
干旱胁迫对桂叶山牵牛成熟苗生理生化的影响
2021-05-30 13:26唐庆,黄欣,秦波,李冰,林茂
广东农业科学 2021年4期
唐 庆,黄 欣,秦 波,李 冰,林 茂
(广西壮族自治区林业科学研究院,广西 南宁 530002)
【研究意义】干旱是影响植物生理生态特性及生长发育过程的重要因子,也是制约花卉产业发展的全球性问题。近年来,在全球变暖的气候背景下,极端天气所造成的气候灾害频发,干旱受灾区域逐渐由我国北部地区向南部地区蔓延。因此,如何提高植物对干旱环境的适应性一直以来都是花卉专家研究的热点之一。干旱是限制桂叶山牵牛推广应用的逆境因素,由于这些地区水资源匮乏,限制了以灌溉途径提高推广面积[1-2]。【前人研究进展】大量研究表明,干旱胁迫对植物体内水分的亏缺,导致植株矮小,叶片加速衰老和萎蔫等[3-4]。光合日变化特征是作物光合在一天中对特定环境作出的动态变化的响应[5-6],抗旱是通过维持较低的蒸腾速率来减少水分的缺失[7],适宜水分亏缺有利于提高水分利用效率[8]。在一定干旱胁迫下,叶绿体内部结构发生了不同程度的变化[9]。桂叶山牵牛植物属于速生藤本,适宜在边坡绿化、矿山修复等地种植,这不仅可以可以美化环境,提高绿地覆盖率,具有较好的景观效果;其次,边坡绿化可以涵养水源,在一定程度上预防和减少水土流失、滑坡等现象的发生[10]。但是边坡、矿山等地,地势险恶,土壤残留量少,严重缺水,直接影响植物的生长,因此研究桂叶山牵牛藤本植物具有生产指导实践意义。目前,桂叶山牵牛成熟苗对干旱胁迫的生理生化及叶绿体超微结构等这方面的研究尚未见报道,但有遮荫对大花山牵牛和桂叶老鸦嘴生长及叶片组织结构影响的相关报道。【本研究切入点】本研究选择桂叶山牵牛成熟苗为研究对象,采用盆栽控水法,以探讨干旱对桂叶山牵牛成熟苗的生理生化的影响。【拟解决的关键问题】通过比较桂叶山牵牛在不同干旱胁迫条件下的形态、光合参数和叶绿体超微结构,了解桂叶山牵牛对干旱胁迫的响应机制,为其抗旱栽培提供参考依据。
1 材料与方法
1.1 试验材料
试验地位于广西壮族自治区林业科学研究院园林花卉研究所(108°21′E,22°56′N),属南亚热带季风气候,年均气温20~21 ℃,≥10 ℃的年积温7 206 ℃,一般年份有2~4 d 轻霜,年均降水量1 350 mm,5—9 月为雨季、月降雨量在100 mm 以上,10 月至翌年4 月雨量较少、月降雨量在80 mm 以下[11]。
供试材料为桂叶山牵牛成熟苗,选择长势一致、具有8~10 对叶片,根系发达、无病虫害、株高190~200 cm 的优良植株。
1.2 试验方法
