变量进行逐步判别分析,除了D1-2/BL(X13)、D4-5/BL(X21)、D5-8/BL(X25)、D6-8/BL(X27)和D7-8/BL(X28)5个变量外,其余27个变量进入判别函数模型,建立了10个群体的判别函数。
(1)拉孜群体。YLTS=3002.369X1+732.151X2+7047.822X3-1949.241X4+3499.036X5-1870.299X6+1387.652X7+3625.944X8-1624.360X9+519.739X10+1932.954X11-2105.085X12+3239.348X14+1171.691X15+1008.424X16+5411.707X17-470.030X18+4523.303X19-2775.975X20+196.919X22-58.460X23+2062.115X24+5514.266X26+2302.522X29+1531.528X30+196.629X31+1225.034X32-6598.516
(2)日喀则群体。YSG=2989.667X1+643.754X2+7358.882X3-1281.108X4+3923.461X5-2011.638X6+1339.438X7+3676.911X8-1287.225X9+353.823X10+1765.852X11-2049.128X12+3264.665X14+1072.897X15+1097.811X16+5399.343X17-490.341X18+4487.499X19-2869.418X20+165.807X22-34.800X23+2162.706X24+5529.401X26+2158.717X29+1647.495X30+179.736X31+1286.859X32-6626.670
(3)墨竹工卡群体。YMG=2981.756X1+792.050X2+6915.334X3-1976.734X4+3340.831X5-1910.287X6+1294.349X7+3583.220X8-1158.271X9+370.989X10+1834.297X11-2092.989X12+3343.405X14+1029.428X15+1105.368X16+5394.602X17-184.436X18+4484.203X19-2884.122X20+70.116X22+23.154X23+2178.918X24+5563.006X26+2219.596X29+1569.653X30+199.959X31+1190.729X32-6577.920
(4)桑日群体。YSR=3001.853X1+784.068X2+7229.501X3-2290.213X4+3268.801X5-1629.678X6+1347.496X7+3624.255X8-1330.922X9+544.721X10+1767.158X11-2208.857X12+3308.798X14+826.075X15+1026.208X16+5494.513X17-280.854X18+4507.298X19-2772.519X20+106.457X22+24.307X23+1992.492X24+5627.834X26+2280.207X29+1470.125X30+190.271X31+1243.161X32-6669.560
(5)加查群体。YGC=3022.222X1+772.190X2+7129.981X3-2019.010X4+2989.977X5-1315.493X6+1345.486X7+3636.092X8-1465.768X9+428.654X10+1819.289X11-2133.470X12+3307.839X14+1042.368X15+1055.139X16+5415.418X17-470.365X18+4443.698X19-2724.452X20+112.696X22+8.163X23+2123.102X24+5611.999X26+2084.288X29+1581.902X30+215.544X31+1244.319X32-6644.989
(6)朗县群体。YNAN=3055.395X1+744.379X2+6909.057X3-1883.445X4+2991.248X5-1359.882X6+1385.155X7+3642.168X8-1330.583X9+344.176X10+1834.385X11-2174.764X12+3330.055X14+1178.572X15+1180.788X16+5285.327X17-358.094X18+4410.660X19-2697.382X20+89.154X22+12.095X23+2115.027X24+5553.574X26+2127.554X29+1564.525X30+233.389X31+1254.994X32-6667.308
