
微生物菊糖蔗糖酶及在食品中的应用研究进展
2021-05-19 02:22:30倪大伟张文立沐万孟
食品科学 2021年7期
倪大伟,徐 炜,张文立,2,沐万孟,2,
(1.江南大学 食品科学与技术国家重点实验室,江苏 无锡 214122;2.江南大学 食品安全国际合作联合实验室,江苏 无锡 214122)
自然界中存在两种果聚糖,按照果糖基之间糖苷键的差异,可分为β-(2,1)糖苷键连接的菊糖和β-(2,6)糖苷键连接的左聚糖(图1)。左聚糖在植物中含量极少,目前工业上主要以蔗糖为底物,通过果聚糖蔗糖酶(levansucrase,LSase)生产左聚糖[1];相反地,菊糖在一些植物中含量丰富,尤其是菊科植物,如菊苣、菊芋和大丽花等[2]。作为一种益生元和膳食纤维,菊糖具有许多显著的生理功能,如抗氧化、促进矿物质吸收、缓解便秘等[2]。食品工业中,菊糖作为膨化剂、增稠剂、脂肪替代品等,常用于冰淇淋、巧克力、酸奶等的生产[3]。
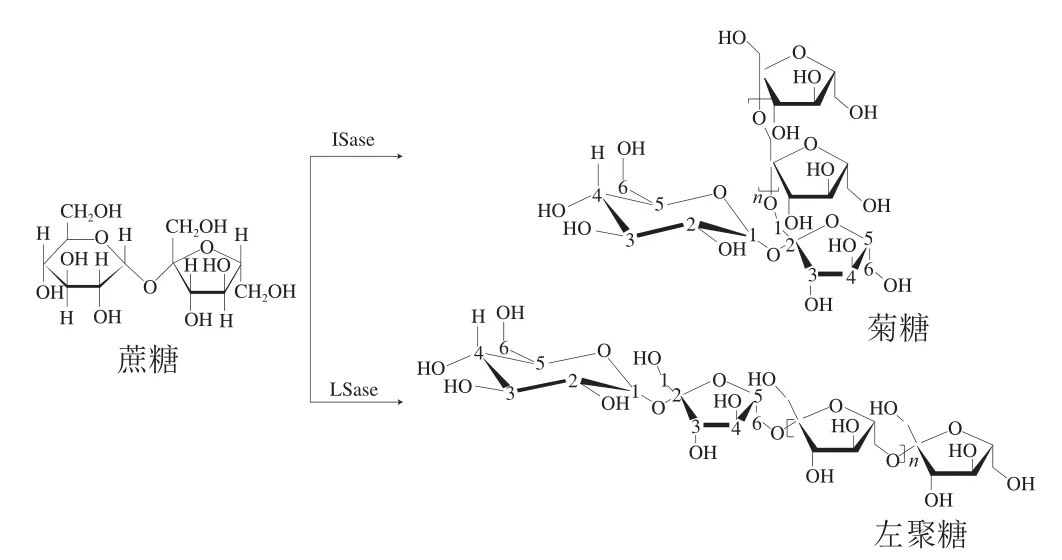
图1 菊糖和左聚糖结构式[1-2]Fig.1 Structures of inulin and levoglycan[1-2]
一些微生物可以通过菊糖蔗糖酶(inulosucrase,ISase)合成菊糖。微生物菊糖与植物菊糖相比,其分子质量是植物菊糖的数百倍。近年来,以蔗糖为唯一底物,利用ISase合成微生物菊糖受到人们关注。作为一种兼具水解和转糖基能力的酶,ISase也能够以蔗糖为果糖基供体,以某些其他糖类为果糖基受体,合成新型低聚糖。
ISase自1973年被发现以来[4],在新酶挖掘与性质鉴定、构效关系与分子改造等方面的研究不断被报道。然而,国内关于该酶的研究鲜有报道。本文从近年来学者们对于ISase结构、功能和催化机理的研究出发,总结了在酶分子改造方面,尤其是其产物——菊糖链长调控方面的研究进展。重点讨论了ISase在食品领域的应用价值,并对其发展趋势提出合理的展望。
1 ISase的晶体结构
酶分子的结构决定其功能,对酶结构的解析可以通过酶分子改造手段而改善酶的性质,提高酶的应用价值。ISase和LSase同属于糖苷水解酶GH68家族(http://www.cazy.org/),按照催化口袋的整体构像,又属于Clan-J家族。目前,总计15 种微生物来源的ISase被鉴定,仅有一种来自Lactobacillus johnsoniiNCC533的ISase(Laje-ISase)被解析了晶体结构[5]。图2和图3A分别展示了Laje-ISase的二级结构和三维结构。ISase的整体构象由5 个β-折叠的结构域组成,每个结构域又由4 个反相平行的β-折叠组成。20 个β-折叠共同构成了漏斗状的催化口袋,这种催化口袋的构象称为“5-foldβ-propeller”。
除了裸蛋白结构,Laje-ISase与蔗糖和蔗果三糖复合物的共晶结构也被解析出来。由此揭示了在酶催化反应过程中起关键作用的3 个氨基酸(称为催化三联体),具体为两个天冬氨酸和一个谷氨酸(D272、D425和E524),分别起到亲核试剂、过渡态稳定剂和提供催化质子酸/碱的作用(图3B)。Laje-ISase的结构显示,在催化口袋的外围,有一个Ca2+结合位点,因此,Ca2+可能对酶催化反应活力和稳定性起到一定的影响。一些对酶学性质的研究证明了这一点,如Lactobacillus reuteri121来源的ISase在添加1 mmol/L CaCl2的条件下,酶活力提高了20%以上[6]。