试验于2020 年7 月10 日至2020 年8 月10日在广西林科院园林花卉所基地进行,采用高透光遮雨棚下盆栽试验。胁迫前将试验苗充分浇透水肥,使土壤含水量及营养基本保持一致,待土壤自然干后,进行干旱胁迫处理。参照Hsiao[12]的方法,试验设正常供水(CK)、中度干旱(MD)和重度干旱(SD)3 个水分处理,即土壤含水量分别控制在田间持水量的70(± 5)%、50(± 5)%和35(± 5)%。于每天早上8:00、中午12:00、下午18:00 这3 个时间段用WET-2 土壤水分温度电导率速测仪测土壤含水量,水分不足时用有刻度的烧杯进行补水,并记录加水量。处理30 d 后,分别测定形态指标、光合指标并观察叶片叶绿体超微结构。
1.3 测定项目及方法
1.3.1 形态指标 胁迫前2020 年7 月10 日、胁迫后8 月10 日分别测定株高、地径、叶片数、叶长和叶宽、根系生物量(干鲜重)和地上部分生物量(干鲜重)等指标。
株高、叶长和叶宽:将试验植株水平放置,用卷尺测定株高(植株从基部至生长点)、叶长(从叶尖至叶柄)、叶宽(选取叶片最宽的位置测定)。
地径:用电子游标卡尺,轻轻卡住植株基部,测定地径值。
根冠比:将不同干旱胁迫处理的桂叶山牵牛成熟苗从盆中取出,用流水冲洗干净,将地上部分与地下部分分开,晾干并称重,用锡箔纸包好,放入烘箱,120 ℃烘48 h 至恒重,再分别称地上部和地下部分的干重。
根冠比=地下部重量/地上部重量
1.3.2 光合特性指标 2020 年7 月10 日至8 月10 日,在天气晴朗的条件下连续测定。测定时间为9:00~19:00,每隔1 h 测定1 次,每个处理测定3 盆,每盆测定3~5 片成熟叶,用Li-6400xt 便携式光合仪(Li-COR USA)测定叶片的净光合速率(Pn)、气孔导度(Gs)、细胞间CO2浓度(Ci)及蒸腾速率(Tr)等生理指标,同时记录气温(Tair)、叶温(Tleaf)及空气相对湿度(RH)等参数。
1.3.3 叶片细胞超微结构观察 2020 年8 月10日以测定光合特性的叶片为材料,按要求截取叶片样品大小为2 mm×2 mm,置于2.5%戊二醛中固定过夜,用0.1 mol/L 磷酸漂洗液漂洗3 次,每次15 min,将样品放入1%锇酸4 ℃冰箱中固定2 h;然后在0~4 ℃用50%、70%和90%乙醇逐级脱水,每次脱水时间15 min,再用100%乙醇和100%环氧丙烷分别脱水5 min,3 次重复;用环氧丙烷+包埋液(2∶1)包埋2 h,再用环氧丙烷+包埋液(1∶2)包埋3 h,纯包埋液浸泡过夜,最后用纯包埋液在室温浸泡3~4 h。将样品块置于包埋模板内,加入纯包埋剂后放恒温烘箱内聚合,37 ℃ 12 h,45 ℃12 h,60 ℃48 h,半薄(1 um)定位及超薄切片机切片(70 nm),3%醋酸铀-枸橼酸铅双染色,透射电镜观察,拍片。
