
不同更新方式对亚热带常绿阔叶林土壤氮矿化的影响*
2021-05-21 08:07高艳丽杨智杰熊德成
林业科学 2021年4期
高艳丽 杨智杰 张 丽 熊德成
(福建师范大学地理科学学院 福建三明森林生态系统与全球变化野外科学研究站 福州 350007)
我国亚热带森林作为重要的商品林区,长期以来大量的天然林经皆伐、火烧等转换为人工林、经济林(杨玉盛等, 2007; 张睿等, 2015),森林转换造成了大量的水土流失和林地养分损失(杨萌等, 2016)。人工促进天然更新林(人促更新林)作为一种近自然育林经营模式,不仅有天然林的维持土地肥力作用,而且也有人工林卓越的速生丰产功能(孟祥江等, 2018),是较合理的森林培育方式(吴水荣等, 2015; 惠刚盈等, 2018)。氮素是土壤营养元素最重要的组分之一(Geetal., 2018),土壤氮矿化作用通常被认为是森林生态系统氮循环的重要组分(赵阳等, 2013; 李贵才等, 2001)。因此,探讨人促更新林模式下的土壤氮循环成为深入了解人工营林生态系统养分流动的重要内容。
大量研究发现,森林转换中火烧整地、皆伐营林等措施给植被群落及地表层带来剧烈扰动,对土壤养分循环影响显著(杨玉盛等, 2003; 秦倩倩等, 2018; 肖文娅等, 2018)。亚热带天然林转换为人工林后易引起土壤氮矿化速率降低、矿化作用减弱和氮素流失(宋庆妮等, 2013; 袁磊等, 2017)。而人促更新林是以保护和抚育措施为主的干预营林模式,在维持土壤团粒结构、地上地下养分循环、改善土壤肥力与质量方面的优点比传统经营的人工林更明显(黄清麟等, 2000; 邱仁辉等, 2001; 陈绍栓等, 2001; 何朋俊等, 2017)。人促更新林的采伐剩余凋落物覆盖措施利于维持林地原状并增加凋落物归还量,同时皆伐整地等常规人工更新的高强度干扰的减少可进一步降低土壤氮损失,使林地原位土壤有机质和营养元素得到更好保存(孟祥江等, 2018; 李光耀等, 2014)。但是,有关对人促更新林土壤氮素有效性和氮矿化作用的研究还十分缺乏。同时,目前大多数林型转换前后树种有明显变化(邢肖毅等, 2013; Jinetal., 2019),导致不同树种的养分利用需求策略有较大差异,这是森林转换后研究结果争议较大的原因之一(李金华等, 2017; 丁国昌等, 2017)。
