
百合Sorbonne 和Red Life 香气差异及其单萜合成酶结构分析
2021-05-24 01:52刘芝芝樊俊苗王银柱高建华
山西农业科学 2021年5期
刘芝芝,樊俊苗,王银柱,高建华,杜 方
(1.山西农业大学园艺学院,山西太谷030801;2.南京农业大学园艺学院,江苏南京210095;3.山西农业大学生命科学学院,山西太谷030801)
百合(Liliumspp.)是百合科(Liliaceae)百合属(Lilium)具有重要经济价值的球根花卉[1]。目前,百合属约有120 个种,还有很多人工杂交产生的新品种[2]。花香是百合重要的观赏性状之一。不同种类百合香味不同,如东方百合香味浓郁,而亚洲百合香气微弱[3]。香气改良是百合育种的重要目标之一。
百合主要释香部位是花被片,单萜类化合物是百合花香物质的主成分之一。从花被片中释放的萜类物质成分和含量因品种不同,差异较大[4-6]。萜类物质的合成受甲羟戊酸(mevalonic acid,MVA)和甲基赤藓糖醇磷酸(methylerythritol phosphate,MEP)途径多种酶的调控,其中,萜烯合酶(terpene synthase,TPS)是MVA 和MEP 途径中催化最后一步反应的酶。TPS基因在浓香型百合中的表达水平显著高于无香或淡香型中的表达水平[3-4,7-8]。王婷等[5]已从41 个百合品种(含Sorbonne)中鉴定出46 种香气成分,并对其进行香型分类[9];同时,樊俊苗[10]从浓香Sorbonne 中克隆出3 个单萜合成酶基因(LsoTPS-1、LsoTPS-2、LsoTPS-3),从无香Red Life中克隆出2 个基因(LrlTPS-1、LrlTPS-3),并揭示了其核酸的多态性。但是对Sorbonne 和Red Life 香气差异及其TPS 蛋白结构和功能未深入研究。
本研究通过气相色谱- 质谱联用技术对2 个百合品种挥发性物质进行定性和定量分析,旨在研究浓香与无香百合香气成分差异;同时对2 个品种中TPS 蛋白结构进行生物信息学分析,从而揭示二者单萜合成酶在蛋白水平上的差异,为之后TPS 蛋白功能验证奠定基础。
1 材料和方法
1.1 材料与仪器
1.1.1 材料 2016 年,Sorbonne 试验材料来自山西农业大学园艺站露地百合资源圃,在晴朗天气下采集生长健壮植株盛花期的花被片用于香气测定;2017 年,Sorbonne 来自山西省晋中市太谷县切花市场,在3 枝切花中各取1 朵正处于盛花期的花作为供试材料,重复3 次;2018 年,Sorbonne 来自山西农业大学园艺站温室内,选择正处于盛花期、长势一致且健壮植株3 株,在晴朗天气的8:00—10:00 每株各采1 朵花组成1 个样品,重复测定3 次。2020 年,Red Life 材料来自山西农业大学园艺站露地百合资源圃,供试材料选择与制样同2018 年Sorbonne。Sorbonne 和Red Life 的相关信息如表1 所示。
表1 Sorbonne 和Red Life 相关信息
1.1.2 仪器 Trace ISQ 气相色谱质谱联用仪(美国Thermo Fishe 公司);DB-5MS(30 m×0.25 mm×0.25 μm)色谱柱(美国ThermoFishe 公司);50/30 μm PDMS/DVB/CAR萃取头和进样器(美国Supelco公司)。
1.2 方法
1.2.1 香气采集及GC-MS 分析 称取花被片约1.5 g 置于20 mL 顶空固相微萃取专用顶空瓶中,利用顶空进样器提取香气成分并完成进样,对百合挥发性物质进行GC-MS 分析。
