
高原鼢鼠震动信号发生部位及室内信号特征分析
2021-05-27 03:09董克池侍俊杰周建伟花立民
草原与草坪 2021年1期
董克池,侍俊杰,周建伟,花立民
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心/国家林业和草原局高寒草地鼠害防控工程技术研究中心,甘肃 兰州 730070)
动物通过不同的感官及通信方式实现种内个体的识别定位、求偶竞争、逃避天敌以及觅食等行为[1-3]。影响动物通信方式的因素很多,如动物的形态特征、生理特点、行为特性以及动物所处环境的特征[4]。这些因素决定着动物在通信过程中使用什么样的信号能够最有效地传递信息。动物通信根据传播的方式分为化学通信、声音通信、视觉通信、电通信和震动通信(seismic communication)[5],其中震动通信在最近二十年才被发现并受到动物学家们的高度重视。目前已发现,从小型的白蚁、狼蛛等到大型的脊椎动物,如大象,都可以通过介质(如地面、植物的叶子或茎、水面、蛛网等)产生震动波(substrate-borne vibration)并传递相关信息[1,4-6]。
对于栖息于地下洞道的啮齿动物,由于其视觉受地下洞道黑暗、低氧等环境的严格限制,视觉系统严重退化[7]。在独立的地下洞道系统中,视觉和嗅觉等受到隔离土壤的限制无法实现种内信息交流。声音虽然可作为种内个体之间信息传递的渠道,但是,在地下独立的洞道系统中声波的传递会被极大地削弱[8]。研究发现,营地下生活的滨鼠科(Bathyergidae)和鼹形鼠科(Spalacidae)的地下啮齿动物通过产生震动信号来完成种内信息的传递。目前已报道有12种啮齿类动物利用不同介质传递震动波作为信息传递的一种方式[9]。但是,地下啮齿动物因个体间的差异使其震动信号发生部位和信号特征各有不同。滨鼠(Fukomysdamarensis)利用后肢交替击打洞壁产生震动信号,用于吸引邻近洞穴中的异性伴侣[10];盲鼹鼠(Spalaxehrenbergi)繁殖其间用头部击打洞壁顶端产生震动信号保护其领地[11],同时利用自身产生的震动波通过障碍物反射来确定障碍物的大小及尺寸[12]。岬鼠(Georychuscapensis)通过后肢产生震动来传递性别信息以求偶[13]。