三联体活性位点的协同作用是ISase和LSase催化反应的共同特征。晶体结构的研究表明,ISase和LSase的催化口袋均采用“5-foldβ-propeller”构象,并且两种酶三维结构的重合度很高。相似的结构导致ISase和LSase的催化机理和反应进程基本一致。目前,5 种微生物来源的LSase晶体结构已被解析,分别来自Bacillus subtilis[7]、Gluconacetobacter diazotrophicus[8]、Erwinia amylovora[9]、Bacillus megaterium[10]和Erwinia tasmaniensis[11]。ISase和LSase结构上的不同主要在于ISase中无规卷曲结构多,从而蛋白的柔性较大。目前仅解析出一种来源的ISase结构,这可能与其蛋白分子柔性大,不易结晶有关。

图2 Laje-ISase的二级结构[1,5]Fig.2 Secondary structure of Lactobacillus johnsonii NCC533-ISase (Laje-ISase)[1,5]
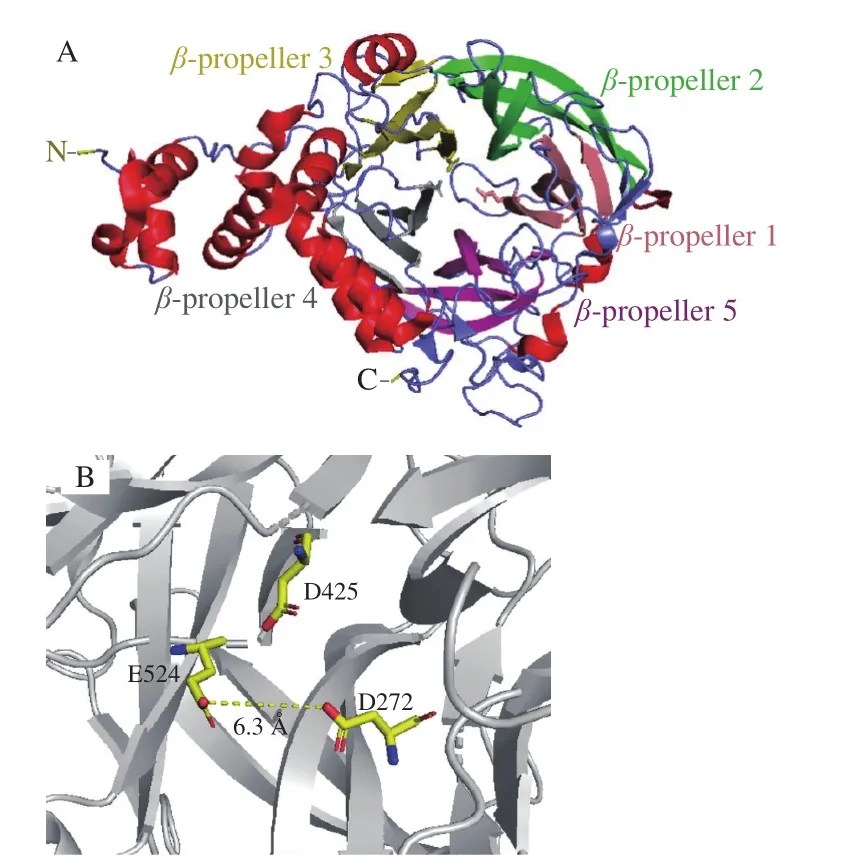
图3 Laje-ISase的三维结构图[5]Fig.3 Three-dimensional structure of Laje-ISase[5]
2 ISase的催化机理
根据糖苷键水解后异头碳上羟基的构象,糖苷酶的反应机制分为保留机制和反转机制两种。通过对糖苷酶的结构分析,涉及催化反应的一对羧基的距离决定了两种不同的反应机制。糖苷酶的催化位点由两个酸性氨基酸组成,若两个氨基酸上羧基的距离在5.5 Å左右,则为保留机制;距离在10 Å左右,则为反转机制[12]。Laje-ISase在催化过程中,D272和E524的羧基参与反应,二者羧基氧相距6.3 Å(图3B)。因此,可以判定ISase采取保留机制,催化糖苷水解。
ISase以蔗糖为唯一底物时,表现出3 种主要功能:1)剪切蔗糖;2)将果糖基转移到另一分子蔗糖上;3)延伸菊糖链。因此,当ISase利用蔗糖为唯一底物时,会发生3 种连续的反应,见图4A。首先,蔗糖的果糖基和葡萄糖基分别结合在ISase的-1和+1位点,D272在催化过程中起到亲核试剂的作用,其羧基上的羟基“攻击”蔗糖果糖基的C2。与此同时,E524羧基上的羟基提供质子酸,形成酶-果糖基复合体中间产物。D425的作用是稳定这一过渡态中间体。其次,在水分子参与下,E524羧基上的羟基氧提供质子碱,果糖基脱离,发生水解反应;相反,如果另一分子蔗糖作为果糖基受体,则会发生转糖基反应,生成蔗果三糖。最后,不断延伸的糖分子作为受体,发生聚合反应,菊糖分子不断延伸。关于ISase正负位点的定义可以理解为:-1位点是供体果糖基的结合位点,+1、+2……+n位点是受体糖链糖基结合位点。糖苷键的断裂和形成,发生在-1和+1位点的交界处。
ISase和LSase具有同样的催化活性中心构象及活性位点,且两种酶的反应进程几乎一致。因此,ISase和LSase的催化机制经常被同时讨论。目前,学者认为ISase和LSase具有同样的催化反应机制。保留机制仅针对糖苷键水解后异头物的构象变化而言。催化反应过程包括两步:蔗糖的断裂和果聚糖的延伸,因此学者们提出了两种不同的反应机制来分别描述这两个过程。

