
干旱及复水对‘红富士’苹果生长及果实品质和产量的影响
2021-05-28 02:48高传彩惠基运魏玉兰张蕊刘建廷肖伟李玲
山东农业大学学报(自然科学版) 2021年2期
高传彩,惠基运*,魏玉兰,张蕊,刘建廷,肖伟**,李玲**
干旱及复水对‘红富士’苹果生长及果实品质和产量的影响
高传彩1,2,3,惠基运1,2,3*,魏玉兰4,张蕊1,2,3,刘建廷1,2,3,肖伟1,2,3**,李玲1,2,3**
1. 山东农业大学园艺科学与工程学院, 山东 泰安 271018 2. 山东果蔬优质高效生产协同创新中心, 山东 泰安 271018 3. 作物生物学国家重点实验室, 山东 泰安 271018 4. 临沂市平邑县吴台镇农业综合服务中心, 山东 平邑 273302
本文以多年生‘红富士’ (Borkh. ‘Red Fuji’) /M26为试材,设置正常灌溉、持续干旱和干旱不同天数后复水三组处理,探究不同程度干旱对‘红富士’生理生化特性的影响以及果实品质和产量随干旱时间延长的变化规律。结果表明:随干旱处理时间延长,叶片光合性能、叶绿素含量呈先升后降趋势;超氧化物歧化酶(SOD)活性及可溶性蛋白含量也表现出先升后降;过氧化物酶(POD)活性和脯氨酸(Pro)含量呈上升的趋势,以上指标在干旱胁迫60~70 d之间变化最剧烈;干旱胁迫引起植株产量下降,但轻度干旱胁迫有利于果实着色,使果实颜色更深,干旱20 d后进行复水处理的植株其果实品质保持在较高水平。
干旱; 复水; 苹果; 果实品质
我国是农业大国,但是全国面积的一半左右是干旱半干旱区[1],很多地区水资源匮乏,作物不能及时地补水,造成产量的巨大损失。因此,干旱问题必须得到我们足够的重视。干旱胁迫可能导致渗透平衡和氧化还原失衡、光合作用受损和细胞能量耗尽[2]。本文研究了干旱对苹果光合、抗逆相关生理生化指标和产量、果实品质等的影响,探究苹果成龄树干旱致死的生理生化过程、致灾机理、影响关键期,以期为生产上应对干旱灾害提供理论参考。
1 材料与方法
1.1 试验材料
2019年在山东农业大学国家苹果工程技术研究中心开展本试验,供试材料为39棵长势一致,生长良好的树龄12年的‘红富士’(Borkh.‘Red Fuji’) /M26苹果植株。采用行间生草加起垄栽培,树形为小冠疏层形,株行距为2 m×4 m,年产量约为42 t·hm-2。5月中旬选取30棵植株进行简易防雨设施的搭建,将塑料薄膜置于树冠之下,高度大约与树干一致,上部用绳子系于树体主枝或铁丝上,下部用木棍进行支撑,进行干旱及复水处理;剩余9棵植株正常灌溉。
1.2 材料的处理
1.2.1对照处理选取9棵正常浇水的试材为对照(CK),浇水频率及浇水量视土壤的干旱状况而定。每隔10 d测定对照组植株光合参数并取样监测其他指标的变化规律,同时测定土壤含水量。
1.2.2干旱与复水处理于6月上旬对30棵试材充分灌溉后停止供水进行干旱处理,为干旱处理的起点。选取9棵植株持续干旱处理(T),每隔10 d测定光合参数并取样监测其他指标的变化规律,同时测定土壤含水量。剩余21棵植株用于复水处理(Rew),分别对干旱胁迫20、30、40、50、60、70 d的植株进行复水,分为6个处理Rew1、Rew2、Rew3、Rew4、Rew5、Rew6,每处理选取3棵植株,观察其生长状况,待植株果实成熟后测定复水处理植株果实品质和产量等指标。
