
荔枝成花过程及其重要制约因子
2021-05-30 13:26梁立峰
广东农业科学 2021年4期
梁立峰
荔枝(Litchi chinensisSonn.)为亚热带常绿果树,原产我国南方,目前在两广交界地区、海南及云南仍有野生荔枝分布[1-3]。越南也是荔枝起源地之一[3]。我国荔枝种质资源非常丰富,是世界上最重要的荔枝产区。世界各地的荔枝基本是直接或间接从我国引入[1-3]。荔枝果实色、香、味俱佳,营养丰富,是著名的岭南佳果。然而,荔枝也以其大小年结实之严重著称,特别是晚熟品种和一些中熟品种。历史上,一年丰收,要两三年才能恢复正常收成是常有之事,这是一种实质性的“不规则结实”。荔枝的大小年结实现象向来受到栽培者及研究者重视。一般认为,大小年结实与树体内的营养状况及生理状态有着密切关系,但环境条件对荔枝开花结实过程的制约太过强烈,或者说荔枝开花结实对环境条件的要求太过苛刻,也是重要原因之一。有花不一定有果,无花肯定无果,因此人们一直重视对荔枝成花过程及促进荔枝成花的研究。研究者们从物候学、形态学等角度对荔枝花序、花器官的发育及成花进程作了很多研究。他们的众多不同观点常让读者在概念上无所适从。本文旨在梳理前人有关荔枝花序结构及成花过程的研究成果,并在此基础上提出见解,然后讨论影响荔枝成花的重要因子,特别是树体营养状况的改变及环境温度变化对荔枝成花过程的制约作用,举例说明应对不良环境因素制约的主要手段。
1 荔枝的成花过程
1.1 荔枝花序的结构
荔枝花序发生于上年的秋梢结果母枝的顶芽或近顶部叶片的腋芽,也有从上年落花落果枝或结过果的枝条的近顶部叶腋间抽出[1-5]。田间观察可知,荔枝花序也可以直接从主枝或主干的潜伏芽抽生出来。
对荔枝所抽生的花序有多种不同表述,一般称其为圆锥花序[6-9],也有人称为复总状圆锥花序[3]或复圆锥花序[10]。多位学者认识到荔枝花序里的聚伞花序成分,Pandey 等[11]指出荔枝花序在经过多级分枝后聚伞状着花,有人认为荔枝花序是由许多小聚伞花序形成的圆锥花序[1],或是大型的复合或单独的聚伞状圆锥花序[2,12]。吴仁山[4]认为,荔枝花序属聚伞花序,呈圆锥状排列,组成一个聚伞圆锥花序。Robbertse 等[13]对荔枝花序结构仔细观察后提议当“Thyrsoid”(英文词典将其译为“聚伞圆锥花序”)看待。查阅英文植物术语文献可知,Thyrsoid(Thyrse)是指花序主轴无限生长、侧生花序轴有限生长、形似圆锥或圆柱的花序,但荔枝花序与此有一定区别,其侧生花序轴也是总状花序,会无限生长。
许多学者报道过荔枝花序轴的多级分化过程。倪耀源等[2]从物候学及形态学角度对荔枝花序的发育作长期观察后指出,花序原基伸长成为圆锥花序主轴,并由下而上分化出雏形复叶,叶腋内产生肥大的第一级枝梗(侧轴)原基,肉眼渐见嫩梢叶腋出现越来越明显的“白点”,一般称为抽穗;第一级枝梗伸长并产生数片苞片,各苞片腋间产生第二级枝梗(支轴)原基;第二级枝梗再产生苞片及第三级枝梗(小穗轴)。童宗伦等[12]也观察到荔枝有三级侧花序轴的分化,分别定义为第一级支序、第二级支序及小花序。张君圻[10]则将荔枝三级侧花序轴分别定义为侧序、支序及小序。吴素芬等[9]发现荔枝花序轴有三级分枝,定义第三级分枝为小穗轴。Robbertse等[13]也报道荔枝有三级侧花序的分化。花序的多级分枝为荔枝花序构建了一个良好的骨架。
