
雌雄同株黄檗有性繁殖特性研究
2021-05-27 05:59:58范永芳魏雪苹贺利宏
中国农学通报 2021年15期
范永芳,徐 硕,张 昭,于 晶,魏雪苹,贺利宏
(中国医学科学院北京协和医学院药用植物研究所,北京 100193)
0 引言
芸香科黄檗属植物黄檗为传统意义上的雌雄异株植物。但笔者在2018年首次发现了多株雌雄同株黄檗,称之为两性植株,此前未曾有过关于黄檗性别可塑性报道。这些两性植株主干在幼年时上端死亡,于15.8~30.7 cm处分杈,形成不同性别的分枝,雌枝平均周长31.1 cm,雄枝平均周长23.2 cm。经过连续3年观察,发现不同性别分枝性别表达稳定,未发生性别转换。之前也有少量的研究报道过在雌雄异株居群中发现罕见的两性植株个体,两性植株个体的性别表达方式多为在雄株上发现少量雌花,如胡杨[1]、银杏[2]、连香树[3]和红豆杉[4];或是雄花和雌花无规律的分布在植株上,如黄连木[5]。笔者所发现的两性黄檗的性别表达方式与上述有所不同,不同性别分枝的性别表达方式在以往研究中也未见报道,两性黄檗植株的发现丰富了植物性别表达方式的多样性。
两性黄檗植株主干均较短,分枝分化为不同性别。可能是由于北京冬季天气干冷导致幼苗上部干枯,侧芽分化为不同性别而形成的。笔者推测,黄檗的性别由表观遗传方式决定,在年幼时植株可能不存在性别,在一定的树龄才分化出性别,不同的分枝分化为不同的性别,这在植物中还未曾报道过。但在动物研究中早就发现,低等动物中的两栖类和鱼类,在生长发育的某一阶段形成性别并维持稳定,如蜥蜴、鳄鱼和龟鳖,在孵化前的某一时期受温度影响并以表观遗传的方式分化出不同的性别[6-7]。也有可能黄檗在年幼时已有性别分化,但性别表达并不稳定,在一定树龄时稳定表达,不同分枝稳定表达为不同性别。如槭树等植物在树龄较小时植株偏向于雄性表达,较大时偏向于雌性表达[8-10]。两性植株黄檗的发现,为研究植物性别决定与分化提供了新的思路和材料。
黄檗的两性植株在同一植株上同时拥有大量的雄花和雌花,可以克服较远距离花粉传播的困难,这对于黄檗野生资源保护和种群恢复具有重要意义。黄檗曾遍布于欧亚及美洲大陆,几经变迁后龟缩于亚洲东部部分地区[11-12],根据《中国植物志》记载,现黄檗主要分布于中国东北地区、华北部分省区及朝鲜、日本和俄罗斯[12]。在20世纪时野生黄檗资源遭到了滥砍乱伐,栖息地受到严重破坏,导致野生资源零星分布,集中分布面积极小[11,13-14]。根据张志鹏的调查,发现野生黄檗主要分布在长白山、张广才岭和小兴安岭南部等地,较之原来的数据,分布范围进一步缩小[14-15]。调查还发现野生黄檗分布呈现严重的碎片式和零星化,在燕山、千山、长白山、张广才岭和小兴安岭5条山脉的38个样点中仅发现966株野生黄檗,平均种群密度只有25.4株/(km·m)[15]。这种分布模式对于雌雄异株的黄檗而言,花粉远距离传播障碍难以克服,不利于繁衍和种群的自我更新。黄檗作为第三纪古热带植物区系孑遗植物,对于研究物种进化、古植物区系以及古气候变化有重要意义[14,16-17],且黄檗是大宗药材“关黄柏”的基源植物和优良的木材原料,对于经济的发展也有重要意义,因此保护黄檗资源刻不容缓。黄檗已经先后被列入《中国珍稀濒危保护植物名录》和《国家重点保护野生植物名录》,成为国家级保护物种[18],有关部门已经对野生黄檗采取了强有力的保护措施。但由于野生资源的严重破坏,其繁衍和自我更新面临极大困难[11,19-20]。两性植株的出现可以弥补这一缺失,克服黄檗碎片式分布导致的授粉困难[11,14,21],在一定程度上有利于野生黄檗植株进行成功的有性繁殖,使笔者对野生黄檗种群恢复增加了更多的希望。