1.3 测定指标与方法
选取树体中部外围枝条中部成熟叶片后用去离子水清洗干净后将水分擦拭干,每次取样将样品剪碎并混匀,每个指标重复3次,称重后迅速在液氮中冷冻,置于-80 °C冰箱中备用。
1.3.1叶绿素含量的测定采用分光光度法测定叶片叶绿素含量。
1.3.2土壤含水量的测定在灌水前及灌水后48 h采用浙江托普仪器有限公司的手持式土壤水分温度速测仪(型号为TZS-IW,测定探头的长度为10 cm左右),在树垄上挖去20 cm的土层后插入探测头进行测定,重复5次。
1.3.3光合指标的测定使用CIRAS-3光合作用仪(PP-Systems, 美国)随机选择植株枝条中部成熟叶片,测定净光合速率(photosynthetic rate, P),气孔导度(stomatal conductance,G)和蒸腾速率(transpiration rate,T)和细胞间CO2浓度(intercellular CO2concentration,C),重复5次,测定时间为上午9:00~11:00。使用LED红色和蓝色光源进行测量,将光强度设置为1200 μmol·m−2·s−1,使用开放式空气路径系统,叶室的温度为25 °C,叶室的面积为18×25 mm。
1.3.4保护酶活性、可溶性蛋白、游离脯氨酸含量的测定NBT光还原法[3]测定超氧化物歧化酶(superoxide dismutase, SOD)活性;愈创木酚法[4]测定过氧化物酶(peroxidase, POD)活性。可溶性蛋白含量参照考马斯亮蓝G-250染色法[5]测定;丙二醛(alternative oxidase, MDA)、游离脯氨酸含量测定参照赵世杰等[6]的方法进行测定。
1.3.5 果实品质、产量的测定果实硬度使用GY-1型果实硬度计测量,可溶性固形物使用折光仪测量,可滴定酸、固酸比和果实组织含水量使用曹建康等[7]的方法,每个处理每次测量15个果实。产量的统计按照株产及株行距求出亩产,再根据亩产算出一公顷的产量,单位为kg·hm-2。
1.4 数据统计与分析
采用Microsoft Office Excel 2012进行数据处理,采用Graphpad Prism 8.3.0软件作图,采用IBM SPSS Statistics 21对数据进行显著性和差异性分析(<0.05)。
2 实验结果
2.1 土壤含水量的变化及复水后植株的生长状况
处理过程中土壤的含水量变化情况如图1所示。对照组土壤含水量变化较小,处理组的土壤含水量随干旱时间的延长不断降低,从干旱处理后20 d开始显著低于对照组。
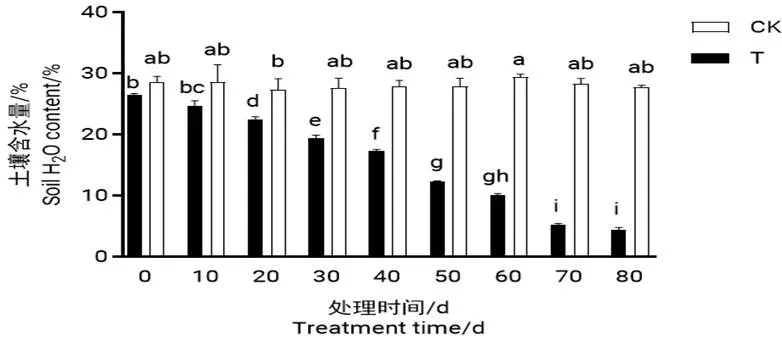
图 1 干旱过程中土壤含水量的变化
注:同一指标数据用不同小写字母标识表示数据间差异显著(<0.05),下同。
Note: The significant differences (P<0.05) between data of the same index are indicated by different lower cases,the same below.