根据植物学的花序定义及各种花序的发育特点可知,上述荔枝花序轴的多级分化过程循圆锥花序(复总状花序)的分化方式。从圆锥花序的主轴抽生出侧轴,越往上分枝级数越少,最终均于顶端出现小穗轴,其中花序下中部的侧生花序轴先分化出小穗轴。以上提及的小穗轴、小花序及小序虽然名称不同,但实质上是同一级别的小花序轴,均为荔枝花序中圆锥花序部分的末次分枝,本文统称为“小穗轴”。小穗轴的出现在荔枝成花过程中具有重要意义,因为此刻荔枝花序将转入聚伞花序的分化。
小穗轴出现后的分化方向一直备受关注[2,9,10,13-14]。Robbertse 等[13]认为小穗轴的进一步分化循二岐聚伞花序(Dichasium,也称歧伞花序)的分化方式进行,即先形成顶花,顶花形成后在其下方两侧同时抽出一分枝,分枝顶端又生顶花,此顶花下方再以同样的方式从两侧各产生一分枝,荔枝二岐聚伞花序中的小花分布方式及开花次序如图1 所示。因分枝级数不同,分化的结果生成由3 朵花、7 朵花或15 朵花组成的大小不同的花穗。
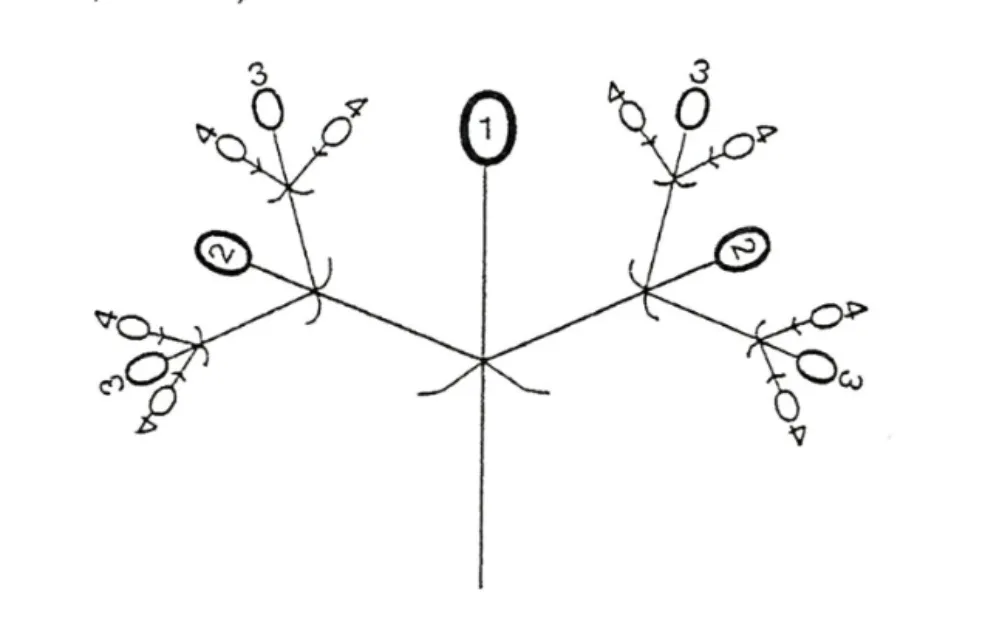
图1 荔枝二岐聚伞花序上的小花分布方式(侧面观)及开放次序[13]Fig.1 Distribution mode of flowers on lychee dichasia in lateral view and bloom order[13]
综上可知,荔枝花序是圆锥花序(复总状花序)和聚伞花序的组合体。两种花序在功能上各司其职,圆锥花序只负责基础性的花序骨架构建,故被认为是“骨架花序”;二岐聚伞花序负责花和花器官的发育及为小果提供发育居所,可以称为“开花花序”。因此,荔枝花序实际上由“骨架花序”和“开花花序”共同构成,是两种花序的分工合作成就了荔枝的开花结实。由于荔枝最终以聚伞花序结果,聚伞花序是花序主角,圆锥花序只是花序基础,笔者认为应将荔枝花序称为“圆锥基聚伞花序”。
1.2 荔枝成花的步骤
一般将果树的花芽分化分为生理分化和形态分化两个阶段。李嘉瑞等[15]指出,从花原基最初形成至各花器官形成完成叫形态分化。此前生长点内进行的由营养生长向生殖生长转变的生理生化变化叫生理分化。引起生理分化的因素叫诱导。
关于荔枝花芽分化进程的研究很多,国内学者将荔枝成花过程划分为几个阶段或时期,国外学者较少作类似划分。倪耀源等[2]将荔枝花芽分化划分为3 个时期:花序原基形成期、花序各级枝梗分化期及花器分化期;吴仁山[4]认为荔枝花芽分化经过生理分化、形态分化和性细胞形成3个阶段;童宗伦等[12]将荔枝花芽分化分为花序原基分化(及花序形成)与花芽分化(及花各部分形成)2 个时期;季作樑等[8]将荔枝花芽分化过程分为圆锥花序分化、花器官分化两个部分,每个部分再细分几个时期;张君圻[10]将荔枝花芽分化分成6 个时期:花序原基分化始期、主序分化期、多级侧序分化期、花萼分化期、雄蕊分化期及雌蕊分化期;吴素芬等[9]将荔枝花芽分化划分为3 个时期:未分化期、花序原基分化期、花序分枝及花器形成期;Menzel[16]将荔枝的生殖过程划分为圆锥花序分化及抽生、圆锥花序发育、开花、果实发育等4 个阶段;黄辉白等[17]以图解方式表述荔枝成花过程的顺序性和阶段性,并将花芽分化进程分成前诱导期、成花诱导期和花发端发育期等3 个阶段。
由于荔枝成花过程需经历两个重要转折点,一是末次秋梢顶芽从生理分化状态转为形态分化,二是发育中的花序从圆锥花序分化状态转入聚伞花序的分化。本文以这两个转折点为基准,认为荔枝成花过程分3 个步骤完成。之所以不以“时期”或“阶段”对荔枝成花过程进行分段,是因为“时期”或“阶段”之间的界限比较明确而且接续性明显,而荔枝在整个花序发育过程中有一段时间是圆锥与聚伞两种花序分化同时进行的[2,8-12,14]。荔枝成花过程的3 个步骤包括花序原基的诱导和孕育、圆锥花序抽生及花序骨架构建、聚伞花序分化及花器官建成。