繁殖是生物生存的关键[22],两性黄檗植株能否和雄株和雌株一样正常执行雄性功能和雌性功能具有重要意义,也为进一步的实验提供基础,因此笔者分别对两性植株的雄性功能和雌性功能进行了研究。花粉和柱头在有性繁殖中具有重要作用,是传粉系统的重要组成[23],其表观形态对授粉具有较大的影响,笔者首先对黄檗两性植株花粉和柱头的形态和花粉活力进行观察。并于2019年和2020年连续2年通过人工授粉和自然受粉方式,对两性植株雄性繁殖能力、雌性繁殖能力和自交繁殖能力3个方面进行评估,以描述两性植株的繁殖能力。
1 材料与方法
1.1 材料
两性黄檗植株为北京药用植物园(40.03°N;116.27°E)保育黄檗个体。研究涉及4株个体,2株为2005年北京种源播种,2株为1999年从吉林省移植;对照植株均为同源保育材料中的单性植株个体。物种由中国医学科学院药用植物研究所张昭研究员鉴定为芸香科黄檗属植物黄檗(PhellodendronamurenseRupr.)。实验在北京药用植物研究所中草药物质基础与资源利用教育部重点实验室,于2018年5月—2020年10月进行。
1.2 方法
1.2.1 两性植株雄性功能研究
(2)花粉形态观察。将干燥后的花粉置于金属靶上,JFC-1600离子溅射仪镀金膜,用JSM-6510LV扫描电子显微镜以合适的倍数观察花粉形态和表面纹饰,测量花粉极轴和赤道轴长度。两性植株和雄株花粉各测量49个。每种花粉选取10个×200倍数的视野,观察并记录正常花粉和畸形花粉的数量,计算花粉的畸形率。
(3)花粉活力检测。固体培养基培养法观察花粉萌发率,采用肖美林等人的培养基:10%蔗糖+0.008%硼酸+0.02%硝酸钙+5g/L琼脂[24];用毛笔蘸取花粉,均匀涂抹在培养基表面,放入25℃培养箱中,培养24 h后进行观察。共设置光照、黑暗和光照/黑暗(2h/2h)3种培养条件处理,每个处理3个重复。在体视显微镜下观察,每个培养皿选取5个视野,观察并记录花粉萌发情况,计算花粉萌发率。
(4)雄性繁殖能力研究。分别在2019年和2020年通过人工授粉方式,花粉为雌株授粉,以雄株花粉为对照,统计果实和种子数量判断两性植株雄性繁殖能力。在开花前2日给雌花序套袋,从雌花序第一朵花开放开始,每日进行授粉,直至最后一朵花授粉后结束,在群体花期结束后去袋。两性植株花粉共授粉51个花序,1785朵雌花;雄株花粉共授粉51个花序,1954朵雌花。果实成熟时采摘果序,记录果实数以及单果中种子数。
1.2.2 两性植株雌性功能研究
(1)柱头形态观察:采集花蕾期、开花1天和开花2天的雌花,FAA固定液[25]固定。梯度乙醇脱水,乙醇-叔丁醇逐级替换直至100%叔丁醇,ALPHA2-4LD PLUS冷冻干燥器干燥[25-27],干燥后的柱头于金属靶上,JFC-1600离子溅射仪镀金膜,用JSM-6510LV扫描电子显微镜以合适的倍数观察柱头形态,并拍照记录,以雌株的柱头为对照进行比较。
(2)雌花繁殖能力研究:分别在2019年和2020年,通过人工授粉的方式,以雄株花粉为两性植株的雌花和雌株的花授粉,授粉方式同上。两性植株授粉19个花序,762朵雌花;雌株授粉19个花序,680朵雌花。待果实成熟后采摘果序,记录果实数以及单果中种子数。
目前洋地黄毒苷临床已很少应用,所以市面上很难买到。为保留该实验项目,我们对实验进行了改进。用维拉帕米(钙通道阻滞剂,临床主要用于治疗心律失常)[4]作为取代药,以异丙肾上腺素为工具药来区分普萘洛尔和维拉帕米对心脏的抑制作用及其作用机制。