(7)米林群体。YML=2983.518X1+714.391X2+7038.261X3-1516.378X4+2970.776X5-1432.526X6+1307.074X7+3626.569X8-1201.976X9+461.619X10+1711.132X11-2068.102X12+3319.962X14+1143.675X15+1150.670X16+5372.931X17-277.712X18+4479.056X19-2810.657X20+127.260X22+47.399X23+2094.169X24+5551.864X26+2252.351X29+1576.706X30+256.352X31+1245.163X32-6668.613
(8)派镇群体。YPZ=3038.265X1+744.877X2+6924.664X3-2012.521X4+3040.877X5-1684.007X6+1406.175X7+3725.842X8-1335.267X9+412.869X10+1811.571X11-2032.629X12+3327.985X14+1074.745X15+1078.089X16+5400.738X17-326.087X18+4480.470X19-2787.921X20+113.527X22-81.855X23+1913.674X24+5625.300X26+2125.409X29+1566.582X30+238.636X31+1189.939X32-6648.842
(9)通麦群体。YTM=3029.269X1+715.645X2+7055.980X3-1198.808X4+2647.934X5-1651.777X6+1217.272X7+3567.004X8-652.596X9+149.849X10+1743.001X11-1903.262X12+3426.079X14+1244.540X15+1154.638X16+5273.629X17-69.383X18+4268.999X19-2703.402X20+68.565X22+240.262X23+2064.480X24+5520.375X26+2051.476X29+1514.430X30+368.857X31+1378.797X32-6745.938
(10)卡达桥群体。YKDQ=3042.155X1+671.836X2+7001.299X3-1236.848X4+2458.777X5-1575.414X6+1287.672X7+3585.757X8-1013.749X9+216.647X10+1754.327X11-1837.031X12+3352.578X14+1182.510X15+1129.104X16+5222.019X17-168.748X18+4209.209X19-2638.592X20+91.632X22+180.261X23+2036.048X24+5541.165X26+2000.552X29+1494.862X30+358.715X31+1373.484X32-6634.083
将每个个体的比例性状数据代入上述公式进行判别分析,群体的判别准确率为53.33%~100.00%,总的判别准确率为88.60%。其中SG群体的判别准确率为100.00%,LTS、MG和TM群体的判别准确率大于95.00%。10个群体的预测分类及判别准确率详情见表4。
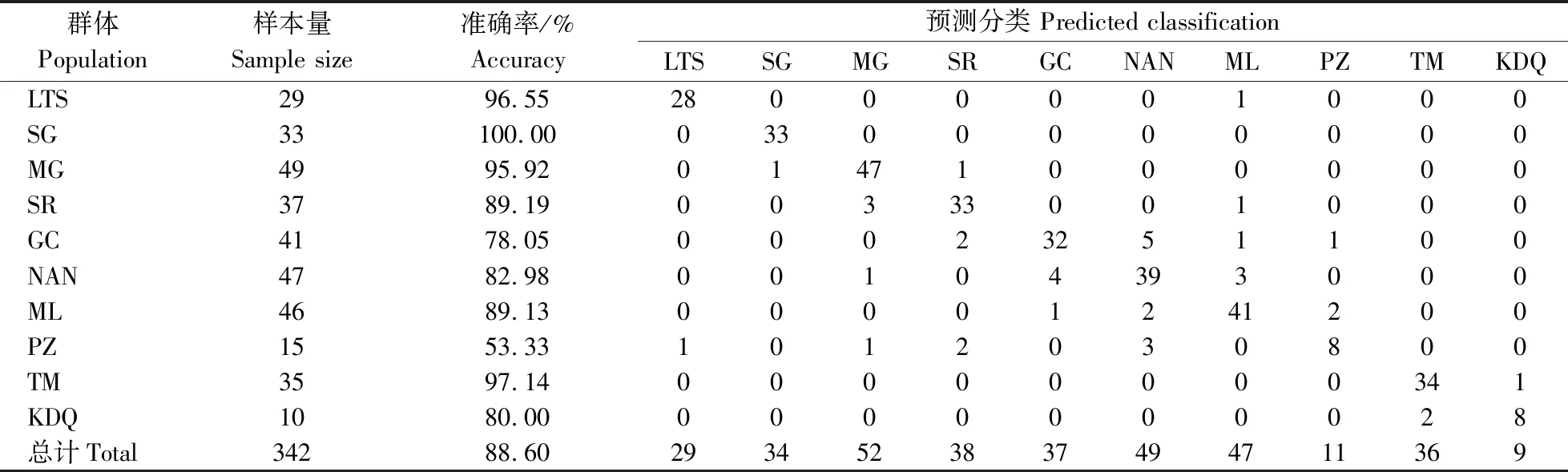
表4 判别函数对异齿裂腹鱼10个群体的预测分类及准确率Table 4 Predicted classifications of discriminant functions and their percentages of accuracy for ten populations of S. o’connori
3 讨 论
群体分化是进化生物学的重要研究内容,而形态发生是物种水平上鱼类群体鉴定的重要标准[19]。鱼类形态特征受到遗传因子和环境因素的影响,群体间的形态差异是生物多样性的外在表现[17,20-21]。鱼类形态的多元统计分析信息更加丰富、有效,是对传统形态分类研究的补充和完善[22],因此,形态度量学在水产动物的物种识别[13,18,22]、鱼类群体形态分化及生物多样性保护研究[14-17,20-21]等方面得到了广泛应用。本研究基于32个常规和框架形态性状,采用主成分分析、聚类分析、判别分析和单因素方差分析等方法对雅鲁藏布江异齿裂腹鱼10个群体的形态差异进行了比较分析。