图4 ISase的反应机制[1,13-15]Fig.4 Reaction mechanisms of ISase[1,13-15]
如图4A所示,ISase和LSase采取“乒乓机制”,亦称双置换机制剪切蔗糖,形成葡萄糖和酶-果糖基复合体中间产物[13]。如图4B所示,“延续性机制”和“非延续性机制”被用来解释高分子质量果聚糖和低分子质量果聚糖的形成[14-15]。在果聚糖链延伸的过程中,形成的果聚糖始终结合在酶上,这种糖链的延伸方式称为“延续性机制”;相反,如果糖链在每次结合新的果糖基后都会从酶上脱落再结合,这种延伸方式称为“非延续性机制”。“延续性机制”下形成长链产物,而“非延续性机制”形成短链产物。
在酶分子采取双置换机制剪切蔗糖时,3 个活性位点构成的催化三联体起着绝对的作用。然而,目前关于糖链延伸的“延续性机制”和“非延续性机制”所涉及的关键氨基酸残基还没有被揭示。
3 ISase产物的链长调控
ISase以蔗糖为唯一底物时,能够合成菊糖。菊糖的分子质量因ISase的微生物来源不同而存在差异。微生物菊糖的分子质量通常比植物来源的菊糖高出100~1 000 倍。目前已经测定出分子质量的微生物菊糖不多,包括L.johnsoniiNCC 533(4×107Da(分子质量,下同))[16]、Streptomyces viridochromogenesDSM 40736(2.5×107Da)[17]、Streptococcus mutansJC-2(2×107Da)[18]、Streptococcus mutansGS-5(7×107Da)[19]、L.reuteri121(>107Da)[6]以及Lactobacillus gasseriDSM 20604(5.858×106Da)[20]。与以上只合成单一分子质量的ISase不同,Leuconostoc citreumCW28来源的ISase合成的菊糖,其分子质量有两个范围,分别是低分子质量2.6×103~3.4×103Da和高分子质量1.35×106~1.6×106Da。Bacillus agaradhaerensWDG185来源的ISase是目前已知仅能合成低分子质量菊糖的ISase,合成的菊糖聚分子质量为3×103Da[21]。Aspergillus sydowiIAM 2544是目前鉴定的唯一能合成菊糖的真菌,合成的菊糖分子质量为7×107Da[4]。
多糖的分子质量是影响其性质和功能的重要因素[22-23],植物菊糖的性质也受分子质量影响[24-25]。通过改变反应条件或利用分子改造的方法,获得特定分子质量的菊糖,是ISase研究和应用的方向。LSase合成左聚糖的分子质量受到反应条件(底物蔗糖浓度、加酶量和离子强度等)的影响[26]。分子改造是研究ISase产菊糖链长调控的主要方式。
Ozimek等[27]报道了对ISase产物链长调控的研究,通过研究L.reuteri121来源的ISase,对其-1位点上完全保守的3 个氨基酸进行单点突变,获得3 个改变产物链长的突变体W271N、W340N和R423K。W271N和R423K除了能合成菊糖外,还能合成更多聚合度大于10的低聚果糖,但R423K不能合成聚合度6~10的低聚果糖;W340N对产物链长的影响较大,仅能合成蔗果三糖和蔗果四糖。Anwar等[28]也构建了该酶的突变体N543,仅能合成聚合度小于6的低聚果糖,失去合成长链菊糖的能力。Rodríguez-Alegría等[29]通过对活性位点附近残基进行突变,获得3 个改变产物菊糖链长的突变体(S425A、L499F和R618K)。其中S425A和R618K失去合成高分子质量菊糖的能力,产物中只有低聚果糖,表明这两个位点可能对控制菊糖链的延伸起重要作用;L499F保留了合成长链菊糖的能力,同时产物中也出现了低聚果糖。