2020年采集两性植株和雌株于自然受粉条件下生长的果序进行比较,记录开花数、果实数以及单个果实中种子数,两性植株采摘19个果序,共767个果实,雌株采摘19个果序,共672个果实。
1.2.3 两性植株自交繁殖能力研究 分别在2019年和2020年,通过人工授粉的方式,以两性植株的花粉对该植株雌花进行授粉,共授粉19个花序,523朵花;以雄株花粉对两性植株雌花授粉为对照,共授粉19个花序,762朵花,授粉方式如上,待果实成熟后采摘果序,记录果实数以及单果中种子数。
1.2.4 数据分析 使用Excel和SPSS软件对数据进行整理,采用独立样本T检验和非参数检验分析。
2 结果与分析
2.1 两性植株雄性功能研究
2.1.1 花粉形态 共观察654个两性植株花粉,760个雄株花粉,发现花粉均为单粒三沟花粉,萌发沟延伸到两极,赤道面观呈长圆形,极面观呈三叶圆形(图1)。根据王开发等[28]对花粉形状类型的分类标准,这2种花粉均属于长球形(1.14<P/E<2)。花粉外壁被网状纹饰覆盖,并有点状凸起装饰网格内部。

图1 两性植株和雄株花粉的亚显微结构
随机选择两性植株和雄株花粉各49个,测量花粉极轴和赤道轴长,详见表1。两性植株花粉极轴长平均为40.93±1.75 μm,赤道轴长平均为22.67±1.21 μm,P/E值平均为1.81±0.10。雄株花粉的极轴长平均为40.89±1.76 μm,赤道轴长平均为22.81±1.27 μm,P/E值平均为1.80±0.12。统计分析结果表明,2种花粉在极轴长、赤道轴长和P/E上均没有差异,因此两性植株花粉在大小上与雄株花粉发育情况相同。
观察并记录10个视野中总花粉数量和畸形花粉数量,详见表1。两性植株花粉的畸形率平均为13.89±5.31%,雄株花粉的畸形率平均为10.02±3.80%。通过统计分析发现,两性植株花粉的畸形率与雄株花粉相比没有差异,两种花粉发育的水平相当。

表1 花粉形态观察结果
2.1.2 花粉活力 在光照、黑暗和光照/黑暗3种环境下观察花粉的萌发率(详见表2)。两性植株花粉在3种条件下的萌发率分别为23.23±8.17%、34.30±11.47%、26.13±9.95%,黑暗中花粉的萌发率较高;雄株花粉在3种条件下萌发率没有差异,分别为25.50±14.45%、26.73±14.64%、25.60±16.10%。在光照和光照/黑暗条件下两性植株花粉的萌发率与同一条件下的雄株花粉没有差异,但在黑暗条件下,两性植株花粉的萌发率明显高于雄株花粉。因此,两性植株花粉的活力在光照和光照/黑暗条件下与雄株花粉在同一水平,在黑暗条件下则高于雄株花粉。

表2 两性植株与雄株花粉萌发率比较 %
2.1.3 雄性繁殖能力 用两性植株花粉给雌株授粉,共授粉51个花序,1785朵花,采收到1563个果实;用雄株花粉给雌株授粉,共授粉51个花序,1954朵花,采收到1767个果实(详见表3)。两性植株花粉授粉的得果率平均为86.69±13.90%,雄株花粉授粉的得果率平均为87.72±15.78%,两种花粉授粉后的得果率没有差异。
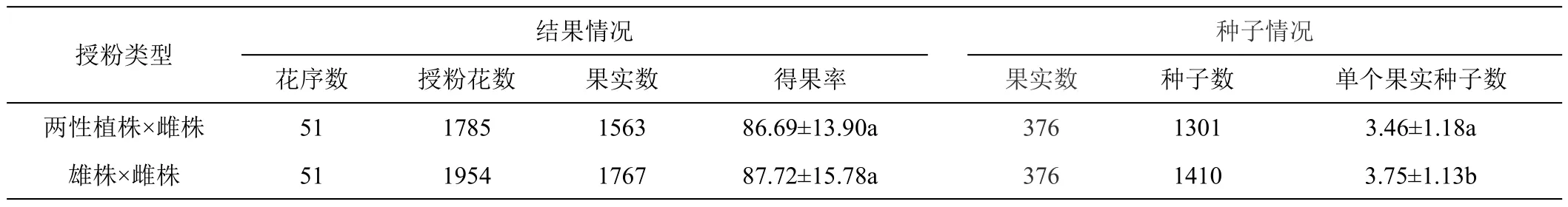
表3 两性植株雄性繁殖能力评估
在种子数量上,对376个两性植株雄花授粉和雄株花粉授粉所得果实的种子进行计数,两性植株授粉共得到1301个种子,单果种子数平均为3.46±1.18;雄株花粉授粉共得到1410个种子,单果种子数平均为3.75±1.13。两性植株花粉所育果实中平均种子数比雄性植株花粉所育果实中平均种子数少0.28,明显少于雄株。