干旱处理后20 d开始,每隔10 d选取3棵植株进行复水处理,观察其能否恢复正常生长。结果发现,干旱处理第60 d土壤含水量降为10.05%,在此之前复水的植株,能恢复生长;干旱处理第70 d土壤的含水量降为5.17%此时解除胁迫的植株并不能恢复正常生长。处理组植株在干旱处理第80 d出现大量落叶、叶片干枯的现象,植株接近死亡,为干旱胁迫的终点。
2.2 不同程度的干旱对‘红富士’叶片光合参数的影响
研究结果(图2)表明,干旱处理前期,处理组植株叶片各项光合参数与对照组无显著差异,从第30 d开始,处理组P均显著低于对照,第80 d达到最低值并趋于零(图2-A);第40 d开始,处理组T不断降低(图2-B);自干旱处理后40 d开始,处理组C逐渐升高,且显著高于对照(图2-C);s在干旱过程中整体呈降低趋势,第50 d开始处理组叶片G迅速降低,并明显低于对照。
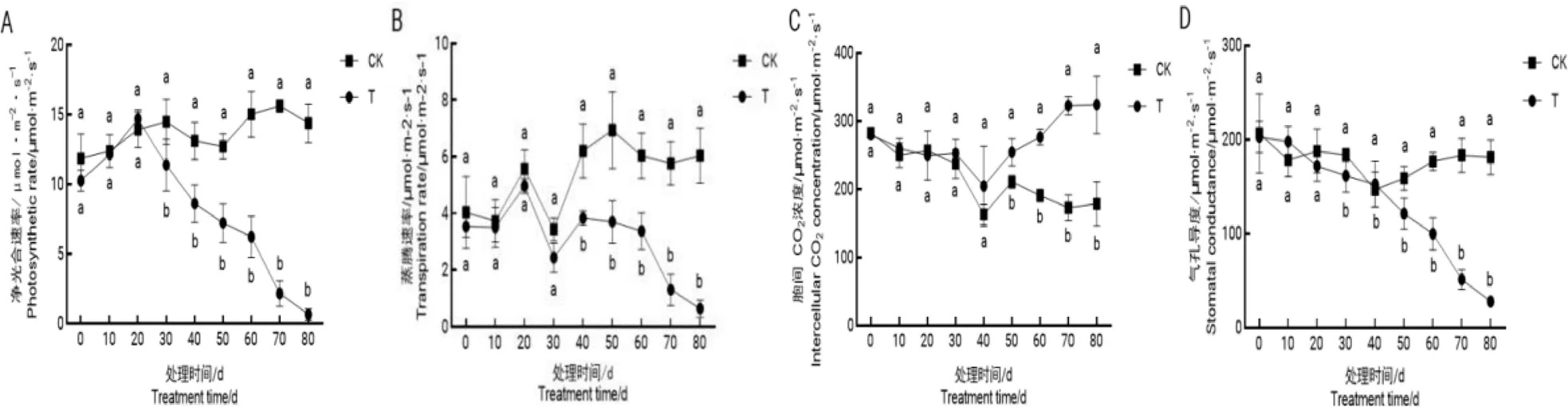
图 2 不同程度干旱对‘红富士’叶片光合参数的影响
光合参数的变化说明干旱前期对‘红富士’苹果光合作用影响甚微,甚至出现了处理组光合参数优于对照组的情况,在干旱处理中后期植株受到的胁迫影响逐渐增大,干旱胁迫后60~70 d光合参数变化幅度最大。
2.3 不同程度的干旱对‘红富士’叶片叶绿素含量的影响
研究结果(图3)表明,干旱处理0~20 d,处理组叶片的叶绿素含量高于对照;此后,处理组叶片叶绿素含量不断减少,显著低于对照组,在干旱处理第50~70 d大幅减少,到第80 d叶绿素含量降为最小值。叶绿素含量的变化说明,轻度干旱有利于‘红富士’苹果叶片叶绿素的合成,但随胁迫的加重,植株受到的伤害加剧,导致叶绿素含量显著下降。

图 3 不同程度干旱对‘红富士’叶片叶绿素含量的影响
2.4 不同程度的干旱对‘红富士’叶片抗氧化酶活性的影响
SOD活性在干旱过程中呈现出先上升后降低的单峰趋势,干旱处理第20 d,SOD活性达到了最高值;此后处理组SOD活性开始降低,到干旱胁迫第50 d,SOD活性依然高于对照,第60 d开始SOD活性显著低于对照,60~70 d下降幅度较大(图4-A)。POD活性在干旱处理第0~50 d不断升高,此后呈波浪式的变化,在干旱处理后70 d活性最高,约为对照的3.73倍(图4-B)。SOD和POD活性在干旱过程中的变化幅度较大,说明它们在‘红富士’植株应对干旱胁迫的过程中发挥了重要作用;干旱胁迫第70 d SOD活性降幅较大,POD活性在此时达到最高值,说明干旱已对植株产生较大影响。

图 4 不同程度的干旱对‘红富士’叶片抗氧化酶活性的影响
2.5 不同程度的干旱对‘红富士’叶片渗透调节物质含量的影响
研究(图5-A)表明,干旱处理过程中,对照组叶片脯氨酸(proline, Pro)含量始终维持在较低水平,而处理组植株叶片Pro含量不断上升。处理第0~40 d叶片Pro含量上升幅度较为平缓,第50~70 d迅速上升,第70 d达到对照组的16.05倍。干旱过程中‘红富士’苹果叶片Pro含量的变化趋势表明,植株在干旱初期受影响较小,干旱胁迫中后期尤其是干旱处理后70 d,植株对胁迫程度反应剧烈。

图 5 不同程度的干旱对‘红富士’叶片渗透调节物质含量的影响
研究(图5-B)表明,叶片可溶性蛋白在干旱过程中整体呈先增加后减少的趋势。第40 d达到最高值,相比对照提高了74.15%;干旱处理第60 d可溶性蛋白降至与对照水平一致,第70 d开始显著低于对照。干旱过程中‘红富士’叶片可溶性蛋白含量变化表明,植株在干旱初期为应对干旱胁迫,合成大量的可溶性蛋白提高植株保水能力;干旱胁迫后期尤其是干旱处理后第70 d,植株受到的伤害加剧,引起可溶性蛋白含量降低。