1.2.1 花序原基的诱导和孕育 在亚热带地区,荔枝的末次秋梢自深秋起因降温或雨水减少出现生理胁迫而进入一种相对休眠状态,常称被迫休眠[18]。此时营养生长停止,体内养分逐渐积累,枝梢逐渐老熟。树体因光照、温度、水分条件发生变化,体外体内营养条件发生变化,体内代谢途径随之改变,朝着有利于孕育花序原基的方向进行。处于被迫休眠状态的荔枝受多条内外信号途径诱导,形成一个复杂的调控网络,激活茎端分生组织特性基因表达[19]。随后营养分生组织转换成生殖分生组织,生长锥逐步突起呈平滑穹顶状,多位学者认为此时花序原基已经初步形成[2,8,10,12]或处于花芽形态分化前奏[6-7,11]。一般认为,上述过程属于成花诱导,其实质是荔枝秋梢在“被诱导”的情况下主动进行一系列重大生理生化变化。
尚未有人研究过枝干上的潜伏芽如何转化成花芽,可以理解为环境因子及体内条件的改变诱导潜伏芽顶端分生组织发生一系列生理生化变化,最终导致花序原基形成。
改变一系列代谢途径以适应孕育花序原基的需要,是荔枝成花过程首先发生的事件,孕育花序原基是荔枝成花过程第1 个步骤的任务。
1.2.2 圆锥花序的抽生及花序骨架的构建 末次秋梢茎端的花序原基在适宜的环境条件配合下,循圆锥花序(复总状花序)的分化方式向上伸长成为花序主轴,主花序轴从基部开始、自下而上从节点苞片腋长出第一级侧花序轴。从花序基部或近基部节点长出的第一级侧花序轴较大,越往上越小,直到最后不再有分枝。在主花序轴不断发育的同时,花序基部及近基部的第一级侧花序轴产生第二级侧花序轴,接下来是第三级侧花序轴(即小穗轴)。小穗轴自下而上先后出现,一个具有多级分枝的荔枝花序骨架逐步构建完成。
抽生圆锥花序(或称“抽穗”)及构建花序骨架是荔枝成花过程第2 个步骤的任务,为下一步着花、开花奠定基础。
1.2.3 聚伞花序的分化及花器官的建成 荔枝花序骨架搭建好后,花序末次分枝(即小穗轴)转入花穗分化,循二岐聚伞花序(岐伞花序)的分化方式。该花序发育方向的转变可能是荔枝在系统发育过程获得的固有发育程序,其中有花序分生组织特性基因的参与[20]。荔枝花穗呈聚伞状着花,3 朵小花组成一个小穗,2 个小穗组成一个7 朵花的中花穗,4 个小穗组成一个15 朵花的大花穗,某些品种还可见31 朵花的特大花穗出现[2]。荔枝小花的花器分化由外到内依次分化萼片、雄蕊、雌蕊,多数不见花瓣分化。
“小穗轴”转入聚伞花序分化及建成各花器官是荔枝成花过程第3 个步骤的任务。
2 影响荔枝成花的重要内外因素
如上所述,荔枝成花过程非常复杂,分3 个步骤进行。从进入生理分化算起,经历花序原基孕育、抽穗及构建花序骨架,到最后开花坐果,需要经历4~5 个月。期间常因不良内外因素的影响而最终不能成花。一直以来,制约荔枝成花及影响荔枝收成的内外因子备受关注,多位学者对此已作过包括生化及分子水平调控机理在内的多方面综合述评[5,7,16,21-24]。本文着重讨论树体营养状况及寒潮入侵带来的温度变化对荔枝成花的制约作用,也涉及水分及日照条件对荔枝成花的影响。
2.1 从增城49 年荔枝产量变化看荔枝大小年结实的规律及内部影响因子
廖美敬[25]研究了1949—2017 年广东增城荔枝总面积及总产量逐年变化情况(图2)。由于增城一直以优质但不稳产的桂味、糯米糍为主栽品种,所以图2 反映的荔枝大小年结实现象很有代表性。该图显示了增城荔枝产量变化的基本趋势:大年结实后的第2 年甚至多年均为小年,而大年结实的前一年或前2~3 年均为小年。其中,1949 年至20 世纪80 年代中期的产量变化最具可比性和参考性,因为期间荔枝栽种面积基本稳定,栽培管理水平差别不大,当时还未大量使用化学或物理调控技术。