上述结果说明,两性植株花粉在花粉的形态和得果率方面与雄株花粉没有差异,但在所育果实中种子数明显少于雄株花粉,两性植株花粉在产生后代的数量上略差。总体而言,两性植株能够成功执行雄性功能,具有繁殖能力,但在产生后代数量的能力上略差。
2.2 两性植株的雌性功能研究
2.2.1 柱头形态 通过扫描电镜对不同开花天数的21个柱头进行了观察,其中两性植株柱头11个,雌株柱头10个。观察发现,两种柱头表面形态相似(如图2),柱头呈近五边形,表面由乳突细胞组成,花蕾期和开花初期可观察到两类乳突细胞,一类细胞饱满,顶端不平整,另一类细胞不饱满,顶端向下凹陷,表面具有浸出物。随着开花天数的增加,两性植株和雌株的柱头整体形态不发生改变,乳突细胞随时间形态有所改变,饱满的乳突细胞顶端逐渐凹陷,点状凸起和表面浸出物逐渐增多。

图2 两性植株和雌株柱头的显微结构
两性植株柱头和雌株柱头形态基本一致,但雌株柱头乳突细胞从饱满到凹陷的时间较快,在开花2天的柱头上基本无饱满乳突细胞存在,浸出物也多于两性植株柱头。乳突细胞形态的变化与功能是否存在关联以及花粉在两类乳突细胞上的萌发和生长是否存在差异需要进一步的观察。
2.2.2 雌性繁殖能力 用雄株花粉分别对两性植株和雌株的雌花进行人工授粉,分别授粉19个花序。两性植株共授粉762朵花,采收到697个果实;雌株共授粉680朵花,采收到636个果实(详见表4)。两性植株得果率平均为91.30±8.51%,雌株得果率平均为91.56±7.98%,二者不存在差异。笔者还对两性植株和雌株自然受粉花序的结果状况进行了观察,分别观察了19个花序,两性植株的得果率平均为95.69±3.80%,雌株的得果率平均为87.30±17.61%,二者之间同样不存在差异。

表4 两性植株雌性繁殖能力评估
在种子数量上,对人工授粉的182个果实的种子进行计数,两性植株共得到729个种子,单果种子数平均为4.01±0.91;雌株共得到685个种子,单果种子数平均为3.68±1.16,两性植株显著高于雌株。分别对自然受粉两性植株和雌株的233个果实的种子进行计数,两性植株共得到924个种子,单果种子数平均为3.97±1.07;雌株共得到862个种子,单果种子数平均为3.70±1.21,两性植株同样显著高于雌株。
在人工授粉条件下,花粉充盈,通常而言后代应多于或等于自然受粉。在本实验中,雌株在人工授粉和自然受粉条件下,得果率和单果种子数均无显著差异。两性植株在人工授粉条件下的得果率显著小于自然受粉,单果种子数没有显著差异。差异的产生可能与光照有光,相对自然受粉,人工授粉的花序位于植株的下端,不如自然受粉的花序接受阳光充足,或许导致果实的发育不如上端的花序。
由上可知,两性植株柱头的形态和发育过程与雌株基本一致,在人工授粉和自然受粉2种条件下,两性植株能够产生与雌株相差无几的果实,且单果种子数高于雌株,表明两性植株柱头的可授性可能要强于雌株。因此,两性植株雌性不仅能够成功繁殖后代,而且在产生后代数量的能力上要高于雌株。
2.3 两性植株自交繁殖能力研究
为了考察两性植株的自交繁殖能力,笔者对两性植株进行自株授粉和异株授粉(详见表5),分别为19个花序,自株授粉523朵花,采集到475个果实;异株授粉762朵花,采集到697个果实。两性植株自交得果率平均为88.01±11.86%,异交得果率平均为91.30±8.51%,二者没有显著差异。
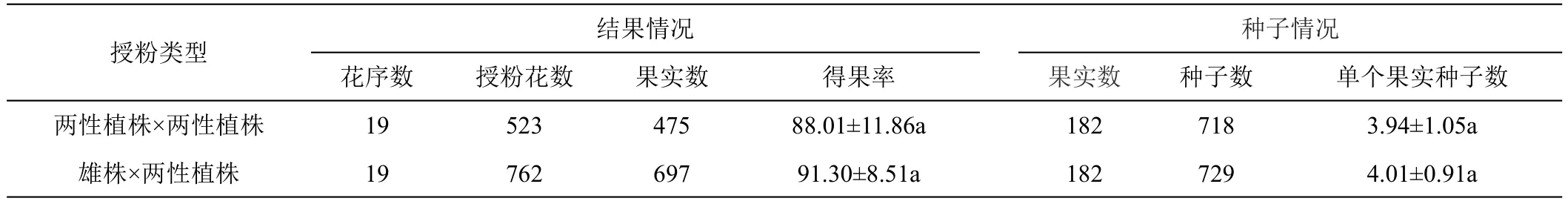
表5 两性植株自交繁殖能力评估