2.6 干旱不同程度后复水对‘红富士’果实品质、产量的影响
2.6.1复水处理对‘红富士’果实外观品质的影响试验中对植株进行干旱后复水(Rew)处理并测定植株果实品质。由图6-A可以看出,对照组的果皮颜色较干旱后复水组果皮颜色更加鲜亮,且表皮整洁光滑,干旱20 d后复水的植株果实表面较为光滑,干旱30 d及以后解除胁迫的植株果实表皮较对照粗糙;干旱处理后30 d解除胁迫果皮颜色最深,偏紫红色,其花青素含量也最高(图6-B),此时土壤含水量为19.46%,干旱30 d以后解除胁迫的植株果实花青素含量明显降低,显著低于对照。随干旱胁迫的加重,植株果实的类黄酮和总酚含量呈上升趋势,且两者的含量均在干旱处理60 d后显著高于对照。
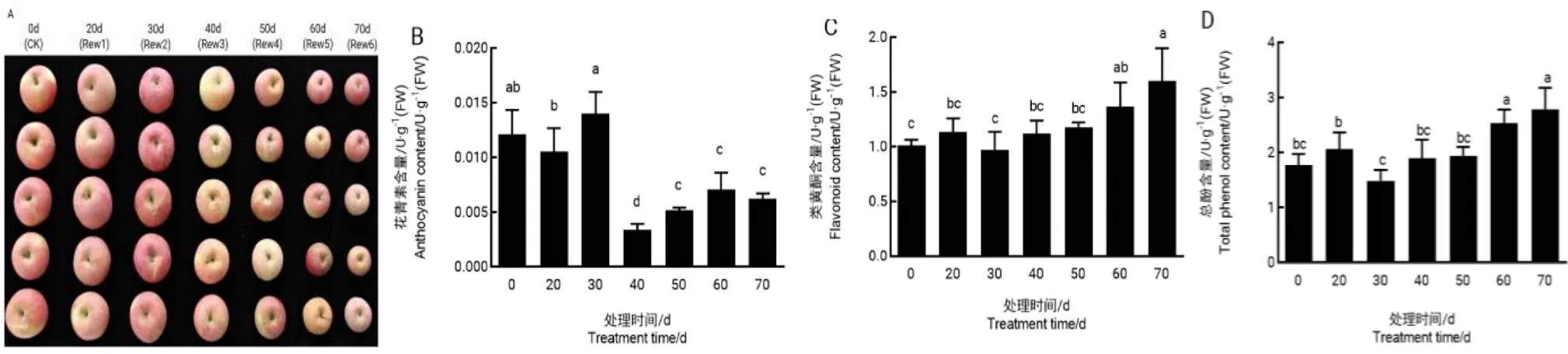
图 6 干旱不同程度后复水对‘红富士’果实外观品质的影响
2.6.2 复水处理对‘红富士’果实内在品质的影响结果表明(如表1),随干旱时间的延长,果实硬度逐渐变大,干旱50 d及以后复水的果实硬度显著高于对照。可溶性固形物含量先升后降,干旱后20 d复水植株果实可溶性固形物的含量最高,但与对照差异不显著;干旱30 d及更长时间解除胁迫的植株果实可溶性固形物含量不断降。可滴定酸含量整体表现为先升高后下降,干旱50 d复水植株果实可滴定酸含量最多,此后越来越低。固酸比能够反应果实口感的酸甜,其比值的高低取决于可溶性固形物和可滴定酸的含量,随着干旱时间的延长,植株果实固酸比先升高后降低又升高,干旱后20 d复水植株果实固酸比最高(此时土壤含水量为21.83%);果实组织含水量在不同干旱程度下没有表现出显著性差异。
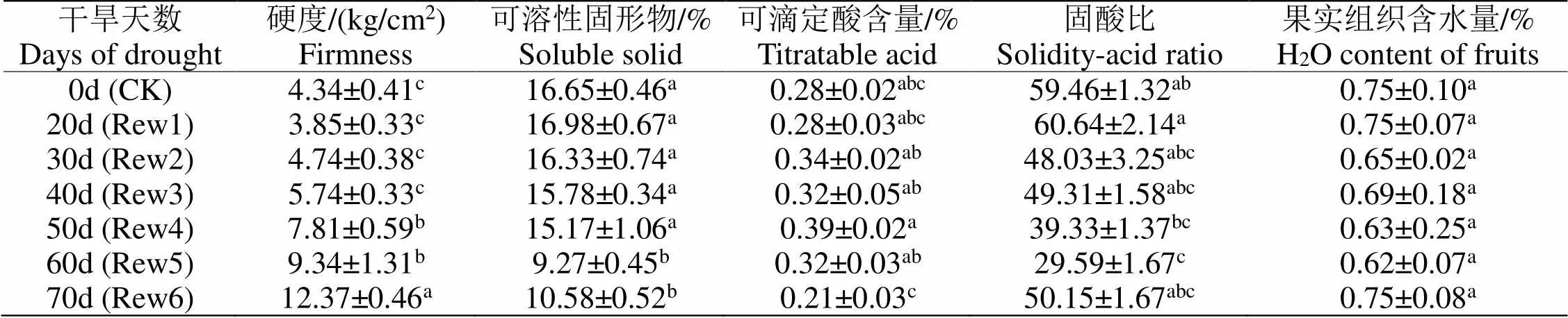
表 1 干旱不同程度后复水对‘红富士’果实内在品质的影响
*每列中字母相同者表示差异未达显著水平(>0.05),字母不同者表示差异达显著水平(<0.05),下同。
*The identical of letter indicate for no significant difference existence (>0.05); different letters indicate for the significant difference (<0.05),the same below.
2.6.3 干旱不同程度后复水对‘红富士’产量的影响在果实生长发育的过程中不断进行着细胞数量的增加和体积的增大,这个过程的进行离不开水分的参与,因此,干旱会对产量造成的影响。研究表明(如表2),干旱后20 d复水植株单果重略高于对照,差异不明显,从干旱第40 d开始,解除胁迫的植株单果重迅速降低并明显低于对照。果实纵横径的大小都随着干旱时间的延长总体呈降低趋势,由于纵横经的下降幅度的不一致,导致果形指数表现为先降低后升高第60 d又开始降低。干旱对果实产量影响较大,Rew组植株产量均显著低于对照,干旱胁迫40 d后复水植株产量下降幅度最大。
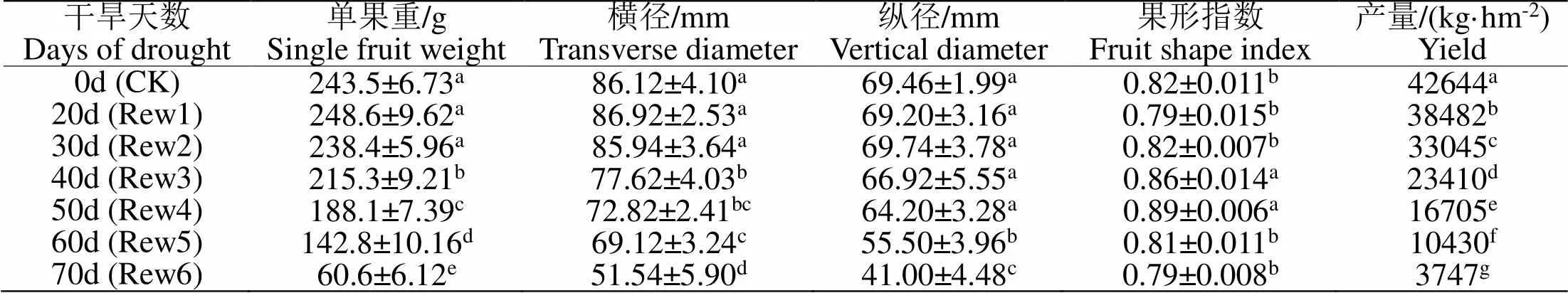
表 2 干旱不同程度后复水对‘红富士’产量的影响
3 讨 论
3.1 不同程度的干旱对‘红富士’叶片光合参数及叶绿素含量的影响