图2 1949—2017 年广东增城荔枝栽培面积和产量变化[25]Fig.2 Changes of annual planting area and yield of litchi in Zengcheng,Guangdong province,1949-2017[25]
由于荔枝每次大年结实后都是小年,很难认为小年都因环境不适引起。一般认为荔枝大年的过量挂果耗竭树体内养分是次年不能再次开花结实的主要内部原因,因为荔枝是一种可溶性固形物含量极高的水果,其成长过程需要消耗大量碳素和矿质营养。对大年结果的前一年或前2~3 年均为小年的现象,可以理解为荔枝树体需要积蓄足够养分,才能在合适气候条件配合下成就一次大年结实。由此认为,荔枝获得良好收成的前提条件,首先是具备充足的树体营养,其次需要有适当的环境条件,包括良好耕作技术的配合。
倪耀源等[26]认为,丰产年份的荔枝过度消耗树体有机营养,使根群生长受影响,吸收能力减弱,秋梢结果母枝不能依时萌发,是导致次年少有或没有花果的原因。Hieke 等[27]研究也支持上述观点,他们发现荔枝新叶的伸张和转绿需要根部输送同化物质。Malhotra 等[23]也指出,荔枝过度挂果会耗尽树体内碳水化合物等营养物质,植株无法提供足够营养用于采后秋梢抽发及老熟。据此,可用荔枝采果后能否抽生健壮秋梢来估量树体采后“体能”及来年成花能力。
荔枝采后抽生秋梢的老熟程度及营养状况对来年成花结实有重要影响。肖秋生[28]发现,自然条件下,荔枝成花率与末次秋梢的成熟度呈正相关,通过对不同成熟度的末次梢作相应处理并促进其老熟,有利于荔枝成花。
“大年过后必小年”现象在海南、粤西、桂南等以早熟和中早熟荔枝为主栽品种的地区表现并不明显,因为这些品种的成花对低温需求不严格,成花相对容易,而且这些品种采收期早,年中有充裕时间让树体恢复,总体表现稳产一些,但若管理不善或天气条件不佳也会出现明显的大小年结果现象。
2.2 温度条件对荔枝成花的影响
在诸多影响荔枝成花的环境因子中,温度因素最受重视,多数研究都是探讨具体的温度指标对荔枝花芽分化及来年产量的影响。由于冬春的温度变化均因寒潮侵袭而发生,即在寒潮入侵与具体温度指标的变动之间存在一种“因果”或“纲目”关系,所以本文以寒潮侵袭为切入口探讨温度因素对荔枝成花的制约作用。
2.2.1 冬春温度变化控制荔枝结果母枝和花序轴上的营养生长与生殖生长之争 荔枝被认为是一种对气候变化非常敏感的作物[29],同时又是一种顶端优势较强、偏好营养生长的果树,只要温度合适,深秋初冬仍可以萌发新梢。荔枝抽穗后,花序主轴及侧轴节点的苞片腋芽会因低温胁迫程度的不同而向不同方向发育。若低温胁迫条件合适,营养生长停止,节点上的苞片脱落,苞片腋芽顺利进入花芽分化。强冷天气还能让刚抽生的冬梢转长成带“白点”的短“花棍”。若低温胁迫强度不够或胁迫条件解除,苞片长成小叶而腋芽长出枝梢,或出现花序与小叶共存的现象,或花序主轴下部有侧花序但中部出现梢叶,即“花变”现象。这并不是真正意义上的“变”,而是营养生长与生殖生长的相互取代,这种取代主要因冬春温度的不断变化而引起。