气相色谱条件:DB-5MS 毛细管色谱柱(30 m×0.25 mm×0.25 μm);流速1 mL/min,He 为载气;程序升温:起始柱温40 ℃,保持2 min,之后以5 ℃/min速率升温至270 ℃,持续6 min。
质谱条件:电离方式为EI;电离电压70 eV;质谱扫描范围45~600 amu;离子源温度280 ℃;进样口温度280 ℃。
利用Thermo Xcalibur 软件系统自带的NIST 标准库检索各组分,参考文献[3,5,9]进一步筛选,综合考虑最终确定Sorbonne 和Red Life 中的挥发性物质。利用峰面积归一化法,计算各组分相对含量。
1.2.2 百合TPS 蛋白生物信息学分析 依据各类生物信息学工具(表2)在线比对分析LsoTPS-1、LsoTPS-2、LsoTPS-3、LrlTPS-1 和LrlTPS-3 这5 个TPS 蛋白的氨基酸序列并预测其理化性质、亚细胞定位、信号肽和蛋白质二、三级结构,以此揭示有香和无香百合中TPS 蛋白间的差异,为原核表达载体构建和体外蛋白表达解析提供依据。

表2 蛋白质结构和功能的分析软件和网站
2 结果与分析
2.1 Sorbonne 和Red Life 香气物质及差异分析
从表3 可以看出,2016 年Sorbonne 中共检测出18 种挥发性物质,其中,主要挥发性物质是(E)-β- 罗勒烯,其次是苯甲酸甲酯。2017 年Sorbonne中共检测出25 种挥发性物质,主要挥发性物质也是(E)-β- 罗勒烯和苯甲酸甲酯,较2016 年多检测出11 种挥发性物质,相对含量均小于3.53%,整体偏低,但也有4 种挥发性物质未检测到,这可能是2016 年试验未做重复,试验误差偏大造成的。2018 年Sorbonne 中共检测出28 种挥发性物质,主要挥发性物质仍是(E)-β- 罗勒烯,其次是苯甲酸甲酯;此外,也检测出新挥发性物质(前2 a 未检测出的物质)10 种,相对含量为0~11.19%,但也有11 种挥发性物质未检测到,这可能是不同栽培环境影响了其挥发性物质的释放。

表3 Sorbonne 和Red Life 挥发性物质种类和相对含量差异
综合分析Sorbonne 在3 a 的GC-MS 检测结果发现,(E)-β- 罗勒烯和苯甲酸甲酯是其主要挥发性物质,其中,(E)-β- 罗勒烯相对含量更高,约超过32.00%。同时,Sorbonne 在3 a 里都还检测到另外3 种单萜(E,Z)- 别罗勒烯、波斯菊烯、芳樟醇;1 种不规则萜6- 甲基-5- 庚烯-2- 酮;2 种倍半萜α- 法尼烯、α- 柏木烯;另外2 种苯类物质苯甲酸乙酯、苯甲酸苄酯;1 种脂肪酸衍生物惕各酸甲酯。此外,有18 种挥发性物质只在其中1 a 中检测到,相对含量在0~11.19%;有10 种在其中2 a 里检测出,相对含量在0~7.23%,这可能由于它们来自不同生长环境引起的。总之,不管Sorbonne 中挥发性物质的种类和含量如何变化,其主要挥发性物质均是(E)-β- 罗勒烯和苯甲酸甲酯。
由表3 可知,Red Life 中共检测出23 种挥发性物质,其中,鉴定出单萜4 种,倍半萜3 种,苯类物质3 种,苯丙烷类物质1 种,脂肪酸衍生物12 种。其相对含量最高的挥发性物质是D- 柠檬烯(26.25%),其次是2- 甲基丁酸甲酯(19.97%),之后是丁酸甲酯(10.77%),而其他挥发性物质的相对含量均不超过8.02%。
综合表明,Sorbonne 中挥发性物质种类较Red Life 更加丰富,且挥发性物质含量也较Red Life 更高。Sorbonne 中的主要挥发性物质是(E)-β- 罗勒烯和苯甲酸甲酯,且单萜类物质的相对含量比苯类物质整体偏高;Red Life 中尽管D- 柠檬烯的相对含量最高,但其单萜种类和含量都整体偏低,且Red Life 中不含Sorbonne 中拥有的(E)-β- 罗勒烯、(E,Z)- 别罗勒烯、月桂烯、波斯菊烯、(-)-Alpha- 胡椒烯和β- 蒎烯,其挥发性物质多为脂肪酸衍生物。由此推测,单萜类物质很可能是引起Sorbonne 和Red Life 香气差异的主要原因之一。