光合作用对植物的生长发育至关重要,植物通过叶片制造有机物来维持自身正常的生命活动。干旱条件下,植物表现出光合速率下降和叶绿素的含量下降[8]。本试验中,处理组净光合速率呈先升高后降低的趋势,这可能是由于前期适度干旱叶片叶绿素含量增加导致。张栋[9]对一年生苹果进行不同程度的水分胁迫处理,发现中度胁迫下光合的主要限制因素是气孔因素,随着胁迫的加剧,非气孔因素占主导。本试验中,C在干旱处理过程中先下降后上升,干旱处理前40 d呈下降趋势是由于在轻度干旱条件下植物所受到的伤害有限,只是通过关闭气孔降低蒸腾减少自身水分的散失来抵御干旱,由于气孔的关闭导致C降低[10];C在干旱处理40 d以后显著高于对照,这是因为此时植物遭受到严重的干旱胁迫,限制光合作用的因素变为非气孔限制因素,因为植物遭受的逆境伤害已经破坏了光合器官的结构[11],C浓度升高,与P和G的变化相反。
植物叶片叶绿素含量对干旱胁迫非常敏感。试验中,对照组植株叶片叶绿素含量先升高后降低,这与杨晓玲等[12]对二年生‘新红星’的研究结果一致,说明轻度的干旱有利于成龄树叶绿素的合成,这恰好能反应干旱前期‘红富士’植株的光合作用没有受到抑制。干旱处理30 d开始叶绿素含量持续减少,干旱胁迫引起叶绿素含量的降低一方面是由于叶绿素的分解加强,合成减少,还可能由于干旱损害了叶绿体的结构从而导致叶绿素含量的降低[13,14]。在干旱处理50~70 d (土壤含水量为12.38~5.17%)时叶绿素含量急剧下降,说明此时植株遭受的干旱伤害已经破坏了光合器官的结构。
3.2 不同程度的干旱对‘红富士’叶片抗氧化酶活性的影响
大量研究结果表明许多保护酶与植物抗旱性密切相关,其中SOD、POD等与植株的抗旱能力成正相关[15]。当水分胁迫的程度较轻、胁迫速度较慢时,植物体内的抗氧化酶SOD、POD等活性会增加,降低胁迫对植株造成的伤害;当干旱程度较重、速度较快、持续的时间较长时,保护酶的活性会降低,导致植物遭受严重的逆境伤害[16]。但阎腾飞等[17]对多年生富士苹果幼树进行水分胁迫发现POD活性在处理末期会升高。本试验中,处理组植株叶片SOD活性呈先升高后下降的趋势,干旱处理第20~50 d SOD活性均高于对照;植株的POD活性呈不断升高后趋于稳定的趋势。后期POD活性的升高可能是由于其发挥作用的时间在SOD之后,即二者存在补偿机制[18,19]。
3.3 不同程度的干旱对‘红富士’叶片渗透调节物质含量的影响
渗透调节也是植株抵抗水分胁迫的重要途径。Pro是植株体内最关键的渗透调节物质之一。前人研究表明,多种果树在干旱胁迫时体内会累积大量的Pro[15,20]。本研究中,处理组植株叶片Pro含量随干旱程度的加剧不断增加;干旱胁迫下,成龄树的Pro含量在干旱前40 d增加缓慢,此后迅速积累,第70 d(土壤含水量为5.17%)增加幅度最大,即此时受到的逆境伤害明显增加。
可溶性蛋白也是植株关键的渗透调节物质之一。试验中,干旱胁迫下‘红富士’植株叶片可溶性蛋白含量先升高后降低,与阎腾飞等[17]研究结果相似。干旱前期可溶性蛋白增加较缓慢,说明植株需要一定程度的干旱胁迫才能诱导可溶性蛋白的大量合成,后期蛋白质含量的降低是由于干旱使植物体内产生的有害物质抑制了蛋白质的合成,加速了蛋白质的分解。
3.4 干旱不同程度后复水对‘红富士’果实品质及产量的影响
干旱胁迫是限制果树生长和果实产量的重要非生物胁迫之一。适度干旱可以提高糖代谢相关酶活性,增加果实的可溶性固形物、可溶性糖等的含量,使果实的内在品质提高[21]。试验中,干旱20 d后复水的植株果实的可溶性固形物含量升高,到干旱60 d解除胁迫后,其果实可溶性固形物含量显著低于对照,可能由于干旱胁迫过重影响了糖代谢相关酶活性,这和阳姝婷[22]的研究结论一致。张兴涛等[23]认为干旱能提高果实硬度,本试验的研究结果与其一致。有研究结果表明,水分亏缺可提高果实有机酸含量[24],但也有研究者得出相反的结论[25]。本研究中,与对照相比,干旱20 d之后解除胁迫的植株果实可滴定酸增加,在干旱70 d解除胁迫后可滴定酸又显著降低。干旱胁迫对果实的外观品质尤其是果实着色会产生一定的影响,本试验得出,轻度干旱胁迫有利于果实着色,使果实的颜色更深,与阳姝婷[22]的研究结果一致。