2.2.2 寒潮入侵导致冬春温度变化进而制约荔枝成花过程 华南冬春经常有寒潮入侵。伍红雨等[30]观察到,广东、广西、海南3 省192 个气象站在1961—2008 年间共有221 次寒潮出现,平均每年4.6 次,出现时间以12 月至3 月为主,1 月是寒潮发生最频繁的月份,其次是2 月。林爱兰等[31]对广东均匀分布的11 个气象站1951—1994 年间的寒潮情况进行统计,发现43 年共出现103 次寒潮,平均每年2.3 次,出现时间以12月至2 月为主,1 月频数最高、强度最大,研究还显示广东寒潮的出现存在年际变化,一般冬春出现1~5 次,最多出现6 次,有5 个冬季无寒潮。曾踪等[32]发现,广东冬春历次寒潮或强冷空气造成的低温日数、降温幅度和最低温度年际间差别很大,从未造成低温日(日均温≤12℃)到最多连续13 d 出现低温日不等。综上可知,华南冬春寒潮入侵次数、强度、造成的低温日数及最低温度在年际间差别很大。
若冬春没有冷空气入侵,荔枝无法进入被迫休眠状态,结果母枝及花序轴上的营养生长与生殖生长之争就无法解决。不仅如此,梁立峰[18]还指出,广东、广西地区如果冬季前后两次寒潮间隔时间太长,其间出现温度较高时段,荔枝将萌发大量冬梢,导致花芽分化无法进行。可见,冬春寒潮如何入侵是制约荔枝成花的最重要环境因素。如果冬春寒潮出现少、强度小或间隔时间过长导致冬春温度偏高,无法抑止营养生长,未能让生殖生长占据优势,荔枝将很难良好成花。
2.2.3 从玉林两个冬春的寒潮活动看荔枝大丰收与寒潮降温过程的相关性 2002 年广西玉林市荔枝大丰收,为历史最高产;此前的1999 年也获得丰收,为当时历史最高产。陈国保[33]研究两个丰产年的气象成因发现,2001—2002 年冬春有3次降温过程,分别于11 月中旬、12 月中旬初及1 月中旬末出现,其中第2 次降温持续1 个月,日均温≤12℃总天数66 d、≤10℃总天数40 d,连续≤10℃的最长天数在3 次降温过程中依次为5、6、7 d,极端最低温为3.4℃。1998—1999 年冬春有4 次降温过程,分别于12 月初、1 月初、2 月初及2 月下旬初出现,日均温≤12℃总天数41d、≤10℃总天数30 d,连续≤10℃的最长天数在4 次降温过程中依次为3、8、3、3 d,极端最低温为5.6℃。根据推算,2001—2002 年冬春3 次降温间隔时间依次是12、7 d,1998—1999 年冬春4 次降温间隔时间依次是16、12、13 d。
以上数据显示,2002 年荔枝比1999 年更丰产的原因有几个:(1)2001—2002 年冬春日均温≤12℃及≤10℃的总天数多于1998—1999 年时间;(2)2001—2002 年冬春降温时间比后者提前18 d;(3)2001—2002 年冬春两次寒潮间隔时间较短,最长为12 d;(4)2001—2002 年冬春各次降温过程都有连续5 d 以上的≤10℃总天数,且极端最低温较低。
2.2.4 华南地区荔枝成花过程需要冬春寒潮适时适度入侵 前人对10℃在荔枝成花过程的重要意义非常关注[2,21,33-35]。综合前人研究成果初步认为,有利于荔枝成花的冬春温度条件是:冬春寒潮入侵4~5 次,早寒潮能于11 月中下旬出现,两次寒潮间隔期在12 d 以内,寒潮日均温≤10℃的连续最长天数达6 d、日均温≤10℃的总天数达40 d,极端最低温达3℃~4℃。以上温度条件主要针对晚熟、中熟品种而言,一般很难完全满足,但具备条件越多对荔枝成花越有利,该结论有待进一步证实。由于华南地区的寒潮频发时间与广东增城、广西玉林一带的晚、中熟荔枝花芽形态分化关键期大致相近,可以认为岭南能成为世界上最著名的优质荔枝产地,是因为华南地区的冬春季节常有寒潮多次适时适度入侵。