高原鼢鼠(Mospalaxbaileyi)属于啮齿目鼹形鼠科鼢鼠亚科(Rodentia:Myospalacinae),是青藏高原高寒草甸区特有的地下独居性啮齿类动物[14]。在适应地下环境的过程中,高原鼢鼠演化出许多与地下环境相适应的形态和行为特征,如极度变形和骨骼特化的前肢,严重退化的视觉等[15],加之雌雄个体独居的洞道环境限制其种内听觉和嗅觉通信。李金刚等[16]报道了甘肃鼢鼠(Eospalaxcansus)利用鼻吻部连续敲击洞道顶部而产生有节奏的震动信号。高原鼢鼠是否存在震动通信行为目前报道很少。本研究通过室内模拟高原鼢鼠栖息洞道,观察高原鼢鼠是否会产生震动行为,如果发生,发生部位和信号特征是什么。本研究结果将为明晰高原鼢鼠震动通信机理奠定基础。
1 材料和方法
1.1 高原鼢鼠捕获
高原鼢鼠捕获点位于甘肃省天祝县抓喜秀龙(N 37°11′,E 102°31 10′,海拔2 958 m)。草地类型为高寒草甸。利用鼢鼠无损伤活捕笼[17],于2018年6月捕获高原鼢鼠雌雄成体各10只,平均体重分别为152 g和220 g,带回甘肃农业大学养鼠室单笼饲养。笼上罩有遮光板以模拟地下环境。室内温度控制在23~25℃。每日早晚饲以胡萝卜、土豆、卷心菜等。待高原鼢鼠个体适应7 d后,开始室内观察试验。
1.2 研究方法
采用透明PVC管(长=120 cm;内径=8 cm)模拟高原鼢鼠洞道。管道两端各连通一个正方形亚克力箱体(50 cm×50 cm×50 cm)。管道中间用透明塑料板分隔,避免两鼠相互接触。利用照相机(Nikon D7100)录制其震动行为,以查明高原鼢鼠震动通信信号发生的身体部位。将震动波检波器(LGT-20,4.5 Hz) 每隔20 cm布置于塑料管上方,共计4个。试验时,单独或同时放入不同性别高原鼢鼠个体,然后由检波器和BL-420生物机能实验系统(采样频率为1 000 Hz)收集和记录。单只个体连续观测记录12 h。
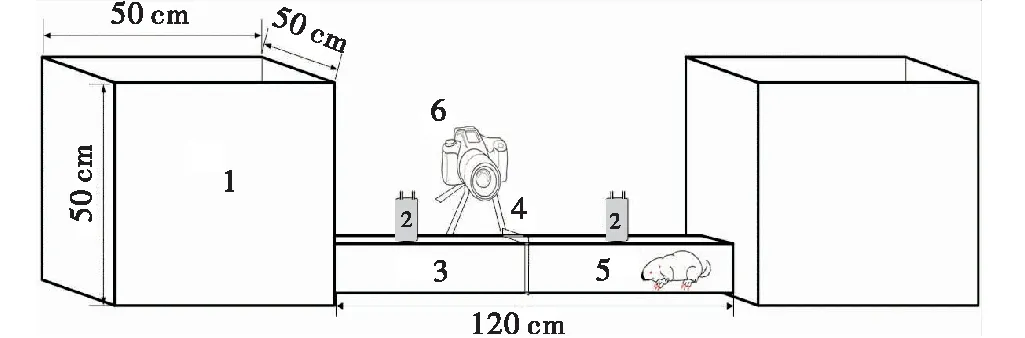
图1 高原鼢鼠震动通信信号发生部位观察示意图Fig.1 Schematic diagram for observing the seismic communication signal of plateau zokor in laboratory注:1:亚克力箱;2:检波器;3:模拟通道;4:隔板;5:实验鼢鼠;6:照相机
1.3 数据统计与分析