试验数据运用Microsoft Office Excel 进行整理与作图,并进行均值和方差分析。不同干旱胁迫桂叶山牵牛形态指标、生理指标等采用DPS 7.05软件进行单因素方差分析(one-Way ANOVA)。
2 结果与分析
2.1 干旱胁迫对桂叶山牵牛生物量及气孔的影响
由表1 可知,桂叶山牵牛株高和叶片数随着不同干旱胁迫程度而变化,差异显著;MD 处理地径、叶长和叶宽与CK 差异不显著,而SD 处理与CK 差异显著;MD、SD 处理根系生物量(干重)、地上部分生物量(干重)、冠根比与CK差异显著,MD 与SD 处理间差异不显著,说明桂叶山牵牛成熟苗对不同干旱胁迫的生理调控机制,在形态上表现为桂叶山、牵牛成熟苗受到的干旱胁迫程度越大,生长受到的抑制就越严重。干旱胁迫对桂叶山牵牛成熟苗造成一定的影响,尤其是MD、SD 处理,地上部分株高、地径、叶片数和叶长宽显著降低,根冠比差异不显著,表明桂叶山牵牛成熟苗通过不同生长器官的结构资源分配比例保证自身的生长。桂叶山牵牛干旱胁迫前的生长量与胁迫后的生长量有显著差异,CK株高增长15.33 cm,而MD、SD 处理株高分别下降21.43、68.70 cm;地径在正常供水(CK)和中度干旱(MD)胁迫下分别增加0.94、0.51 mm,而重度干旱(SD)的地径下降0.30 mm,说明在重度干旱胁迫下桂叶山牵牛地径的生长受到一定的抑制;CK 桂叶山牵牛叶片数增加10 片,而MD、SD 处理的叶片数分别减少19 片和47 片,说明叶片是植物进行物质、能量及水气交换的主要器官,对环境变化敏感、可塑性强,能够影响植物的行为和功能。桂叶山牵牛干旱胁迫后的叶片大小均下降,这可能跟植物本身的生长有关,盆栽试验的基质以及营养不足,导致叶片的营养不足;干旱胁迫后根冠比增加,说明地下部分在增加,而地上部分减少。
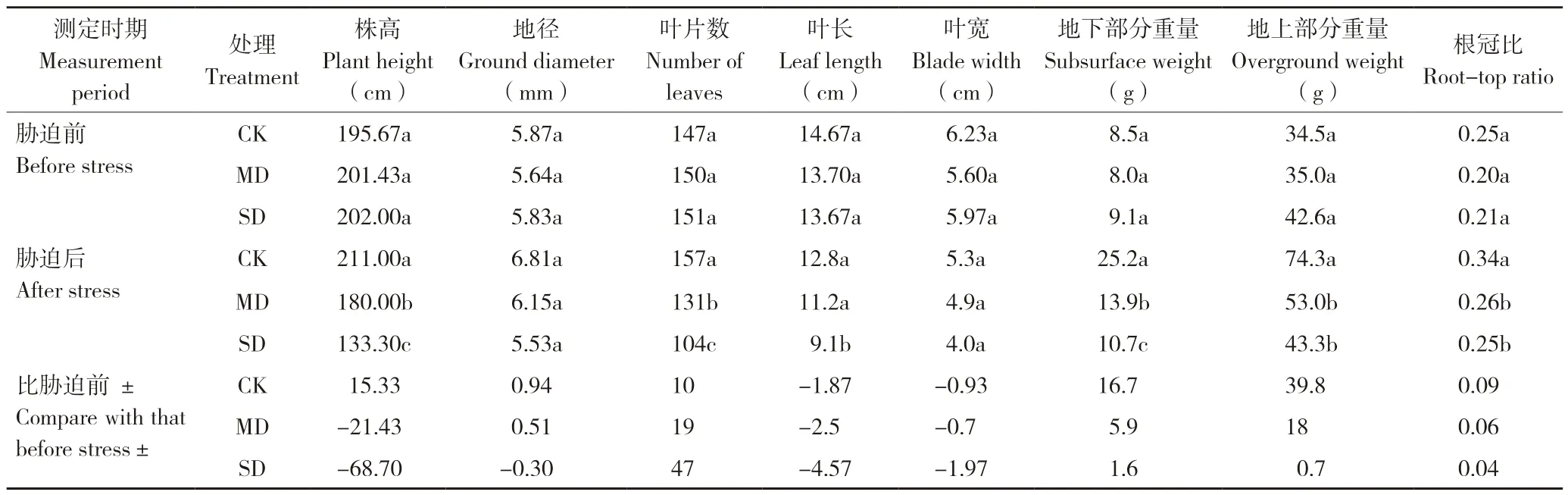
表1 桂叶山牵牛胁迫前后的相关生长量指标Table 1 Growth indicators of Thunbergia laurifolia before and after stress
2.2 干旱胁迫对桂叶山牵牛的气体交换参数日变化的影响