2.2 百合TPS 蛋白的生物信息学分析
2.2.1 TPS 蛋白氨基酸序列及理化性质分析 5 个TPS可以分为3 类,LsoTPS-1和LrlTPS-1的编码区全长1 761 bp,编码586 个氨基酸;LsoTPS-2的编码区全长1 773 bp,编码590 个氨基酸;LsoTPS-3和LrlTPS-3的编码区全长1 647 bp,编码548 个氨基酸。DNAMAN 软件氨基酸序列比对结果表明,相较于LsoTPS-1 和LrlTPS-1 蛋白,LsoTPS-3 和LrlTPS-3缺失了38 个氨基酸,LsoTPS-2 插入了4个氨基酸(图1)。

不同品种中同一类TPS 存在氨基酸突变,LsoTPS-1 与LrlTPS-1 第316 位置的氨基酸不同,由缬氨酸(Val)变为丙氨酸(Ala),都属于脂肪族类氨基酸;LsoTPS-3 与LrlTPS-3 有3 个氨基酸不同,第80 位置的亮氨酸(Leu)变为脯氨酸(Pro),前者属于脂肪族类氨基酸,后者属于亚氨基酸;第230 位置的组氨酸(His)变为精氨酸(Arg),都属于碱性氨基酸;第531 位置的异亮氨酸(IIe)变为蛋氨酸(Met),即由脂肪族类氨基酸变为含硫类氨基酸(图1)。
使用ProtParam 分析5 个TPS 蛋白氨基酸理化性质,结果显示(表4),5 个TPS 蛋白分子质量均在65 ku 左右,理论等电点介于5.50~6.00,负电荷残基Asp+Glu 数目都大于正电荷残基Arg+Lys,推测为酸性蛋白;LrlTPS-3 蛋白不稳定系数为40.78(不稳定系数<40 时稳定),稳定性较差,其余蛋白不稳定系数都小于40,均为稳定蛋白;5 个TPS 蛋白总平均疏水值小于0,均具有亲水性。综上可知,5 个TPS 蛋白既是酸性蛋白,又是亲水蛋白,除LrlTPS-3 蛋白外,其余都是稳定蛋白。
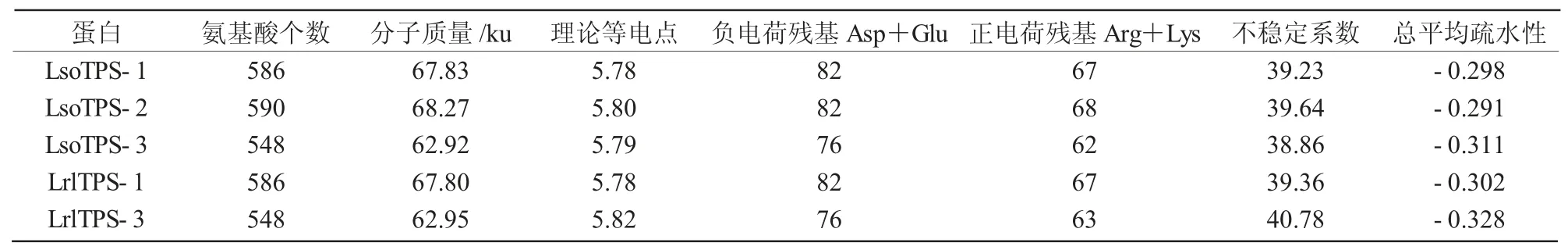
表4 不同TPS 蛋白一级结构预测
2.2.2 TPS 蛋白信号肽和亚细胞定位预测 利用SignalP3.0 预测5 个TPS 氨基酸序列,发现这几种蛋白均不存在信号肽,推测为非分泌蛋白质,无需在内质网膜上合成。运用TargetP1.1 Server 进行亚细胞定位预测发现,5 个TPS 蛋白均可能具有叶绿体转运肽,其中,LsoTPS-2 可靠级别3 级,其余可靠级别2 级(表5)。经ChloroP1.1 Server 分析发现,同一品种中不同类TPS 的叶绿体转运肽长度不一样,但不同品种中同一类TPS 叶绿体转运肽的长度相 同,LsoTPS-1 和LrlTPS-1、LsoTPS-2、LsoTPS-3和LrlTPS-3 蛋白的叶绿体转运肽分别有31、41、15个氨基酸残基(表5)。由此推测,TPS 蛋白在游离核糖体上合成后,直接运输到叶绿体中行使功能。
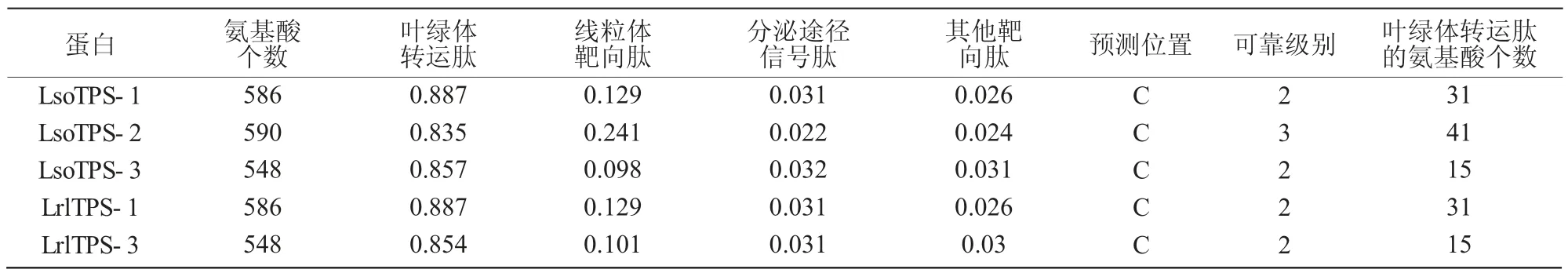
表5 不同TPS 氨基酸序列亚细胞定位预测
2.2.3 TPS 蛋白二、三级结构预测 通过SPOMA预测5 个TPS 氨基酸序列的二级结构发现,这5 个蛋白以α- 螺旋(65.69%~67.92%)为主,β- 折叠仅占3.5%(表6)。由于5 个蛋白与数据库中(+)- 柠檬烯合酶的同源性均高于40%,因此,可使用SWISSMODEL软件进行三级结构同源建模。结果显示,5 个蛋白具有丰富的α- 螺旋,且由2 个结构域组成,它们之间的重要差异主要在2 个结构域的连接处(图2)。