人促更新林已成为一种有效的森林转换经营模式,在全球大量推广应用。米槠(Castanopsiscarlesii)林是福建省分布较广、极具中亚热带代表性的森林类型。本研究选取福建省三明市典型的米槠天然林及其转换后的米槠人促更新林和米槠人工林3种森林类型,比较其表层土壤有效氮含量及土壤氮矿化和硝化速率的差异,分析亚热带米槠常绿阔叶林转换为人促更新林和人工林后的土壤氮矿化作用的变化,为评价和选择森林经营方式提供参考。
1 研究区概况
研究区位于福建省北部三明市的格氏栲自然保护区和金丝湾森林公园陈大林场(117°28′—117°36′E,26°11′— 26°19′N),两地区直线距离不超过30 km。土壤类型分别为砂岩和花岗岩发育的红壤。该区属中亚热带季风性气候,年均降水量1 749 mm,年均气温19.1 ℃,区域内典型植被为常绿阔叶林,植物多样性丰富。保护区及陈大林场内有米槠的天然林及受不同强度人工干扰后形成的约40年生的次生林和人工林。这些林分的本底条件相似,构成了不同转换方式的林分类型对比,为本研究提供了良好的研究对象。3种林分的植被概况和表层土壤性质见表1和2。
仅分布在格氏栲保护区(117°28′E,26°11′N)内的米槠天然林面积近190 hm2,其林龄为260~300年,乔木层伴生树种主要有米槠、木荷(Schimasuperba)、格氏栲(Castanopsiskawakamii)、桂北木姜子(Litseasubcoriacea)和杜英(Elaeocarpusdecipiens)等,灌木层(树高小于3 m)主要有米槠、黄丹木姜子(L.elongata)、赤楠(Syzygiumbuxifolium)和冬青(Ilexpubescens)等,草本层由草珊瑚(Sarcabdraglabra)、山姜(Alpiniajaponica)和扇叶铁线蕨(Adiantumflabellulatum)等组成。
陈大林场(117°36′E,26°19′N)内的米槠人促更新林是由米槠天然林经过择伐、保留目标树种、劈除杂灌和人工扶持后经封禁保护形成的。乔木层伴生树种主要有米槠、木荷、东南野桐(Malloyuslianus)和白檀(Symplocospaniculata)等。灌木层以鼠刺(Iteachinensis)、毛叶冬青(Ilexpubilimba)和石栎(Lithocarpusglabra)等为主。草本层有鳞籽莎(Lepidospermachinense)、狗脊蕨(Woodwardiajaponica)和扇叶铁线蕨等。