由表2 可知,随着干旱程度的增加,桂叶山牵牛叶片瞬时净光合速率、气孔导度、胞间CO2浓度和蒸腾速率显著降低,水分利用效率增加,气孔限制值无显著变化。其中,与对照相比,MD、SD 处理的净光合速率分别下降2.22、2.87 µmol/m2·s,蒸腾速率分别下降5.14、5.83 mmol/m2·s,水分利用效率分别增加0.41、0.29 µmol/mmol。
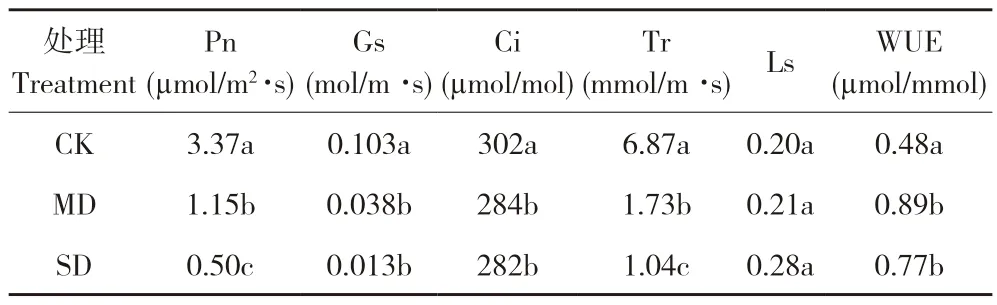
表2 干旱胁迫下叶片瞬时气体交换参数变化Table 2 Changes of instantaneous gas exchange parameters of leaves under drought stress
桂叶山牵牛净光合速率的日变化如图1A 所示,CK、MD 和SD 处理均存在光合“午休”现象,其中CK 的日变化为三峰曲线;MD 处理日变化为双峰曲线,第1、2 峰值出现在12:00 与14:00,以13:00 左右的净光合速率值最低;SD 处理的日变化不显著。综合分析,桂叶山牵牛净光合速率以CK 显著高于MD、SD 处理,依次为CK >MD>SD,说明其净光合速率随着水分的降低而降低。
桂叶山牵牛蒸腾速率的日变化如图1B 所示,CK 为三峰曲线,而MD、SD 处理的日变化均为双峰曲线,在9:00 的蒸腾速率较低,之后随着光合有效辐射的增加,气孔导度的扩张导致蒸腾速率增加,在13:00 和16:00 左右均出现双峰。综合分析,桂叶山牵牛的蒸腾速率以CK 显著高于MD、SD 处理,依次为CK >MD >SD。
由图1C 可知,桂叶山牵牛胞间CO2浓度日变化受到大气中CO2浓度和气孔导度的影响,CK和MD 处理的胞间CO2浓度在13:00 最低,而SD处理胞间CO2浓度在13:00 增加;CK 在9:00 和19:00 显著高于其他时间段,而在18:00 下降,这可能是受到外界因素影响所致。
图1D 显示,不同干旱胁迫下桂叶山牵牛气孔导度依次为CK >MD >SD,其中CK 的桂叶山牵牛气孔导度日变化为三峰曲线,峰值出现在10:00、12:00 和14:00 左右;MD 处理出现明显的双峰;而SD 处理的气孔导度变化不大,与净光合速率呈正相关关系。

图1 不同干旱胁迫桂叶山牵牛气体交换参数日变化Fig.1 Diurnal variationof gas exchange parameters of Thunbergia laurifolia under different drought stress
2.3 桂叶山牵牛气孔限制值和水分利用率的日变化
