
华山松第一代无性系种子园种源研究
2021-05-25 05:21:10董云祥罗正平潘涌智伍健榕许玉兰吴利华
西南农业学报 2021年3期
冯 峻,董云祥,罗正平,潘涌智,伍健榕,许玉兰,吴利华
(1.云南省林木种苗工作总站,云南 昆明 650215;2.楚雄市林业和草原局,云南 楚雄 675000;3.西南林业大学 云南省森林灾害预警与控制重点实验室,云南 昆明 650224;4.西南林业大学 西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224;5.西南林业大学 地理与生态旅游学院,云南 昆明 650224)
【研究意义】华山松(PinusarmandiFranch.)属松科(Pinaceae)松属(Pinus)乔木,是中国特有的五针松树种[1]。云南是华山松主要分布区,林分面积和蓄积量居云南乔木林树种的前10位[2],具有栽培历史悠久、分布广泛、适应性强等特点,也是云南特有造林、用材、用脂、栲胶和种仁食用树种,在林业建设和生态文明建设中占有显著地位。选择适宜和优良的种源,是林木良种选育的基础,是提高林木产量和质量的重要途径,1984年原云南省林业厅以宜良、楚雄、南华、会泽、腾冲、巍山等县(市)的6个种源为选择对象,选出表型优良的129株优树,在楚雄市紫溪山林场营建华山松第一代无性系种子园,种子园1998年开始结实,2002年开始进入盛产期,年均可产3 t左右的良种。林木育种研究的对象是群体[3],优良种源是第一代种子园遗传改良研究中重要的育种群体。【前人研究进展】华山松对生境变化反应比较敏感,受生境差异、自然选择等因素影响,种内遗传分化明显,产生极为丰富的地理变异种源[4];华山松又是一个多层次遗传变异树种,在遗传改良过程中存在多层次的遗传改良可供选择和利用[4],除种源间、种源内的遗传变异外,还有家系(无性系)、单株的遗传变异。不同种源的华山松在相同环境下,各种源在形态、生长量、适应性、遗传稳定性和抗冻性等方面均存在显著的遗传变异和差异[5-9]。【本研究切入点】研究和分析第一代华山松种子园内各种源表型变异规律、差异特点,系统地揭示不同种源对环境的适应情况,进而为第一代种子园再选择和遗传改良提供优良种质材料和理论依据。【拟解决的关键问题】以华山松第一代无性系种子园为研究对象,对华山松不同种源的树高、胸径、冠幅、材积和结实球果等表型性状进行调查、分析和研究,以期揭示第一代华山松种子园表型性状在种源间、种源内的变异规律、差异特点、相关性和优良种源的评定等,为优良繁殖材料的再选择、杂交育种亲本选配、遗传测定林建设和高世代种子园建设等提供理论依据和技术支持,也可为大面积华山松营林提供优良材料。
1 材料与方法
1.1 试验地概况