在种子数量上,分别对182个果实的种子进行计数,两性植株自交共得到718个种子,单果种子数平均为3.94±1.05;异交共得到729个种子,单果种子数平均为4.01±0.91,二者之间没有显著差异。
由上可知,两性植株自交能够成功繁殖后代,与异交相比,两者在得果率和单果种子数上并没有差异,两性植株自交繁殖能力与异交处于同一水平,不存在自交不亲和现象。
3 讨论
根据笔者的研究结果,两性黄檗植株具有不低于正常植株的繁殖能力,在自然条件下可以成功繁殖后代。这也意味着两性植株单株可以产生很多果实,与胡杨等的两性植株不同[1-3]。结合3年观察结果,两性植株性别表达不发生变化,可以推断出两性黄檗植株能够稳定的存在,并具有重要的生存意义。两性黄檗植株雄性和雌性均具有繁殖能力,即雄性和雌性均具有繁殖意义,为黄檗繁殖系统的相关研究提供了基础,也为其性别形成与分化研究提供了方向,但黄檗性别机制还需要进一步研究。两性植株可以自交繁殖,单株即可完成生殖,保障物种的繁衍,可以在一定程度上克服野生黄檗种群个体分散、零星分布格局导致的传粉困难,有利于黄檗资源的恢复和群落生物多样性水平的维护。但野生黄檗中是否存在两性黄檗、个体数量如何还有待于系统的野外调查。
在雌雄异株种群中出现少量的雌雄同株植株时,该居群称为“亚雌雄异株”[29]。有学者指出亚雌雄异株居群的出现只是植物进化发育过程中的“意外”,因为植物不一定能够精确的控制性别表达,在遭遇某些复杂的环境后可能产生一些罕见或是不规律的个体,并无意义[30]。也有学者不同意上述观点,认为亚雌雄异株种群的出现是植物为了适应非生物环境而发生的改变[31-33]。植物所处的环境并非一成不变,大量研究发现,当所处环境不利于植物生长时植株更偏向雄性,当环境利于植物生长时植株更偏向于雌性[34],同时拥有雄性和雌性的两性植株能够更好地适应不同的环境[35]。黄檗的两性植株同时拥有两种性功能,能够在低密度的环境下维持生存和繁衍,其适应环境能力可能高于雌雄异株植株。有学者指出成功的繁殖依赖于植株良好的适应性[36],但黄檗两性植株的出现是由于“意外”,或是对不利环境的主动应对还有待进一步的研究。
4 结论
两性植株的雄性和雌性繁殖器官较之雄株和雌株,在形态和发育上基本没有差异。在繁殖能力方面,两性植株的雄性能够正常繁殖,但能力略低于雄株;两性植株的雌性也能够正常繁殖,且能力略高于雌株。总体而言,两性植株的雄性和雌性都可以成功的繁殖后代,繁殖能力较之雄株和雌株虽有一定的差异,但基本处于同一水平。两性植株也能够通过自交成功繁殖后代,其繁殖能力与异交相比处于同一水平上,不存在自交不亲和现象,单株即可完成繁殖任务。为以后黄檗乃至植物性别相关研究提供了基础,也为黄檗野生资源保护带来了希望,但黄檗性别形成与分化机制以及两性黄檗植株在野外情况还需要进一步的研究和调查。
猜你喜欢
安徽农业科学(2024年5期)2024-04-20 11:38:27
西北园艺·果树(2024年1期)2024-04-01 09:10:14
蜜蜂杂志(2022年8期)2022-09-14 14:29:36
传奇·传记文学选刊(2020年12期)2020-12-23 06:56:35
园林科技(2020年1期)2020-01-18 05:13:00
现代农业科技(2018年17期)2018-10-13 12:31:24
中国林副特产(2013年4期)2013-08-07 10:15:48
现代农业科技(2009年7期)2009-07-29 07:11:32
现代农业科技(2009年20期)2009-03-20 05:27:10