不同干旱胁迫对桂叶山牵牛气孔限制值的日变化见图2A,CK 桂叶山牵牛气孔限制值在13:00呈单峰,而MD、SD 处理桂叶山牵牛气孔限制值均在13:00 最低。水分利用效率的日变化(图2B)均呈双峰曲线,随着净光合速率的增加,桂叶山牵牛通过调节气孔关闭的方式减少水分散失,在13:00 左右均出现光合“午休”现象,此时水分利用效率的值最低,其中CK 的水分利用效率显著高于MD、SD 处理,依次为CK >MD>SD。

图2 不同干旱胁迫桂叶山牵牛气孔限制值和水分利用效率日变化Fig.2 Diurnal variations of stomatal limits and water use efficiency of Thunbergia laurifolia under different drought stress
2.4 干旱胁迫对桂叶山牵牛叶片叶绿体超微结构的影响
由图3A、D 可知,不同程度干旱胁迫下,桂叶山牵牛的叶绿体结构均发生较明显的变化。其中,CK 的桂叶山牵牛叶片类囊体片层排列整齐,层数多,淀粉粒多、饱满、体积大,嗜锇颗粒少、体积小;MD 处理的类囊体片层排列整齐,层数多,淀粉粒少、不饱满、体积小,嗜锇颗增多、体积增大;SD 处理的类囊体片层排列不整齐,层数多但不紧密,淀粉粒少、干瘪、体积小,嗜锇颗粒多、体积大。说明MD、SD 处理干旱胁迫严重影响叶绿体结构类囊体结构的排列,淀粉粒的减少、干瘪以及内含物增加。
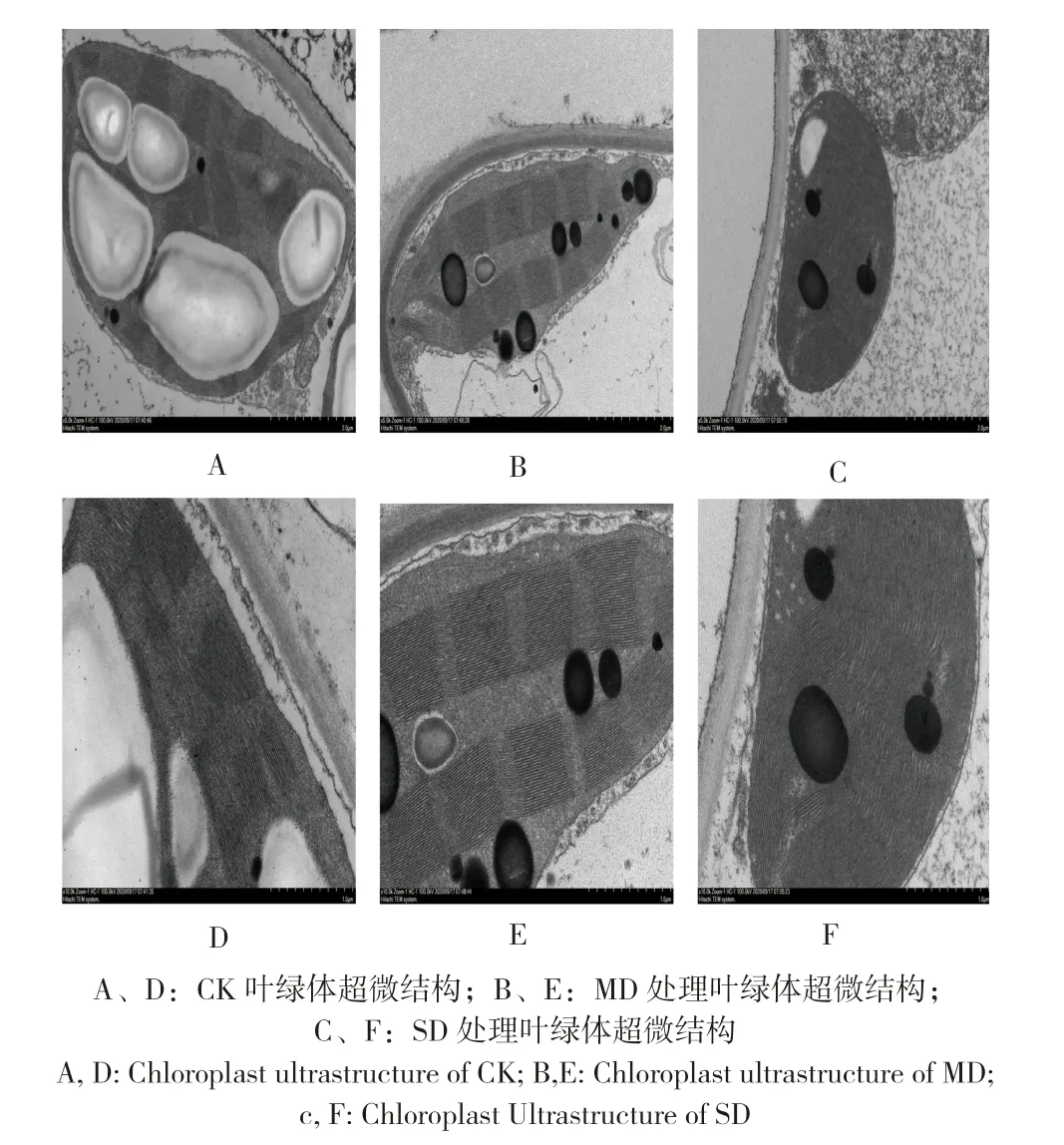
图3 不同干旱胁迫对桂叶山牵牛叶片叶绿体超微结构的影响Fig.3 Effects of different drought stress on chloroplast ultrastructure in leaves of Thunbergia laurifolia
3 讨论
3.1 干旱对桂叶山牵牛形态特征的影响
李颖慧等[13]认为随着干旱胁迫程度的加重,不同籽用西瓜品种间叶片出现萎蔫、变黄、褐化的程度不同;范苏鲁等[14]认为植物受到干旱胁迫时最直观的反应就是外部形态上的变化,干旱通常会导致植物水分亏缺,延缓或破坏植物的正常生长。本试验中,干旱胁迫对桂叶山牵牛成熟苗的生长造成一定的影响,MD、SD 处理地上部分株高、地径、叶片数和叶长宽显著降低,根冠比差异不显著,表明桂叶山牵牛成熟苗通过不同生长器官的结构资源分配比例保证自身的生长。因此,桂叶山牵牛成熟苗对不同干旱胁迫的生理调控机制,表现在形态上便是桂叶山牵牛成熟苗受到的耐旱程度越大,生长受到的抑制就越严重。
3.2 干旱胁迫对桂叶山牵牛光合指标的影响