陈大林场(117°36′E,26°19′N)内的米槠人工林是由采伐3年内的次生林经过皆伐、火烧、挖穴造林和幼林抚育形成的。林下伴生树种主要有冬青、乌饭树(Vacciniumbracteatum)、薄叶山矾(Symplocosanomala)、桂北木姜子和山姜等。
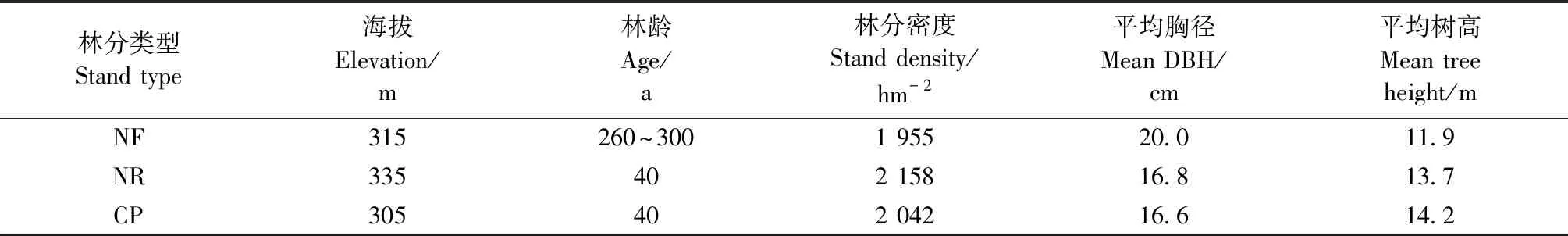
表1 不同林分类型植被概况Tab.1 Vegetation properties of various stand types
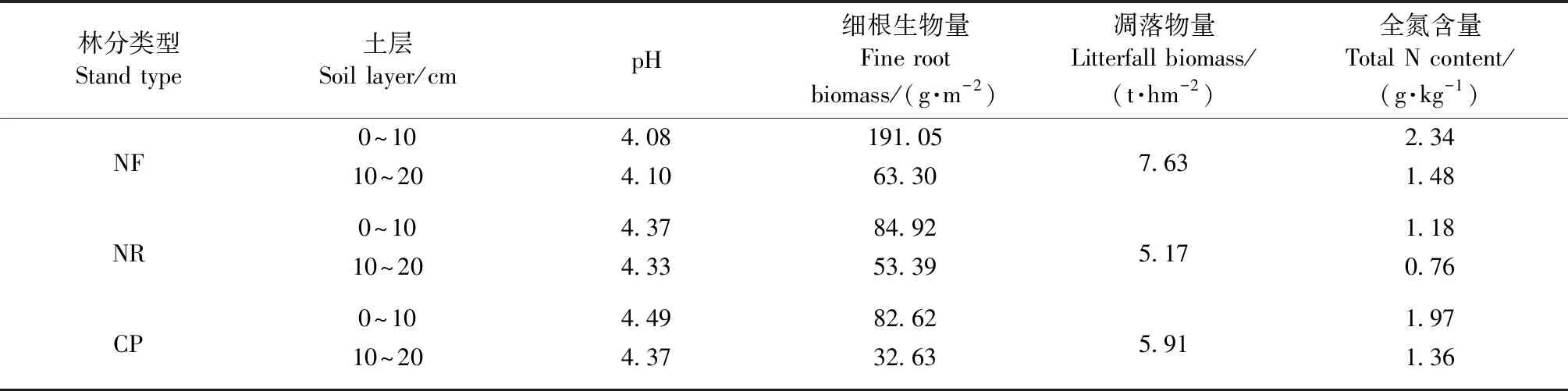
表2 不同林分类型表层土壤性质Tab.2 Top soil properties of various tand types
2 研究方法
2.1 土壤样品采集与氮含量测定
2018年4月,在3种林分类型分别设3块20 m×20 m 样地,在每块样地中按“S”形设置5个取样点采集土样,采集0~10和10~20 cm土层土样,每种林分类型各个土层的土壤样品混合成1个土壤样品,共6个土壤样品。用保温箱储存土壤样品带回实验室,手工拣去石砾和动植物残体,过2 mm土壤筛后,冷冻储藏备用。
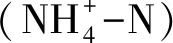
2.2 土壤氮矿化培养试验
称取过2 mm筛的相当于10 g干土质量的鲜土样,平铺于300 mL广口瓶底部,加去离子水至田间持水量的50%,用保鲜膜及皮筋将其密封,并用针在保鲜膜上扎数个通气小孔以确保培养瓶的好气环境,并将培养瓶置于25 ℃恒温箱中预培养3天(每种林分类型每个土层做18个重复,加上2个空白处理,共110个样品)。再次调节土壤含水量为50%,然后进行正式培养,期间所有样品瓶每隔3天将瓶口打开通气30 min,并再次调节土壤含水量为50%。在正式培养的第0、3、7、14、21和28天,每个林分每个土层取出3个样品瓶,每个样品瓶破坏性取10 g土壤样品,用2 mol·L-1KCl溶液浸提土壤矿质氮,使用流动分析仪测定其中铵态氮和硝态氮含量。
2.3 数据处理
利用培养前后土壤铵态氮和硝态氮含量的差值及培养天数,计算土壤净硝化速率及净氮矿化速率。参考Wei等(2010)的计算方法:
(1)
(2)
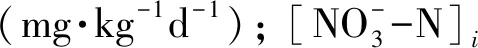
采用SPSS21.0软件对数据进行统计分析。采用单因素方差分析(one-wayANOVA)检验不同林分的数据差异。通过Origin9.1软件绘图。
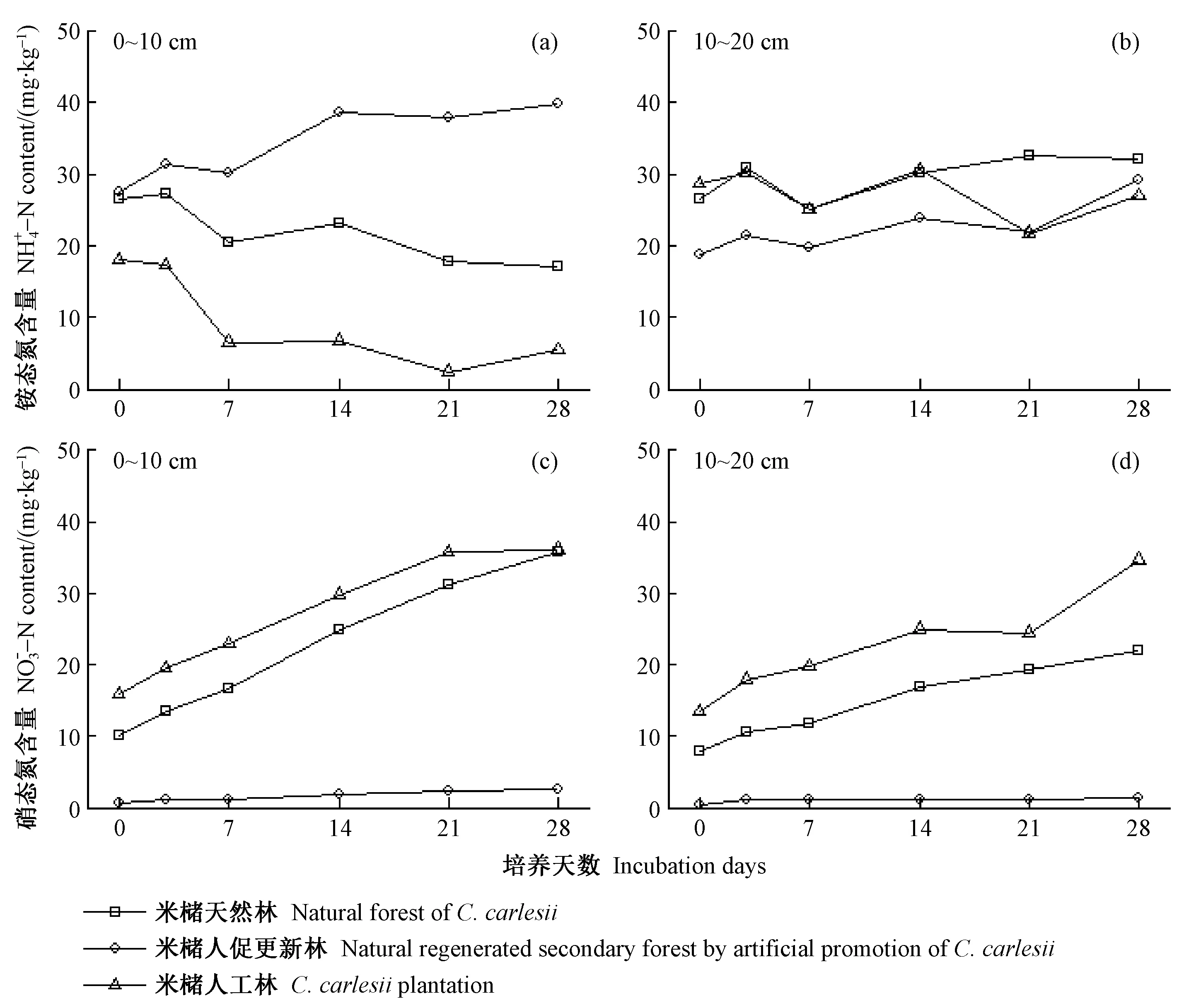