果实体积的增大包括细胞数目及体积的增加,这都需要充足的水分供应。阳姝婷[22]对甜樱桃的干旱试验得出,随着土壤水分的不断降低,甜樱桃的产量显著降低,果实纵横经减小,果形指数增加。本试验中,随着干旱胁迫时间的延长,干旱20 d进行复水的植株果实单果重、纵横径稍有增加,20 d后解除胁迫的植株以上三个指标均不断降低,果形指数随着干旱时间的延长整体呈先增大后减小的趋势,中期的增大说明横径的下降幅度大于纵径;‘红富士’植株的产量随解除胁迫起点的延后不断降低,干旱后复水组植株产量均显著低于对照,这应该是由于干旱胁迫引起不必要的落果导致的。干旱20 d后进行复水的植株果实可溶性固形物、固酸比、单果重都达到了所有处理的最高值,其类黄酮、总酚及可滴定酸含量也保持较高水平,适度的干旱提高了‘红富士’苹果果实品质。
4 结 论
随干旱处理时间的延长,‘红富士’叶片光合性能、叶绿素含量呈先升后降趋势;超氧化物歧化酶(SOD)活性及可溶性蛋白的含量也表现出先升后降;而过氧化物酶(POD)活性和脯氨酸(Pro)含量呈上升的趋势,以上指标在干旱胁迫60~70 d之间变化最剧烈;干旱胁迫引起植株产量下降,但是轻度干旱胁迫有利于果实着色,使果实的颜色更深,干旱20 d后进行复水处理的植株其果实品质保持在较高水平。
[1] 杨阳,申双和,马绎皓,等.干旱对作物生长的影响机制及抗旱技术的研究进展[J].科技通报,2020,36(1):8-15
[2] Golldack D, Li C, Mohan H,. Tolerance to drought and salt stress in plants: unraveling the signaling networks [J]. Front Plant Sci, 2014,5:151
[3] 高俊凤.植物生理学实验指导书[M].北京:高等教育出版社,2006
[4] 孙群,胡景江.植物生理学研究技术[M].杨陵:西北农林科技大学出版社,2006
[5] 王学奎.植物生理生化实验原理和技术[M].第2版.北京:高等教育出版社,2006
[6] 赵世杰,史国安,董新纯.植物生理学实验指导[M].北京:中国农业科学技术出版社,2002
[7] 曹建康,姜微波,赵玉梅.果蔬采后生理生化实验指导[M].北京:中国轻工业出版社,2007
[8] 欧芷阳,庞世龙,谭长强,等.干旱胁迫对桂西南石漠化地区主要造林树种光合与耗水特性的影响[J].生态学杂 志,2020,39(10):3237-3246
[9] 张栋.干旱胁迫对苹果光合作用和叶绿素荧光的影响及叶片衰老特性研究[D].杨凌:西北农林科技大学,2011
[10] 江天然,张立新,毕玉蓉,等.水分胁迫对梭梭叶片气体交换特征的影响[J].兰州大学学报,2001,37(6):57-62
[11] 肖冬梅,王淼,姬兰柱.水分胁迫对长白山阔叶红松林主要树种生长及生物量分配的影响[J].生态学杂志,2004,23(5):93-97
[12] 杨晓玲,鲁玉哲,杜香华.干旱胁迫下新红星苹果叶片一些生理指标的变化[J].河北农业技术师范学院学报,1996,10(3):23-26
[13] 郭爱霞,石晓昀,王延秀,等.干旱胁迫对3种苹果砧木叶片光合、叶绿体超微结构和抗氧化系统的影响[J].干旱地区 农业研究,2019,37(1):178-186