记录并分析高原鼢鼠雌性个体震动信号脉冲间距离、每个敲击回合的脉冲数量及持续时间、两次脉冲组之间的距离。采用独立样本t检验(Independent sample t-test)比较不同性别鼢鼠放入箱体后产生震动信号反应时间之间差异,同时对数据进行方差同质性检验,对方差不齐的数据则采用曼-惠特尼U检验(Mann-Whitney U-test)进行比较。震动信号能量频域由Matlab 2014a软件进行快速傅里叶变换(FFT)获得。利用独立样本t检验比较雌雄个体间信号特征差异,对于方差不齐的数据,同样用Mann-Whitney U-test进行比较。上述数据均采用SPSS 17.0软件统计分析完成。
2 结果与分析
2.1 震动发生部位及信号特征
通过录像数据发现,高原鼢鼠通过鼻吻部撞击洞道侧壁产生有节奏的震动信号。在单独观测的10只个体中,有5只雌性和3只雄性均利用鼻吻部敲击PVC管壁产生震动信号。敲击行为发生在单只个体刚放入箱体内不久,敲击行为雄性在(13.27±8.26)s后产生,而雌性则在(71.54±13.22)s后产生(表1)。高原鼢鼠一次完整的敲击震动由数个敲击回合(bout)组成,一个敲击回合由数个连续脉冲组成。1只雄性高原鼢鼠个体产生的震动信号由3个敲击回合构成,每个回合的脉冲数分别为10、10和8(图2)。1只雄性高原鼢鼠产生的震动信号由6个敲击回合构成(表3)。有时也会产生1个孤立的敲击信号。这种敲击行为可能是高原鼢鼠对陌生环境的警觉。随着时间的推移,高原鼢鼠试验个体逐渐适应箱体环境后,未观察到类似的敲击行为。但在箱体两端同时放入不同性别的个体后,也只观察到单只个体的敲击行为。随着记录时间的增加,实验中并未观察到2只个体同时或前后相互呼应的敲击信号。

表1 单只个体或雌雄同时放入箱体后产生的震动信号反应时间
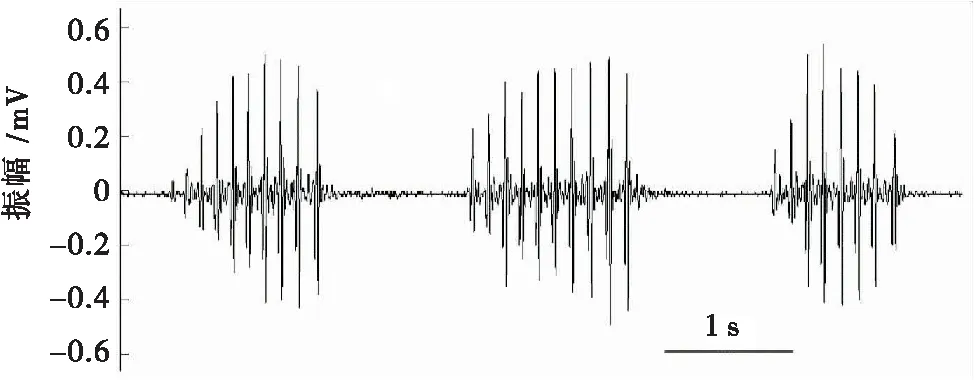
图2 雄性高原鼢鼠震动信号Fig.2 Seismic signals of male plateau zokor (contains 3 bouts)
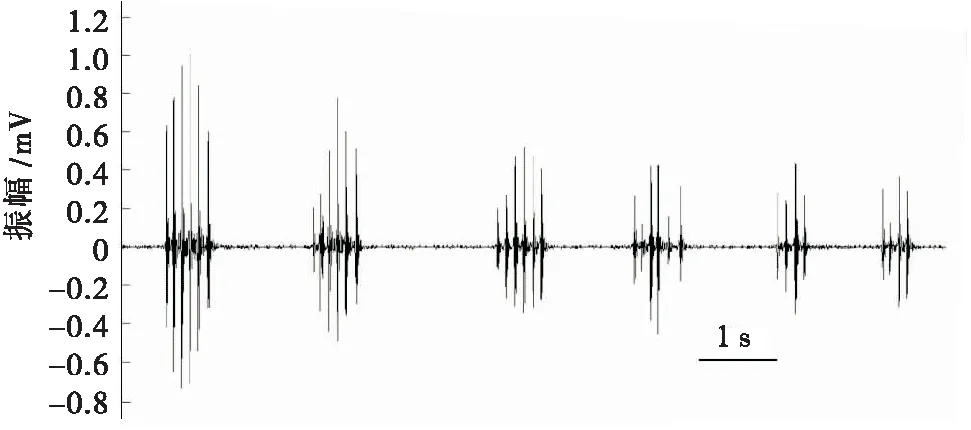
图3 雌性高原鼢鼠震动信号Fig.3 Seismic signals of female plateau zokor (contains 6 bouts)
就单个敲击回合而言,雄性一个敲击回合由2~17个脉冲(平均为6.67±2.63)组成,雌性一个敲击回合由2~14个脉冲(平均为6.04±2.35)组成,雄性显著高于雌性(P<0.05)(表2)。相应的雄性个体单回合敲击持续时间也都显著高于雌性个体(P=0.004)。但是相邻敲击回合时间间隔差异不显著(t=0.485,P=0.628),雌性略高于雄性。雄性单回合敲击信号中相邻2个脉冲时间间隔[(0.13±0.01)s]显著高于雌性[(0.12±0.01)s](t=2.403,P=0.016),说明雄性个体有着更快的敲击频率。