楚雄市紫溪山林场华山松第一代无性系种子园位于楚雄市紫溪山自然保护区内,距楚雄市西南20 km,地处北纬24°58′58″~25°04′02″,东经101°22′29″~101°26′07″,海拔2200~2400 m,地势平缓,光照充足,总面积51 hm2,分为良种生产区、子代测定区和实验示范区等;该地属北亚热带温凉湿润气候,年平均气温12 ℃,平均最高气温20 ℃,最低气温-6 ℃左右,年降雨量1000 mm,平均相对湿度80 %~85 %。土壤为暗红壤,土层深厚,有机质含量高,分解较快,质地疏松,吸水性、透气性良好,土壤中氮、磷、钾等有效成分含量较高。
1.2 试验材料及调查
以楚雄、腾冲、南华、会泽、巍山和宜良等县市6个种源(表1)为研究对象。2018年对6个种源的树高、胸径、冠幅等生长指标进行全面调查,采用测高仪测定华山松树高,用围径尺测量胸径,用皮尺测定冠幅;调查统计6个种源的结实量。
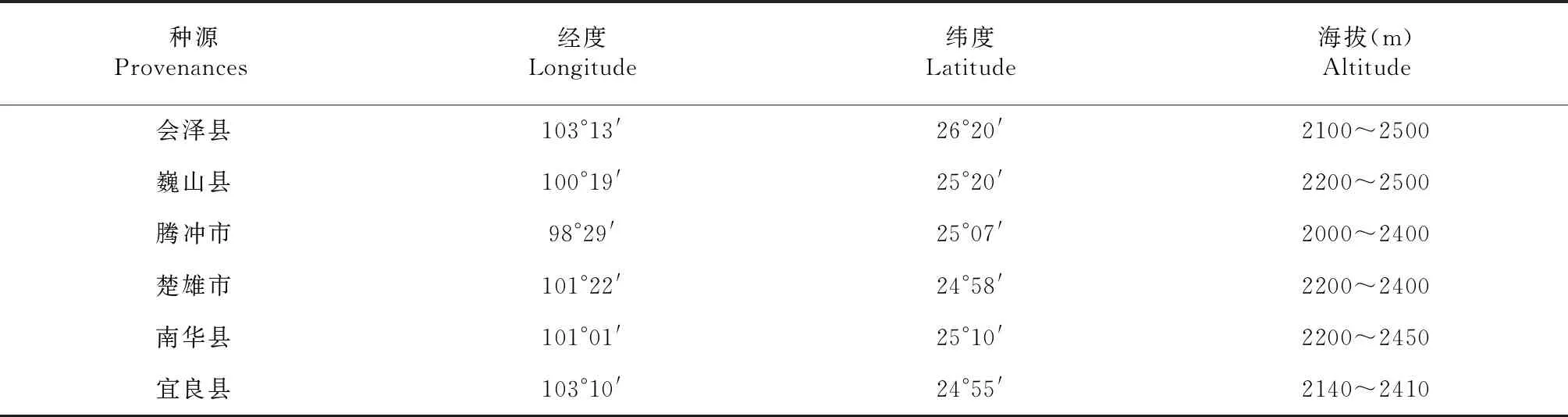
表1 华山松第一代无性系种子园各种源原产地概况Table 1 The general information of origin places for provenances in the first generation clonal seed orchard of Pinus armandi Franch.
1.3 分析方法
采用Excel和SPSS.18对数据进行处理、分析。
华山松材积采用V=aDbHc公式计算,其中V为单株材积,D为胸径,H为树高,a=0.000073535016,b=2.0015694,c=0.7888835。

重复力计算公式[11]:R=1-1/F,式中:R为某一性状重复力,F为方差分析中的F值。
2 结果与分析
2.1 不同华山松种源性状生长和变异分析
2.1.1 不同华山松种源性状生长分析 由表2可知,6个华山松种源平均树高为11.09 m,幅度在10.87~11.95 m,树高最大的是南华种源11.95 m,最小的是巍山种源10.87 m;平均胸径为21.51 cm,幅度在20.74~23.41 cm,胸径最大的是会泽种源23.41 cm,最小的是巍山种源20.74 cm;平均材积为0.257 m3,幅度在0.237~0.307 m3,材积最大的是会泽种源0.307 m3,最小的是巍山种源0.237 m3;平均冠幅为6.97 m,幅度在6.68~7.474 m,冠幅最大的是南华种源7.474 m,最小的是腾冲种源6.68 m;平均结实球果数为6.98个,幅度在4.37~10.83个,结实球果数最大的是南华种源10.83个,最小的是宜良种源4.37个;华山松种子园林分保存情况良好,有较好的生长表现,在种子园遗传改良过程中有着重要的基础作用。

表2 不同华山松种源生长性状的均值、标准差和多重比较Table 2 The mean values,standard deviations and multiple comparisons of growth traits for different provenances of Pinus armandi Franch.