自然条件下,华南地区各地方年际间寒潮的出现次数、出现时间、间隔期以及降温幅度、低温持续时间等差别很大,使得不同年份冬春季节的天气状况千差万别。即使树体营养条件已经达到成花要求,但如果天气不配合(如暖冬频频出现),荔枝也很难成花。除温度条件外,秋冬降雨量也影响荔枝成花,但基本可认为冬春寒潮的不适当侵袭使荔枝无法抑制营养生长进而导致成花不能顺利进行,是荔枝不规则结实的重要原因。
2.3 水分条件对荔枝成花的影响
秋季土壤湿度高会促进营养生长而抑制成花[7,36],但未有结论性证据说明水分胁迫直接参与诱导荔枝的成花过程[7,37],水分在荔枝成花中的作用被认为是间接性或辅助性的[38-40]。
傅汝强[41]研究荔枝收成与花芽分化期间降雨量关系时发现,在18 个考查年份中,凡大年结实年份,2 月份降雨量均在29 mm 以上。尹金华等[36]也发现,能获得荔枝大年收成的年份,其2月上旬、中旬降雨量高于小年收成年份。可见,荔枝在花芽分化后期需要适当水分。笔者有一年在田间观察到的现象支持上述观点,当时久旱不雨导致荔枝雌花无法开放,经历傍晚一场大透雨后,第2 天早上所有雌花均盛开。
尹金华等[36]研究发现,在12 月下旬到次年1 月下旬花芽分化关键期,小年收成年份的总降雨量显著高于大年收成年份。即这段时间降雨太多对荔枝成花有不良影响,这可能与冷雨水大量渗入使根系分布的土层降温太多,影响根系的正常生长与吸收,进而影响花序发育有密切关系。
2.4 日照条件对荔枝成花的影响
2.4.1 日照时数与荔枝丰歉的关系 对日照条件与荔枝成花的关系研究相对较少,多篇综述并未涉及日照条件对荔枝花芽分化的影响[5,7,14,23]。荔枝原是热带低山雨林中的优势树种,具有喜光特性[1],光照条件对荔枝成花结实有重要影响。蔡世同等[39]分析广东高州日照条件与荔枝丰歉的关系时发现,上年12 月至次年2 月的日照总时数平均值丰产年份为402.8 h、歉收年份为288.6 h。傅汝强[41]也发现,在广西博白地区,12—1 月的日照时数≥246 h 的年份多为大年,12—1 月阴雨总日数在<27 d 的年份均为大年,12—1 月阴雨总日数≤30 d 的年份基本为小年。但也有研究者认为冬季日照时数不是影响荔枝花芽分化的主要因子或关键因素[42-43]。日照条件对荔枝花芽分化的影响有待进一步研究。
2.4.2 荔枝在昼长变短的深秋进入花芽分化值得探究 一般认为,多数木本果树的花芽形成对光周期无反应[44]。Nakata 等[45]以“Kwai Mi”荔枝(实为“Mauritius”[14],即“大造”)的4 年生圈枝盆栽树为试材,研究认为荔枝属于日中性植物,此后人们一直认同这个观点[7,14,21-22]。但无论在我国华南地区还是其他国家,荔枝接受成花诱导并进入形态分化均于秋天发生。海南澄迈县的三月红荔枝于11 月上中旬进入生理分化,11月下旬花芽萌动并开始抽穗[46],澄迈县11 月份的日平均气温是24℃,日平均最高气温为28℃、日平均最低气温为21℃。澄迈及海南荔枝为何能在如此温度条件下成花一直是未解之谜。澄迈县每年从11 月起雨水减少直至翌年4 月,曾怀疑海南荔枝成花可能与当地旱季有关,但陈珍莉等[42]及吴志祥等[47]并不认为海南的降雨量与荔枝成花有相关性。澄迈县三月红荔枝在11 月上中旬进入生理分化阶段,澄迈县此时的昼长约11 h 15 min;11 月下旬花芽萌动并开始抽穗,此时的昼长约11 h 3 min。光被认为是能源及发育信号被植物利用[29],昼长变短能让落叶果树的叶片转黄及脱落,昼长变短与三月红荔枝成花是否相关值得探究。周碧燕等[48]发现三月红荔枝能在较高温度条件下进入花芽分化,提出并非所有荔枝品种均为日中性植物的疑问。