图2 不同林分类型表层土壤铵态氮和硝态氮含量变化趋势Fig. 2 Variation trend of ammonium nitrogen and nitrate nitrogen contents in topsoil of various stand types
3 结果与分析
3.1 林分类型转换对土壤铵态氮和硝态氮含量的影响
3.2 林分类型转换对土壤净氮矿化速率和硝化速率的影响
由表3可知,与米槠天然林相比,人促更新林0~10和10~20 cm土层土壤净氮矿化速率分别降低了12%和11%,人工林分别降低50%和27%; 人促更新林2个土层净氮矿化速率高出人工林76%和22%。
与米槠天然林的土壤净硝化速率相比,人促更新林降低,人工林的则增加。人促更新林0~10和10~20 cm土层的土壤净硝化速率比天然林分别降低93%和92%,比人工林分别降低921%和95%。米槠人工林0~10 cm土层的土壤净硝化速率比天然林降低了21%,10~20 cm土层增加了52%。
总体来看,米槠人促更新林的土壤净氮矿化速率高于米槠人工林,而土壤净硝化速率则远低于米槠人工林。
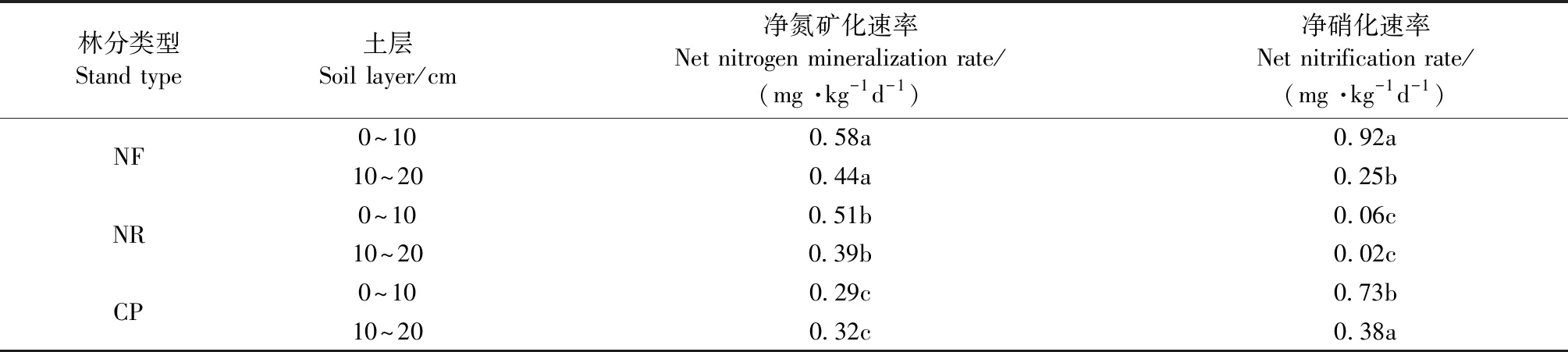
表3 不同林分类型表层土壤氮矿化速率和硝化速率①Tab.3 Nitrogen mineralization and nitrification rats in topsoil of various stand types
4 讨论
4.1 林分类型转换对土壤有效氮含量的影响
本研究中,米槠天然林、人促更新林和人工林的土壤有效氮含量都以铵态氮为主,与许多热带亚热带森林的研究结果相似(王磊等, 2016; 崔晓阳等, 2005; 王光军等, 2009; 李志萍等, 2013)。崔晓阳等(2005)认为,大多酸性、弱酸性的森林土壤铵态氮含量远高于硝态氮;王光军等(2009)在研究湖南省森林土壤氮矿化时发现,4种森林的土壤无机氮都以为铵态氮为主,占70%~80%,而硝态氮占比则很小。可能原因是亚热带森林酸性土壤特有的保氮机制抑制了土壤自养硝化过程,土壤的反硝化势低(续勇波等, 2014),使土壤氮素以铵态氮为主。亚热带地区高温多雨,因硝态氮极易随土壤溶液移动(Mcbratney,2001),所以土壤中的硝态氮较铵态氮更易通过降雨淋洗和地表径流迁移和淋失(陈伏生等, 2004),因此降低了土壤硝态氮含量。而在川西亚高山地区,受气候、植被、微生物和土壤底物等的综合影响,典型的云杉(Piceaasperata)人工林及桦木(Betulaspp.)次生林土壤氮均以硝态氮为主(李志杰等, 2016),表现出与本研究区不同的氮保留机制。