2.1.2 不同华山松种源性状变异分析 由表3可知,变异系数可反映华山松第一代无性系种子园种源间与种源内性状值的离散情况,变异系数小则性状稳定,变异系数大则离散程度大。6个华山松种源树高平均变异系数为22.98 %,幅度在21.01 %~23.49 %,变异最大的是巍山种源23.49 %,最小的是腾冲种源21.01 %。胸径平均变异系数为25.57 %,幅度在20.48 %~27.64 %,变异最大的是会泽种源27.64 %,最小的是腾冲种源20.48 %。材积平均变异系数为60.76 %,幅度在53.93 %~62.07 %,变异最大的是巍山种源62.07 %,最小的是腾冲种源53.93 %。冠幅平均变异系数为22.29 %,幅度在16.16 %~24.29 %,变异最大的是巍山种源24.29 %,最小的是腾冲种源16.16 %。结实球果平均变异系数为130.45 %,幅度在96.67 %~170.47 %,变异最大的是宜良种源170.47 %,最小的是南华种源96.67 %;6个华山松种源性状平均变异系数中宜良种源变异系数最大58.64 %,南华种源变异系数最小42.92 %。结果表明南华种源性状稳定,华山松种子园种源间、种源内均存在广泛和丰富的变异,有很大的选择潜力,为种子园开展选育提供有利条件。
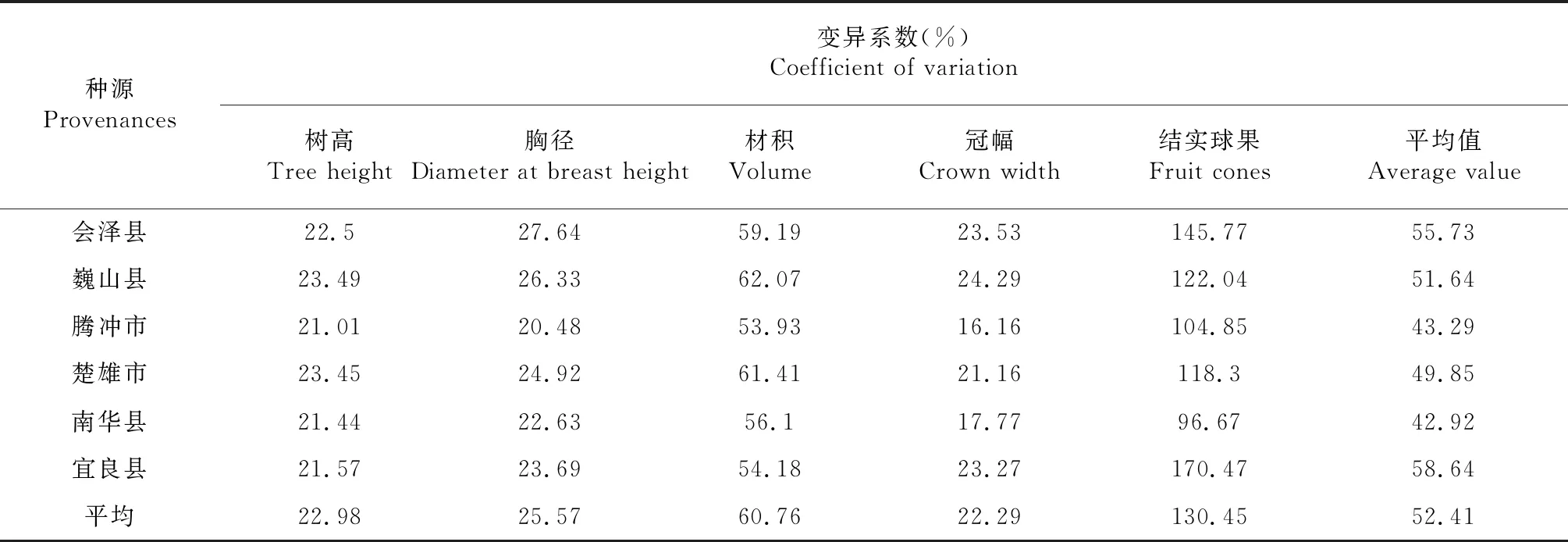
表3 不同华山松种源性状的变异情况Table 3 The variations of traits for different provenances of Pinus armandi Franch.
2.2 不同华山松种源性状差异分析
2.2.1 不同华山松种源性状方差分析 华山松第一代无性系种子园种源间和种源内的方差分析结果见表4。种源间的F值为2.42~8.26,经显著性检验显示,除树高在种源间的差异呈显著水平(P<0.05)外,其他4个性状的差异均呈极显著水平(P<0.01),生长差异较大;各性状的重复力为58.74 %~87.90 %,其中:胸径、材积和结实球果均达到80 %以上。种源内的F值为2.30~2.78,显著性检验显示,5个性状的差异均呈极显著水平(P<0.01),生长差异较大;各性状的重复力为56.44 %~63.99 %,其中:胸径、冠幅和结实球果均达到60 %以上。

表4 不同华山松种源性状方差分析Table 4 Analysis of variances of traits for different provenances of Pinus armandi Franch.