主成分分析可将多个相互关联的指标转换为新的、个数较少、相互独立的综合指标,以反映群体间的主要形态差异[14-15,18]。本研究的主成分分析提取了6个主成分,其方差累积贡献率74.23%,未达到累积贡献率≥85%的要求,说明难以用几个相互独立的主成分来全面地概括异齿裂腹鱼不同群体间的形态差异[14-15,22]。PC1主要与全长/体长、吻长/体长、眼径/体长、D1-2/体长、D1-3/体长等14个比例性状有关,主要反映吻端至尾柄端鱼体水平轴向的差异;PC2主要与体高/体长、尾柄高/体长、体宽/体长、胸鳍起点间距/体长、腹鳍起点间距/体长等10个比例性状有关,主要反映鱼体垂直轴向躯干部高度、宽度以及尾柄高度的差异。PC1与PC2的散布图显示,大峡谷下游支流通麦(TM)和卡达桥(KDQ)2个群体差异很小,它们与大峡谷上游8个群体形态分化明显,且在PC1轴上差异更大。拉孜(LTS)、桑日(SR)、加查(GC)、朗县(NAN)4个群体间形态差异较小,它们与日喀则(SG)群体差异较大。方差分析表明,在32个比例性状中,TM和KDQ与大峡谷上游8个群体之间有显著差异的性状数相对较多;在大峡谷上游8个群体中,SG群体与ML、MG之间的显著差异性状数较少,与其余5个群体间的显著差异性状数较多,而SR、GC、NAN、LTS群体之间显著差异性状数较少。方差分析显示的群体间形态差异与主成分分析结果基本一致。此外,与大峡谷上游8个群体相比,TM和KDQ 2个群体的全长、眼径、背鳍基部-臀鳍基部之间的斜向高度较大、尾柄部较长,而躯干部较短(P<0.05);同时,TM群体的头长和吻长较长、眼间距较小、尾柄较高(P<0.05)。因此,头部、躯干部和尾柄部的部分性状是区分TM、KDQ 2个群体与大峡谷上游8个群体的关键因素,这与其他鱼类群体间或近缘种间在头部、尾部或躯干部一些性状上存在差异的研究相似[13,16,22]。
判别分析是常用的群体鉴定方法,通过建立判别函数能较准确地进行群体判别[14-15]。本研究中,基于27个比例性状建立了10个群体的判别函数,总的判别准确率为88.60%,其中SG群体的判别准确率为100.00%,LTS、MG和TM群体的判别准确率高于95.00%,除PZ外,其余群体的判别准确率高于78.00%,判别效果较好。SR、GC、NAN、ML群体之间有少数个体相互判别错误,这说明地理相邻群体间的形态差异较小,与主成分分析结果一致。而PZ群体判别准确率最低,可能是因为其样本量较小、或与上游SR、NAN等群体存在一定的基因交流有关。
聚类分析可以根据样本多个指标的原始数据信息量化样本间的差异程度并进行初步归类,以分析群体间的相似性[13-14]。聚类分析显示,TM和KDQ 2个群体的欧氏距离最短而聚为一支,而大峡谷上游8个群体聚为另一支,2个分支间的欧氏距离较大。大峡谷上游8个群体又聚为SG/ML/MG和SR/GC/LTS/PZ/NAN 2个组群。聚类分析结果与主成分分析、方差分析的群体差异基本一致,认为大峡谷下游支流帕隆藏布江TM和KDQ 2个群体形态差异小而为一支,大峡谷上游8个群体为另一支,两者存在明显的形态分化;同时,大峡谷上游群体间也存在一定的形态差异。TM和KDQ 2个异齿裂腹鱼群体与大峡谷上游8个群体在外部形态上的明显分化,与武云飞等[1]报道的大峡谷上游和下游支流中异齿裂腹鱼在须长、下咽骨形态等少量形态特征差异相互印证,也与He等[10]基于单个mtDNA序列以及Guo等[12]基于2个mtDNA序列与微卫星标记分析揭示的大峡谷上游和下游支流中异齿裂腹鱼群体之间存在显著遗传分化的结果一致。大峡谷上游8个群体可划分2个形态差异组群,这与微卫星标记分析显示大峡谷上游异齿裂腹鱼群体包含2个遗传谱系相吻合[12],但ML群体在形态聚类中位置与遗传结构中的谱系不一致。因此,异齿裂腹鱼不同群体间的外部形态分化与其在青藏高原隆升过程中因地理隔离等造成的遗传分化有关[10,12]。此外,鱼类外部形态除了受到遗传因子的影响,也会受到栖息环境因素影响[13,16-17,20-21]。雅鲁藏布江中游江段山高坡陡,有瀑布和跌水,水流较急;而大峡谷下游支流帕隆藏布江底多滩礁、乱石,水流甚急[1],中游江段8个群体与大峡谷下游支流2个群体的异齿裂腹鱼可能因为长期隔离并适应特定栖息环境而出现表型的差异[1,13]。例如,与大峡谷上游群体相比,大峡谷下游支流TM、KDQ群体的全长和尾柄部较长,TM群体头长和吻长较长、尾柄较高,这些特征使其具有稳定、相对较强的游泳能力,这可能是对急流的适应[16,22]。这与娄晋铭等[23]对甘肃省境内的3种裂腹鱼的研究中也得出了长期地理隔绝导致种群分化的结果是一致的。
本研究对雅鲁藏布江异齿裂腹鱼10个群体的外部形态差异进行了分析,探讨了雅鲁藏布大峡谷上游和下游支流不同群体之间的形态特征、分化、判别方法以及与地理分布的关系,可为雅鲁藏布江异齿裂腹鱼群体的划分、鉴定、增殖放流、种质资源保护及其适应性进化研究提供重要的基础数据。
猜你喜欢
水产科学(2020年5期)2020-09-23
疯狂英语·新悦读(2019年11期)2019-12-18
新农民(2019年2期)2019-02-20
当代水产(2018年8期)2018-11-02
幼儿100(2017年10期)2017-05-04
中国三峡(2016年11期)2017-01-15
中学生数理化(高中版.高二数学)(2016年4期)2016-03-01
浙江理工大学学报(自然科学版)(2015年5期)2015-03-01
数学年刊A辑(中文版)(2014年4期)2014-10-30
生物进化(2014年3期)2014-04-16