3 调节树体生理状态是荔枝促花保花的主要手段
自然环境的变化是不可控因子。荔枝速生快长,可对自然环境变化作出快速响应,这是可改变因子。在不良环境条件下,调整荔枝树体的内部生理状态是主要促花保花手段,通过调节荔枝各器官间的养分分配,平衡营养生长与生殖生长,对荔枝的成花结实有重要影响。
20 世纪80 年代兴起的荔枝控梢促花技术,正是以深入研究荔枝花芽分化及其调控机理为技术依托。控梢促花技术分化学措施、物理措施两类,均以抑制营养生长、防止因暖冬春引起的冬梢或花序小叶的发生为目标,这些技术包括花序抽生前后施用控梢促花素及适时环割、环剥等[49-50]。广东东莞荔枝专业户叶钦海选用优质但不稳产的中早熟品种妃子笑作为主攻品种,创立“一抹二疏三短截”和“一控一短截”技术分别应用于幼年结果树及成年结果树,达到调整花期、减少花量、提高花粉发芽率、增加雌花比例、延长雌雄花相遇期等效果,提高了荔枝座果率[51]。坐果后再作环割处理,抑制根系生长,使养分集中支持果实发育,从而提高了产量[51]。该研究成果使妃子笑成为一个能够高产稳产的荔枝品种,在广东、广西、海南乃至全国得到迅速发展。
荔枝雌花开过后常因雄花的再度开放而大量脱落,雄花数量因树体营养状况及天气条件而异。笔者于20 世纪90 年代初研发了“花开即保”技术[52],该技术花与果兼保,目的是保住刚完成开放及授粉过程的雌花,技术要点是选择在雌花刚开放、柱头转褐色时喷施生长调节剂和营养素合剂“坐果露”(也称“花果露”),以促进受精子房迅速发育并成为耗用型代谢库,以减少下批雄花开放量、降低雌花落花率、提高座果率[52-53]。该技术也称雌花期保果技术,由于其保花保果功效稳定、使用安全,时至今日仍为广大果农所使用。
荔枝树挂果过量会引发体内营养分配失衡。为防止因树体养分过度消耗进而影响根系生长及秋梢抽发,需要及时早期疏果及适时提前采果,这是一种提前促花措施。果树学要求叶片数量与果实产出量应有一个适当比例。
4 展望
我国荔枝栽培技术自20 世纪80 年代起获得迅猛发展,研究和实践成果丰硕。近年来,荔枝产量和果品质量不断提高,但离真正解决荔枝大小年结实问题还有一定距离。对荔枝大小年结果影响最大的内部因素是大年过度挂果,因此研究荔枝的适度挂果、提出合理的“叶果比”及配套的栽培技术很有必要。在外部因子中,由寒潮入侵导致的温度变化对荔枝成花起着重要制约作用。深入研究史上荔枝丰收年与相应天气状况的关系有助于认识环境因子如何制约荔枝成花,为解决生产问题提供理论依据。
荔枝品种间梢期、抽蕾期、花期等生长及成花习性不同,均随天气状况而发生变化,相应的栽培技术也应因品种及气候条件不同而有所区别,对各优良品种加以细致的物候及生长发育特性观察,获取准确的田间观察或田间试验数据,可促进新栽培技术的创立,也可对品种区划及对特殊小气候环境的利用具有重要指导意义。
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23
扬子江诗刊(2022年1期)2022-01-10
扬子江(2022年1期)2022-01-07
金山(2021年10期)2021-11-02
中国食用菌(2021年3期)2021-05-08
园艺与种苗(2021年1期)2021-03-16
青年文学家(2020年22期)2020-08-31
果农之友(2020年5期)2020-06-21
农业与技术(2019年5期)2019-05-29
农民致富之友(2018年21期)2018-12-27