对于链长调控的研究,主要集中在对L.reuteri121来源的ISase上。酶法合成多糖的链长调控主要通过以下两种方式:1)消除或减弱底物与酶结合位点的相互作用,可通过将残基突变为丙氨酸的方式实现;2)阻断底物结合和糖链延伸的通道,可通过将残基突变为芳香族氨基酸的方式实现[30]。
通过对84 种糖酶的分析发现,在糖分子与酶的结合位点附近,与糖分子产生相互作用的氨基酸,80%以上是酸性氨基酸和极性氨基酸[31]。据此,Charoenwongpaiboon等[31]选择L.reuteri121来源的ISase底物结合位点周围、酶分子表面的一些酸性氨基酸和极性氨基酸作为突变位点,发现了两个可能存在的糖链延伸通道:通道A(N543、W551、R483、D479、S482和D478)和通道B(N543、N561、N555和D689)。将这些氨基酸单点突变为丙氨酸,以消除可能与糖链存在的氢键或极性相互作用。结果发现,这些突变体合成长链菊糖的能力均不同程度降低;相反,却合成了更多聚合度小于10的低聚果糖。尤其是突变体R483A,比野生酶产生了更多蔗果六糖(GF5)。Charoenwongpaiboon等[32]针对该位点采用计算模拟的方法进一步研究第二种链长调控方式的作用。只有蔗果五糖(GF4)结合到催化位点,才能经过转糖基作用产生GF5。因此,通过将GF4与突变体R483A、R483F、R483Y和R483W对接,计算结合自由能ΔGbinding,发现除R483W的ΔGbinding不变外,其余突变体的ΔGbinding均减小。进一步对突变体产物链长分布进行研究,发现4 个突变体均比野生酶产生了更多聚合度在4~8的低聚果糖,并且产物的最大聚合度均低于12。
对于ISase的产物链长调控,关键是找出产物低聚果糖结合的更深层次的位点。目前只获得了ISase与蔗果三糖的共晶结构,揭示了糖链结合的+2位点以及与糖基产生相互作用的氨基酸[5]。因此,获得ISase与蔗果四糖等更高聚合度寡糖的共晶结构,是揭示糖链延伸过程更深层次位点的关键,对产物链长调控具有重要意义。
4 ISase在食品领域中的应用
4.1 生产高分子质量菊糖
多糖的分子质量是影响其性质和功能的关键因素。微生物菊糖和植物菊糖的不同在于二者的分子质量,但二者在性质和功能上的差异,目前还没有研究涉及。相关左聚糖的研究显示了高分子质量左聚糖在烘焙方面的优势[33]。面包中添加等质量高分子质量和低分子质量的左聚糖,结果发现,添加高分子质量左聚糖的面包质地更柔软且体积更大[33]。因此,微生物菊糖的潜在价值和应用领域值得进一步挖掘。
目前,工业化应用的菊糖是植物菊糖。虽然其来源广泛,但提取过程复杂,成品中难免会含有其他植物多糖等杂质。酶法合成菊糖是以蔗糖为唯一底物,反应体系中仅含有果糖、葡萄糖、蔗糖、低聚果糖等成分的一种方法,纯化过程简单,产物纯度高。微生物菊糖的生产研究发现L.gasseriDSM 20604来源的ISase在最适条件下可产生53 g/L的菊糖。另外,除了蔗糖,制糖工业的副产品如糖蜜等,也具有作为酶法生产菊糖的潜力,该方法不仅能降低成本,还有利于资源的综合利用。