2.2 高原鼢鼠震动信号能量特征比较
雌性个体震动信号能量频率为19.13~120.57 Hz(图4A1),雄性个体总体分布范围为19.55~138.92 Hz(图4A2)。雄性个体敲击震动信号的振幅显著大于雌性个体(图5A1),这可能与雌雄个体体重差异有关。雌性敲击震动波主峰频率介于39.25~71.79 Hz(平均54.28±1.12 Hz),雄性敲击震动波主峰频率集中在38.42~60.94 Hz(平均为(49.16±0.97)Hz)(图5A2)。雌雄个体间震动波主峰能量频率差异显著(P<0.001),但峰值能量密度差异不显著(P=0.923)(图5A3)。
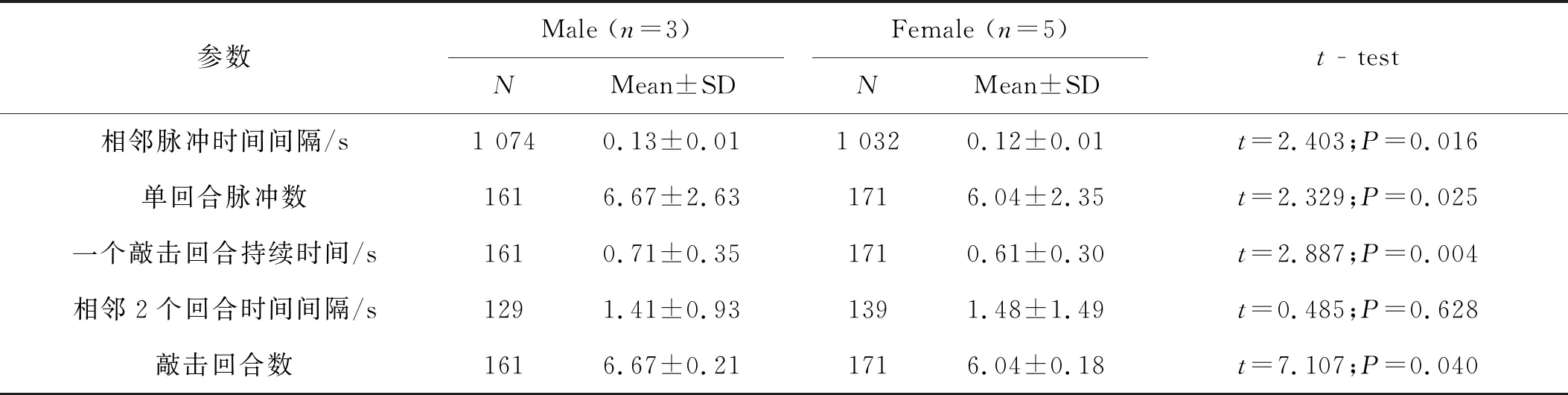
表2 高原鼢鼠鼻吻部敲击信号特征
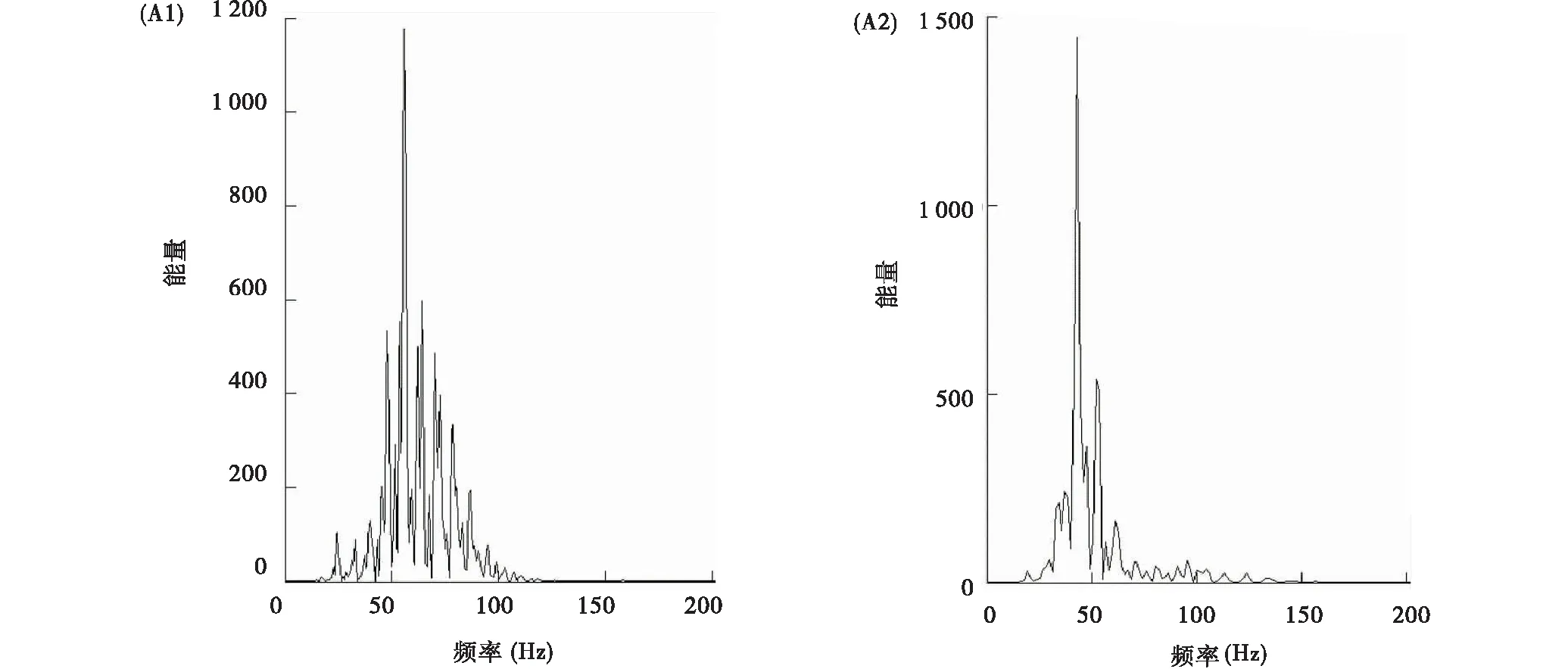
图4 高原鼢鼠单个震动信号能量谱Fig.4 Energy spectrum of one bout of seismic signal of plateau zokor
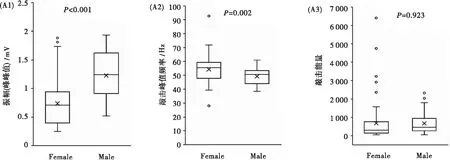
图5 不同性别高原鼢鼠震动信号能量比较Fig.5 Comparison of seismic signal energy between male and female
3 讨论
本研究结果表明,高原鼢鼠能够通过吻端敲击洞道侧壁产生震动波。该震动波一般都会形成数个连续的脉冲组合,每一个脉冲组信号由2~17个脉冲组成。目前研究已发现,只有鼹形鼠科的物种产生震动信号的方式是通过头部特定部位击打洞壁,如盲鼹鼠(S.ehrenbergi)[18],甘肃鼢鼠(E.cansus)等[16]。而滨鼠科啮齿动物是通过后足击打洞道下壁产生震动信号,如岬鼠(G.scapensis)[10]。本试验中高原鼢鼠刚放入箱体中就利用吻端敲击箱体侧壁,这可能是对陌生环境的警觉而产生敲击信号,敲击的力度由弱变强。在模拟的人工管道中雌雄个体相互接近并未发生头部敲击的行为,这与盲鼹鼠雌雄个体相互敲击响应对方的方式不同[18]。
高原鼢鼠雌雄个体吻端敲击产生震动信号能量的主峰频率分别介于39.25~71.79 Hz和38.42~60.94 Hz,总体上属于低频震动波。甘肃鼢鼠雌雄个体产生的震动信号能量主峰频率在100 Hz左右[16],高于高原鼢鼠的震动信号能量主峰频率。地下啮齿动物在长期的地下适应过程中,其听觉系统从形态到功能已出现明显的适应性进化,如耳廓退化,听觉趋于低频。Rado等[11]首次在野外记录到盲鼹鼠利用头部撞击洞道顶部产生震动信号,其大部分能量在集中在100~300 Hz。