森林类型转换后土壤有效氮含量下降,这与宋庆妮(2013)、李明锐等(2005)的研究结果相似,其重要原因是转换方式的干扰程度不断增强。但在Yan等(2008)的研究中,常绿阔叶林转换为次生灌木、杉木(Cunninghamialanceolata)人工林等后,铵态氮和硝态氮含量都有不同程度的增加,这可能是因经营历史及物种组成差异造成的。在本研究中,米槠人促更新林表层土壤铵态氮含量高于米槠人工林,但硝态氮含量低于人工林。人工林经营过程中受到人为营林措施的强烈干扰,导致土壤结构发生变化,加剧了水土流失,人工林的原位土壤养分积累优势在剧烈的森林转换过程中丧失(Yanetal., 2008); 由表2可知,人促更新林的细根生物量较多,可通过密集的根系吸收被矿化的土壤氮素。同时根系可能释放生物硝化抑制剂来调控氮转化循环,提高氮素利用效率,进而避免有效氮的快速反硝化和淋失(Subbaraoetal., 2007),因此人促更新林土壤保留了更多铵态氮。由于铵态氮是亚热带森林有效氮库的主体部分,所以相较于米槠人工林,铵态氮含量更高、硝态氮含量低的米槠人促更新林更利于维持土壤氮素。
4.2 林分类型转换对土壤氮矿化速率和硝化速率的影响
林分类型转换后林地表层土壤氮矿化速率表现为米槠天然林>米槠人促更新林>米槠人工林,硝化速率则以人促更新林最低(表3)。对川西亚高山3个典型森林群落的研究发现(李志杰等, 2016),土壤净氮矿化速率和净硝化速率总体表现为云杉人工林和桦木次生林都显著低于天然林,其氮矿化速率降低与本研究结果相同,可能反映了人工营林期间的强烈干扰(如采伐和土地扰动)对森林养分循环的影响(闫恩荣等, 2007); 但硝化速率结果与本研究不同,则可能是两者在营林历史中的立地管理、区域植被物种组成及局地地理条件差异导致的。本研究的米槠人促更新林土壤氮矿化速率高于江西中部的亚热带杉木人工林、马褂木(Liriodendronchinense)人工林(宋庆妮等, 2013),这可能是因人工林的土壤养分积累和林冠层遭受严重干扰、凋落物质量和数量的输入减少(表2),导致了其净氮矿化速率低(李志萍等, 2013)。相比而言,人促更新林营林过程中土壤受扰动少,采伐剩余物保留堆积,林地劈除杂灌封禁保护,人工促进形成目的树种的复层混交林,凋落物输入量丰富,具备更多氮矿化有机底物,因此氮矿化速率高于人工林。