表6 百合TPS 蛋白二级结构主要构成组件比例分析

3 结论与讨论
花香通常被称为“花的灵魂”,是挥发性有机化合物(volatile organic compounds,VOCs)赋予观赏植物的一个重要品质指标[11]。组成花香的VOCs 根据其代谢来源可分为三大类:萜烯类化合物、苯类/苯丙烷类化合物和脂肪酸衍生物[12],其中,萜类化合物是植物次生代谢产物中最大且结构最多样的一类[13-15]。在浓香百合中萜类化合物就占了极大的比例,尤其是(E)-β- 罗勒烯、桉树脑等多种单萜类物质[16]。张辉秀等[8]和唐彪等[3]的研究也指出,浓香的东方和麝香百合中单萜类物质释放量高,而淡香的亚洲和麝亚百合中香气成分主要是烷烃类物质,且未检测到萜烯类物质。此外,相关学者还发现,百合香气成分的释放量会随花期先增大后减小,盛花期达到最大,也会随日照出现从早到晚先增加后减少的变化[2,17]。本研究发现,Sorbonne 中的主要挥发性物质是(E)-β- 罗勒烯和苯甲酸甲酯,不会随年份发生改变,其中,(E)-β- 罗勒烯相对含量更高;但Red Life 中单萜类物质种类和含量都整体偏低。范燕萍等[18]研究发现,东方百合西伯利亚特征香气成分是苯甲酸乙酯,亚洲百合精粹的挥发性物质含量很少,无香。可见,不同有香百合品种的主要芳香成分不同,且不同无香品种的挥发性物质释放量都偏低,从而导致其无香。
单萜类化合物是百合花致香的关键成分[18],一般是通过MEP 途径在TPS 的催化下合成的,TPS是MEP 生物合成途径最后一步的酶[13]。TPS 的种类和功能决定了萜烯类物质的多样性,因此,对TPS结构和特性的解析是利用其进行生物反应器生产目标物质的重要基础,也为揭示有香和无香百合中萜烯类物质在蛋白水平上的差异提供了理论依据。
Sorbonne 中的LsoTPS-1、LsoTPS-2 和LsoTPS-3 与Red Life 中的LrlTPS-1 和LrlTPS-3 这5 个蛋白之间存在氨基酸差异位点,它们的三级结构也具有不同之处。上述差异很可能会影响它们的蛋白功能。孙静[19]研究发现,秋菊神马和夏菊优香中的CmFTLps1、CmFTLps2、CmFTLps3 和CmFTLps4,这4 个转录本在序列上存在18 个氨基酸位点的差异,而且在空间构象上也存在差异,经遗传转化研究后发现,除CmFTLps4 外其余均能回补其晚花表型,由此推测,CmFTLps4 功能缺失与4 个氨基酸差异位点有关。生物信息学分析发现,5 个TPS 蛋白不具信号肽,但很可能具有叶绿体转运肽。ABBAS等[20]和ZHANG 等[11]通过亚细胞定位试验证明,百合Siberia 中的LoTPS1、LoTPS3 和LiTPS2 分别存在于质体、线粒体和叶绿体中。
本研究使用气质联用仪检测了浓香Sorbonne和无香Red Life 中的挥发性物质,揭示二者香气成分差异,并利用生物信息学方法比较了Sorbonne 和Red Life 中5 个TPS 蛋白之间的差异,从分子水平进一步揭示二者花香差异原因,为之后研究TPS 蛋白功能奠定基础。
猜你喜欢
康复(2022年25期)2022-11-18
分子催化(2022年1期)2022-11-02
热带作物学报(2022年8期)2022-09-16
棉纺织技术(2022年4期)2022-04-15
食品安全导刊(2021年21期)2021-08-30
课堂内外(小学版)(2021年5期)2021-01-17
法制博览(2020年27期)2020-11-30
环境(2019年4期)2019-04-20
孩子(2019年2期)2019-02-20
中学生博览·文艺憩(2017年8期)2018-03-05