岬鼠属(Georychus)动物利用声音产生震动信号,震动在相邻距离在3~4 m的洞道中可以被探测到,而声音信号的振幅在传递到1 m距离的时候会衰减到背景噪音水平[12]。因为在蜿蜒曲折、空气不流通的洞穴中,声音信号传递受到极大限制,特别是在风大或其他嘈杂的环境中。但是震动信号非常适合作为用于地下孤立洞道系统之间的长距离通信,低频的震动波在这种环境中能更有效的传播[19]。因此,岬鼠属动物震动信号可以在地下较远距离被感知。
高原鼢鼠作为青藏高原生态学研究的热点动物之一,对草地生态系统的植物、土壤等有着复杂而深远的影响[20-21]。高原鼢鼠除繁殖期外,常年独居于地下黑暗洞道环境,视觉系统极度退化。独特的洞道环境虽为高原鼢鼠生存提供了天然屏障,但在其繁殖其间,通过何种方式进行信息交流,如何在复杂的地下洞道系统中寻找最佳的通道接近配偶,如何有效地维护领地及个体识别,如何有效地获取最佳采食路线。这些问题都与其独特的震动通信有关。本研究采用室内模拟试验发现其存在震动敲击行为,而且雌雄个体敲击信号特征存在明显差异,这对高原鼢鼠震动通信的进一步研究及高原鼢鼠鼠害防治具有指导意义。研究高原鼢鼠在野外自然环境中产生震动信号的特征并解读其生物学意义,对于了解其种内通信机理具有重要意义,这也是今后类似研究的发展方向。
4 结论
高原鼢鼠利用鼻吻部敲击洞壁产生震动信号。雌雄震动信号都是由数个连续的敲击回合构成一个完整的系列,但是雄性敲击回合数显著高于雌性。雄性个体震动信号单回合的敲击时长及脉冲次数表现出极显著差异,但相邻回合的时间间隔差异不显著,这可能是种内个体间信号固有的特征。此外,鉴于雄性体重的优势,其信号的振幅显著大于雌性,但雄性信号的频率显著低于雌性。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
世界科学技术-中医药现代化(2021年5期)2021-11-05
山西林业(2021年2期)2021-07-21
兽医导刊(2020年3期)2020-12-30
防护林科技(2019年7期)2019-03-25
现代园艺(2018年2期)2018-03-15
湖南中医药大学学报(2015年1期)2016-01-06
Coco薇(2015年12期)2015-12-10