华山松种源间存在显著差异,但不代表各种源两两之间也存在显著差异,因此需要对各性状进行多重比较。经多重比较由表2可知,会泽与南华,巍山与南华、宜良,楚雄与南华种源之间树高生长差异达显著水平。会泽与巍山、楚雄,楚雄与会泽种源之间胸径生长差异达极显著水平;会泽与腾冲、宜良,巍山与楚雄、南华种源之间胸径生长差异达显著水平。会泽与腾冲,巍山与楚雄、南华、宜良,腾冲与楚雄、南华、宜良种源之间冠幅生长差异达显著水平。会泽与腾冲、楚雄,南华与腾冲、楚雄、宜良种源之间结实球果差异达极显著水平;会泽与巍山、宜良,巍山与腾冲、楚雄、南华、宜良种源之间结实球果差异达显著水平。会泽与巍山、楚雄种源之间材积生长差异达极显著水平;会泽与腾冲,巍山与南华种源之间材积生长差异达显著水平。
华山松种子园各性状在种源间和种源内均有差异,不同种源的华山松遗传分化显著,差异由遗传因素引起,而且受高等遗传力制约,有很大的选择潜力,通过选择可获得不同层次的遗传增益。
2.2.2 不同华山松种源间表型分化分析 通过方差分析将表型变异加以分解,分别计算种源间、种源内方差分量占总变异的百分比,结果见表5。种源间的方差分量占总变异的1.75 %,种源内的方差分量占总变异的13.56 %,误差占总变异的84.69 %;5个性状的表型分化系数变异为1.09 %~17.42 %,其中结实球果的表型分化最大17.42 %,树高的表型分化最小1.09 %。种源间的平均表型分化系数为10.89 %,表明华山松种源的表型性状变异主要来源于种源内。

表5 不同华山松种源各性状方差分量及种源间表型分化系数Table 5 The variance components of traits and the phenotypic differentiation coefficients between different provenances of Pinus armandi Franch.
2.3 不同华山松种源相关分析
为研究华山松种源各生长指标与原地产经、纬度之间的关系,以种源为控制量对6个种源的5个生长指标,原地产经、纬度进行相关分析(表6)。树高与纬度呈显著正相关;胸径与经度、纬度、树高呈显著正相关;冠幅与经度、纬度、树高呈极显著正相关;结实球果与经度、纬度呈显著正相关,与树高、胸径、冠幅呈极显著正相关;材积与经度、纬度、树高、胸径、冠幅、结实球果呈极显著正相关。
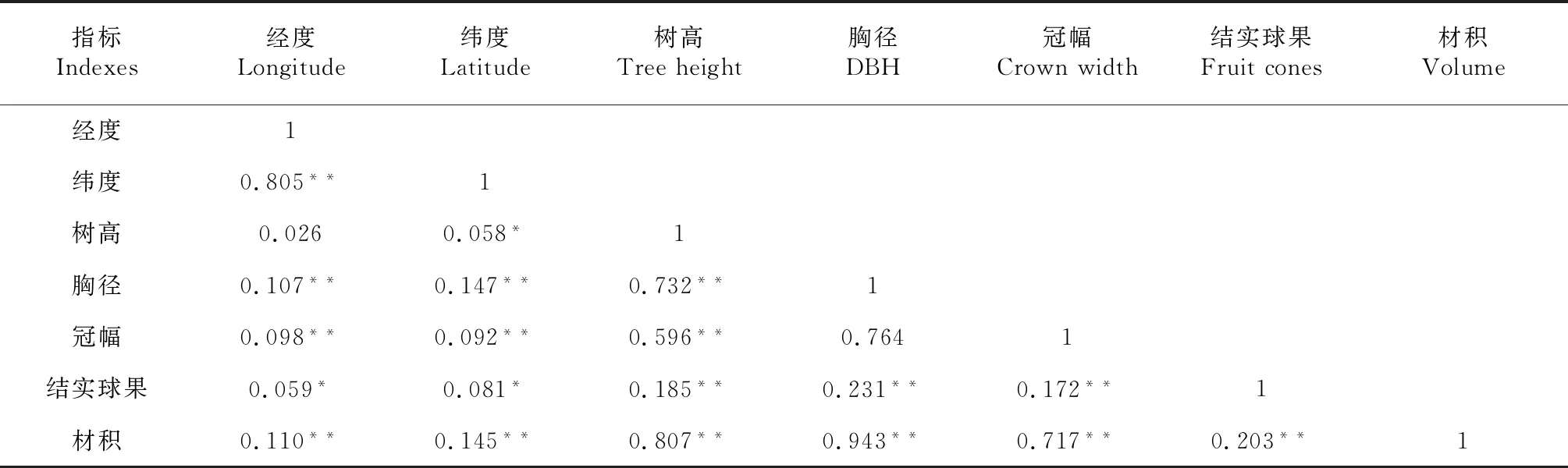
表6 各种源生长性状、经纬度相关性分析Table 6 The correlation analysis of growth traits and latitude,longitude for various provenances
2.4 种源综合评价
如表7所示,Pi值最小的是南华种源,其次是宜良种源、腾冲种源、会泽种源、楚雄种源、巍山种源。排名第1的南华种源各性状变异系数相对其它种源变化不大,表现最稳定,树高、胸径、冠幅、结实球果数和材积相比总体均值,分别增长7.71 %、4.67 %、7.23 %、55.09 %和13.5 %,其各性状预期遗传增益分别为4.79 %、3.95 %、5.63 %、5.63 %、48.95 %和11.22 %。种源的综合评定为进一步开展选择、杂交育种提供了数据和原始材料,也为各造林地区确定华山松生产力高、稳定性好的种源提供了科学依据,但优良种源还需进行遗传测定进一步确定。