[14] 王顺才,邹养军,马锋旺.干旱胁迫对3种苹果属植物叶片解剖结构、微形态特征及叶绿体超微结构的影响[J].干旱 地区农业研究,2014,32(3):15-23
[15] 陈立松,刘星辉.作物抗旱鉴定指标的种类及其综合评价[J].福建农林大学学报(自然版),1997,26(1):48-55
[16] 赵丽英,邓西平,山仑.活性氧清除系统对干旱胁迫的响应机制[J].西北植物学报,2005,25(2):413-418
[17] 阎腾飞,孙耀清,李文扬,等.水分胁迫对富士苹果幼树光合特性以及生理生化指标的影响[J].信阳农业高等专科学校学报,2013,23(4):78-84
[18] 张虎,范俊俊,张往祥.淹水胁迫对湖北海棠叶片生理指标的影响及胁迫相关基因的表达[J].东北林业大学学报,2019,47(10):30-35,63
[19] 汤玉喜,刘友全,吴敏,等.淹水胁迫对美洲黑杨无性系保护酶系统的影响[J].中南林业科技大学学报,2008,28(3):1-5
[20] 张云贵,谢永红,吴学良,等.PEG诱导水分胁迫对柑桔幼苗细胞质膜透性及脯氨酸含量的影响[J].果树科学,1995(S1):25-28
[21] 王元基.干旱对苹果品质的影响及其与糖代谢的关系[D].杨凌:西北农林科技大学,2017
[22] 阳姝婷.干旱胁迫对甜樱桃生理及果实品质的影响[D].雅安:四川农业大学,2016
[23] 张兴涛.节水灌溉处理对苹果碳代谢及产量和品质的影响[D].晋中:山西农业大学,2013
[24] 马福生,康绍忠,王密侠,等.调亏灌溉对温室梨枣树水分利用效率与枣品质的影响[J].农业工程学报,2006,22(1):37-43
[25] 崔宁博,杜太生,李忠亭,等.不同生育期调亏灌溉对温室梨枣品质的影响[J].农业工程学报,2009,25(7):32-38
Effects of Drought and Rehydration on the Growth, Fruit Quality and Yield of Red Fuji Apple
GAO Chuan-cai1,2,3, HUI Ji-yun1,2,3*, WEI Yu-lan4, ZHANG Rui1,2,3, LIU Jian-ting1,2,3, XIAO Wei1,2,3**, LI Ling1,2,3**
1.271018,2.271018,3.271018,4.273302,
The perennial (Borkh. ‘Red Fuji’) /M26 is applied as test material. Simultaneously, one control group, normal irrigation (CK) along with two experimental groups, continuous drought (T) and rehydration after different drought period (Rew) were set to explore the effects of different levels of drought on physiological and biochemical characteristics of perennial ‘Red Fuji’, as well as to understand the pattern of fruit quality and yield changing impacted by diverse drought duration times. The study