本研究中的米槠人促更新林表层土壤净氮矿化速率高出人工林20%~75%,净硝化速率比人工林低90%以上,可能是因人促更新林内植被组成更丰富、林地细根生物量较高、凋落物量大且类型丰富(表2)。天然林转换后通常会削弱林地土壤矿化作用,但采取有效恢复措施保护次生林,土壤氮素有效性则会有所提高(Xuetal., 2008)。本区域内米槠人促更新林在营造过程中采取的采伐剩余物保留、非目的树种择伐等人工抚育措施,可能与其总氮含量和氮矿化速率显著高于人工林有关,这可能是因为林地受干扰程度低,采伐剩余物堆积能增加凋落物质量,凋落物在土壤微生物等的作用下快速得到分解,将大量氮素归还到土壤表层(曾晓敏等, 2018)。人促更新林表层土壤硝化速率低,可能是较低的土壤pH值和根系生物硝化抑制作用(BNI)限制了硝化微生物繁育,从而对土壤硝化作用产生一定抑制(朱兆良, 2008; Subbaraoetal., 2007)。硝化作用较弱有利于降低硝态氮损失风险和维持铵态氮固存,也利于森林表层土壤总氮含量增加。因此米槠人促更新林能有效保存大量有效氮,供给植物和微生物利用,增加土壤养分积累,提高土壤肥力,相比于米槠人工林有更好的氮储存能力和氮循环过程。
5 结论
亚热带地区米槠天然林转换为人促更新林和人工林后,土壤铵态氮含量下降,硝态氮含量略有增加,总体有效氮含量明显降低。但人促更新林的土壤铵态氮含量和氮素有效性均高于人工林。与米槠天然林相比,人促更新林和人工林的土壤净氮矿化速率下降,但人促更新林土壤净氮矿化速率仍高于人工林,且土壤净硝化速率远低于人工林。人工林的有效氮固持能力差,氮素流失风险大,而人促更新林的有效氮含量高且流失风险小、氮矿化作用更强。因此,米槠人促更新营林方式比人工林营林方式更利于维持良性土壤氮循环过程和保障氮素供给。
猜你喜欢
土壤学报(2022年2期)2022-06-09
水土保持学报(2022年3期)2022-05-26
建材发展导向(2022年6期)2022-04-18
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
山东农业大学学报(自然科学版)(2020年5期)2020-11-03
核农学报(2020年5期)2020-04-10
环境与发展(2019年12期)2019-02-12
农家顾问(2016年5期)2016-05-14