表7 不同华山松种源选择结果Table 7 The selection results of different provenances of Pinus armandi Franch.
3 讨 论
3.1 关于种子园各种源的差异
华山松是雌雄同株异花授粉的植物,长期在种子园内进行自由授粉,6个华山松种源生长性状表现良好,证明表型选择是有效的。相同环境下,不同种源的不同性状在发育过程中产生不同程度的差异,形成广泛和多样的杂种群体;不同性状在种源间和种源内,均存在不同程度的差异,差异呈显著或极显著水平,种源间的差异大于种源内的差异,均受高等遗传力控制,再进行性状选择,可获得更高的遗传增益。表型选择建立的第一代无性系种子园各种源的不同性状分化明显,存在差异,表型优良不代表基因型优良,建设遗传测定林开展遗传测定,才能获得更大的增益。另一方面也表明从各种源内进一步选择优良无性系、优良单株,可获得更高的遗传增益。一些学者对杉木、日本落叶松、马尾松等种子园遗传改良时,在分析种源差异的基础上,进行无性系(家系)的遗传改良,取得更大的遗传增益,选出生产力高的群体[14-16]。因此,华山松第一代种子园种源间的差异是选择育种中重要的变异来源,在遗传改良时应考虑优良种源、无性系和单株的多层次选择。
3.2 关于种子园各种源的变异
遗传变异是群体生物逐渐适应环境和不断进化的产物[11],利用表型性状来研究林木的遗传变异是最有效的方法[17]。华山松第一代无性系种子园在有性繁殖和自然选择条件下,个体发育差异使生长和结实等性状产生与之相适应的变异,不同性状变异为22.98 %~130.45 %,变异系数大小顺序为:结实球果数>材积>胸径>树高>冠幅。种源间各性状的平均变异系数为52.41 %,说明种源内表型性状离散程度较高,其中:变异系数最大的是宜良种源58.64 %,变异系数最小的是南华种源42.92 %,变异系数越大,表型多样性程度越高,存在较高的遗传改良潜力。群体间的变异反映了地理与生殖隔离上的变异[18],是种源间分化的来源,也是种源内多样性关键组成部分;种源内表型多样性程度高于种源间表型多样性,种源内的变异是华山松种子园的主要变异来源。赵扬等[19]对华山松无性系种子园进行遗传多样性分析,也得出变异主要存在于种源内的结论。林木各层次的遗传变异量为种源>家系>单株[20],华山松种源丰富的变异为种子园进一步开展多层次优良繁殖材料选择和杂交亲本选配提供基础,也为开展优良华山松种质资源保护提供参考。
不同树种的地理变异规律十分复杂[21],华山松是以纬向渐变方式表现其遗传地理性分化的树种[22],华山松生长与纬度关系密切呈极显著的负相关[23];而种子园建园亲本6个种源地的纬度间隔不大,现已是成熟林,属于云贵高原种源地理生态型[22],不同种源的华山松随着林龄增大,受环境效应的影响,产生与之相适应的地理变异,各种源的树高与原产地纬度呈显著正相关,胸径、材积、冠幅和结实球果与原产地纬度呈极显著的正相关,这可能与种源多年生长的适应性有关,吴海[24]对21年侧柏地理种源地理变异规律的研究也有相同的结果。这种渐变模式为制定种子调拨、异地用种和适应范围提供了依据。