manifested that the photosynthetic characteristics and chlorophyll content of tested leaves initially increased and then decreased with the extension of drought stress duration. The superoxide dismutase (SOD) activity and soluble proteins content of tested materials also presented the similar tendency over drought stress time. Nevertheless, a significant rising tendency in the process of drought stress treatment was identified for the activity of peroxidase (POD) and content of proline (Pro), which experienced the most intense change between 60 and 70 days of drought stress. Moreover, low-level drought stress was characterized to conduce to fruit coloring. Fruit quality of plants that were rehydrated at the day after 480h of drought stress treatment remained at a comparative crucial higher level.
Drought; rehydration; apple; fruit quality
S661.1
A
1000-2324(2021)02-0194-07
10.3969/j.issn.1000-2324.2021.02.006
2020-12-11
2021-02-24
国家重点研发计划:重大自然灾害监测预警与防范(2017YFC1502805);山东省现代农业产业技术体系果品创新团队项目(SDAIT-06-01);山东省重大科技创新工程(2018CXGC0209)
高传彩(1993-),女,硕士在读,主要从事果树生理研究工作. E-mail:18353231375@163.com
惠基运(1995-),男,博士在读,主要从事果树逆境生理研究工作. E-mail:18562318721@163.com
Author for correspondence. E-mail:gulight986918@163.com;lilingsdau@163.com
猜你喜欢
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
中国果业信息(2021年12期)2021-12-02
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
少儿科学周刊·少年版(2021年20期)2021-01-17
农家科技(2020年3期)2020-05-11
小溪流(故事作文)(2018年11期)2018-12-26
幸福·悦读(2016年8期)2016-10-21
哈尔滨理工大学学报(2016年2期)2016-09-12
农家顾问(2016年7期)2016-05-14