4.2 生产低聚果糖
低聚果糖作为一种重要的益生元,具有促进肠道菌群平衡等诸多生理功能,被广泛应用于食品和医药等领域[34]。低聚果糖是菊糖形成的前期产物,因此,含有菊糖的植物体内都会积累低聚果糖。目前工业生产的低聚果糖主要通过内切菊粉酶水解菊糖获得[35],或利用呋喃果糖苷酶以蔗糖为底物获得[36]。
通过链长调控的手段控制产物的链长,使ISase失去形成长链菊糖的能力,可以有效提高其生产低聚果糖的能力。通过利用生产低聚果糖的突变体R483A,可制备酶交联聚集体,以50 g/L的蔗糖为底物,在优化的条件下反应4 h,低聚果糖的产量达到12 g/L[37]。利用食品级酿酒酵母蛋白表达系统可异源表达L.reuteri121来源的Isase,通过截断部分序列和引入高效信号肽的方式,使ISase高效表达。分批补料发酵生产聚合度在2~20的中链低聚果糖,从300 g/L的蔗糖中获得128.4 g/L的低聚果糖,转化率达85.6%[38]。最近的研究表明,短链低聚果糖,尤其是蔗果三糖,在改善肠道环境和促进免疫系统方面有着比长链低聚果糖更显著的作用[39]。此外,与其他短链低聚果糖相比,蔗果三糖在甜度、生物活性以及作为益生元等方面表现出更好的性能[40]。Charoenwongpaiboon等[41]通过固定化的手段提高ISase的稳定性和循环利用效率、控制反应条件,成功生产出以蔗果三糖为主要成分的低聚果糖。
4.3 生产新型低聚糖
根据ISase分别以H2O和蔗糖作为受体,发生水解和转糖基反应的这一进程,定义水解活力和转糖基活力。水解活力导致副产物果糖和葡萄糖的产生,转糖基活力则生成菊糖。转糖基的能力是糖基转移酶研究和应用的重点。通过将糖基转移到常见的分子上,形成自然界中少有或没有的且具有高价值的产物,是研究具有转糖基能力的酶的重要方向。研究表明,ISase催化反应过程中,除了H2O和蔗糖可以作为受体分子外,其他一些糖类也可以作为受体,从而形成新型的低聚糖。
Rodríguez-Alegría等[29]首次研究了L.citreum来源的ISase转糖基能力,以蔗糖和木糖或蔗糖和麦芽糖作为共同底物,发现反应体系中,除了生成菊糖和低聚果糖外,木糖和麦芽糖也可以作为果糖基的受体,生成新型低聚糖,但其未对产物低聚糖进行结构鉴定。
将ISase应用于新型低聚糖生产的研究中,L.gasseriDSM 20604来源的ISase是最常用的。通过对以麦芽糖为受体生成的产物进行结构鉴定,表明生成的麦芽糖基果糖苷的聚合度为3~6,且以α-D-吡喃葡萄糖基-(1,4)-α-D-吡喃葡萄糖基-(1,2)-β-D-呋喃果糖苷和β-D-呋喃果糖基-(2,6)-α-D-吡喃葡萄糖基-(1,4)-α-D-吡喃葡萄糖为主要转糖基产物[42]。Díez-Municio等[43]以蔗糖和棉子糖为共底物生产新型低聚糖,并对产物低聚糖进行结构鉴定。果糖基通过β-(2,1)糖苷键连接在棉子糖的果糖基上,形成聚合度为4~8的棉子糖基果糖苷——α-D-吡喃半乳糖基-(1,6)-α-D-吡喃葡萄糖基-(1,2)-β-D-呋喃果糖基-((1,2)-β-D-呋喃果糖苷)n,且最大产率为33.4%。