3.3 关于种子园的选择和改良
林木性状间的遗传相关关系的高低,能反映出性状间的相关程度,可为林木遗传改良方案和策略提供理论参考[25]。华山松是兼用树种,各性状的相关性主要是由林木本身遗传特性决定,材积与树高、胸径、冠幅、结实球果呈极显著正相关关系,结实球果数与树高、胸径、材积、冠幅呈极显著正相关关系,过分强调生长性改良将可能导致结实量下降,故在进行华山松遗传改良时,应同时兼顾。
优良种源是林木遗传改良研究中重要的育种资源[26],进行多层次选择,可选出优良无性系和单株作为建立新一代育种群体的繁殖材料。林龄越大,选择结果越可靠,多性状联合选择更有利于保持遗传多样性[27],林木遗传改良的目的是提高林分的生长量,改良木材品质及增强林分稳定性[28]。在华山松第一代无性系种子园种源评定方法上,采用多性状综合评价方法,综合树高、胸径、冠幅、材积和结实球果等5个性状表现,对各种源进行综合评定,避免单一考虑某个性状选择优良种源的局限性。南华种源生长较为均匀,进一步选择优良无性系和单株将获得遗传增益,推广和使用这样的种源有更高的应用价值;宜良、腾冲、会泽、楚雄和巍山种源群体内有不同程度的变异和差异,应进一步选择优良无性系和单株,以提高遗传增益。
4 结 论
楚雄市紫溪山林场第一代华山松种子园生长和结实情况良好,遗传改良潜力大,各性状的重复力较高,受遗传控制程度较强,能为优良种源、无性系和单株的再选择提供基础材料;在选育和改良工作中,将种源间、种源内选择结合起来,充分利用种源内和种源间的变异,全面地收集基因资源,选配亲本开展种间和种内杂交;根据子代测定结果再选定优良种源、无性系和单株,作为高世代种子园建园材料。同时,选出适应环境及生存能力更强的华山松种质资源,以保持种子园较高的遗传多样性水平,实现种子园高产、优质、高效的目标。
致谢:在野外调查过程中得到西南林业大学2016级研究生董诗涛、2017级研究生赵勋、熊鑫鑫等同学的帮助,在数据处理过程中得到西南林业大学王辉老师的指导,在此表示衷心感谢。
猜你喜欢
科海故事博览(2022年28期)2022-11-25 03:09:48
南华大学学报(社会科学版)(2022年3期)2022-10-12 08:06:30
南华大学学报(社会科学版)(2022年2期)2022-08-03 05:35:42
南华大学学报(社会科学版)(2022年1期)2022-05-10 03:44:38
辽宁林业科技(2021年3期)2021-07-08 03:11:36
南华大学学报(社会科学版)(2021年2期)2021-06-03 01:10:44
中国森林病虫(2020年1期)2020-01-18 07:21:52
新闻传播(2018年15期)2018-09-18 03:20:12
广西林业科学(2016年1期)2016-03-20 05:32:59
广西林业科学(2016年1期)2016-03-20 05:32:58