萧自位等[15]认为土壤水分含量的变化导致植株叶片光合的变化;崔莹莹[16]认为不同植物叶片的净光合速率对光质的响应机理并不完全相同;张志焕等[17]认为轻度干旱胁迫时,光合作用下降的原因是气孔限制,而在缓慢干旱、中度干旱和重度干旱时,光合作用下降是非气孔限制。本研究中,以正常供水(CK)为对照,随着水分的减少,桂叶山牵牛净光合速率和蒸腾速率的日变化均存在下降趋势,MD、SD 处理的净光合速率分别下降2.22、2.87 µmol/m2·s,蒸腾速率分别下降5.14、5.83 mmol/m2·s,说明中度干旱和重度干旱时光合作用降低的原因由非气孔限制所导致,同时也这表明当水分严重亏缺时,桂叶山牵牛光合机构受严重损害以致电子传递速率降低,与段娜等的[18]研究结果一致,即桂叶山牵牛成熟苗通过降低光合作用来提高抗旱能力。
3.3 干旱胁迫对桂叶山牵牛水分利用的影响
邓沛怡等[19]认为水分利用效率(WUE)是植物抗旱性的重要指标,干旱胁迫有利于提高植物的水分利用效率;何平如等[20]认为土壤水分下限过高,棉花单株有效铃数降低,产量增加不再明显,水分利用效率较低;郭贤仕等[21]、刘凯强等[22]认为干旱能引起植物的水分利用效率增加并不是耗水绝对量的减少,而是干旱锻炼增强了叶片的同化功能,其同化能力超过蒸腾速率的增加。本研究中,MD、SD 处理的水分利用率较高,分别为0.89、0.77 µmol/mmol,说明MD处理桂叶山牵牛水分利用效率较高,仅限于在一定的范围内能恢复植物受损的细胞,否则植物因光系统的损伤会导致植物净光合速率处于较低水平,这与黄刚等的[23]研究结果一致,即桂叶山牵牛成熟苗通过提高水分利用效率来抵御抗旱。
4 结论
桂叶山牵牛成熟苗会通过降低光合作用、关闭气孔及提高水分利用效率等方式来应对环境恶劣的干旱条件。桂叶山牵牛在正常供水(CK)条件下长势较好,叶绿体结构排列整齐,中度干旱(MD)胁迫下叶绿体结构发生了变化,而重度干旱(SD)胁迫下虽然叶绿体结构严重受损,但是成熟苗未见有死亡,说明干旱胁迫处理桂叶山牵牛成熟苗的耐旱性较强,能够在园林中应用。
猜你喜欢
农业工程学报(2022年4期)2022-04-24
小学生优秀作文·时尚版·中年级(2021年3期)2021-09-10
节水灌溉(2021年5期)2021-05-26
小学生优秀作文(中年级)(2021年3期)2021-03-16
农业与技术(2020年12期)2020-07-04
新课程·下旬(2017年7期)2017-08-14
江苏农业科学(2015年8期)2015-09-10
天津农业科学(2015年8期)2015-08-06
语文世界(小学版)(2015年5期)2015-07-06
湖北农业科学(2014年13期)2014-08-28