目前,对LSase转糖基反应的研究更加广泛,这可能得益于LSase更加广泛的微生物来源。许多糖类、酚类和醇都可以作为果糖基的受体,从而形成新的转糖基产物[44-46]。这些新型的低聚糖和衍生物在食品、化妆品和医药等领域都具有潜在的应用价值。因此,获得高转糖基能力的ISase,并筛选可能的受体底物谱,对于提高ISase的应用价值具有重要作用。
4.4 制备菊糖纳米材料
建立在高分子质量左聚糖纳米材料良好性质和应用价值的基础上[47-49],最近,微生物菊糖纳米材料的研究也不断被报道。Jiménez-Sánchez等[50]利用L.citreum来源的ISase生产菊糖,形成了稳定的球状纳米颗粒,颗粒平均直径为112 nm。菊糖纳米颗粒质量浓度在200 μg/mL以下时,对外周血单核细胞不产生毒性作用。温度对菊糖纳米颗粒的形成和颗粒直径影响显著,此外,通过优化温度、蔗糖质量浓度和加酶量等条件,能够获得高产量的菊糖纳米颗粒[51]。将菊糖纳米颗粒与类黄酮(槲皮素和漆黄素)复合,能增加类黄酮的溶解性、稳定性和抗氧化活性[51]。因此,微生物菊糖纳米颗粒可作为一种具有生物相容性的材料,在提高某些天然物质溶解性和防止降解方面具有潜在的应用前景。
5 结 语
蔗糖是自然界中广泛存在的二糖,其价格低、易制取,是食品行业重要的甜味剂。然而,蔗糖由于致龋齿、易导致肥胖等特点,不能满足人们对健康饮食的要求。ISase作为一种果糖基转移酶,能够以蔗糖为唯一底物生产具有良好性质和功能的菊糖和低聚果糖,有潜力成为蔗糖综合利用的重要原料。此外,ISase具有转果糖基的能力,能够以蔗糖为供体,以其他糖类为受体合成某些非天然有价值的糖类。目前,对ISase的研究主要集中在酶学性质、产物鉴定和分子改造等方面。相比于广泛研究的LSase,ISase仍有许多值得进一步开发的方面:1)目前只解析出一种来源ISase的晶体结构,这很大程度上限制了学者们对ISase结构和功能的认知。虽然通过计算辅助的理性设计和同源模拟等方式能够在一定程度上指导分子改造研究,但获得不同微生物来源ISase的晶体结构,对ISase的结构和功能研究更有指导价值;此外,获得ISase与产物寡糖的共晶结构,对于阐明糖链延伸机制具有重要的指导意义。2)受限于ISase的微生物来源较少,在性质鉴定、酶分子结构解析、分子改造以及受体反应生产新型低聚糖等方面的研究尚不足;因此,探索新来源的ISase,尤其是有较高转糖基能力的新型ISase,对拓展其应用方面有重要作用。3)蔗糖是ISase的反应底物。制糖工业的副产物,如糖蜜等,其蔗糖含量丰富。因此,以糖蜜为底物,合成菊糖或低聚糖等研究,符合资源综合利用和可持续发展的要求。
猜你喜欢
企业文明(2023年5期)2023-09-30 09:49:24
食品与生物技术学报(2022年1期)2023-01-11 09:11:11
中国纺织(2021年12期)2021-09-23 09:49:43
安徽化工(2021年2期)2021-05-15 13:41:22
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
食品工业科技(2019年15期)2019-08-28 12:37:10
食品工业科技(2018年19期)2018-10-22 09:33:32
郑州大学学报(理学版)(2017年1期)2017-04-07 01:29:23
物理化学学报(2015年7期)2015-12-30 12:13:08
首都医科大学学报(2015年4期)2015-12-16 13:00:08
