
胡萝卜薄片红外干燥光学特性研究
2021-05-19 01:50刘玉辉李腾训王相友魏忠彩
农业机械学报 2021年4期
刘玉辉 李腾训 王相友 魏忠彩
(山东理工大学农业工程与食品科学学院, 淄博 255049)
0 引言
红外辐射加热具有能量利用率高、干燥速率快、加热均匀、热惯性小等优点[1],在果蔬干燥领域应用越来越广泛[2]。目前,利用红外辐射对果蔬干燥的研究主要集中在提升干燥效率、降低能耗和提高干制品品质等方面[3-15],这些研究主要从宏观上探索如何利用红外光能优化果蔬干制品的品质和提高对红外辐射能的利用率,而对红外线与果蔬组织之间光学作用机理的研究较少,没有从微观上对红外线在物料中的反射、散射和吸收特征进行深入细致的分析,故无法从根本上揭示红外辐射干燥的内在机理。
红外辐射干燥是红外辐射能最终转化为物料的内能、并在物料内部进行传递与耗散的过程[16]。果蔬结构复杂,所含物质种类繁杂多样,很难通过构建数学模型的方式对红外线在果蔬中的传递特性进行准确的描述,对光与物料进行的干涉与衍射、偏振与色散等作用机理和作用效果也很难进行精准测量。本文基于红外线对果蔬的微观作用机理,研究分析红外辐射能转化为果蔬内能的机制。
胡萝卜含水率高、所含成分复杂,对不同波长的光具有不同的折光率,且其中所含种类繁多的原子以及复杂多变的化学键,对不同波段红外线的吸收以及红外线在物料中传输过程的阻尼状态,都具有复杂的形式[17]。本文选用胡萝卜为研究对象,通过测量与分析胡萝卜干燥过程中的外观品质、质构特征、光谱特性、超微结构、能耗、可溶性固形物含量以及含水率的变化规律,分析各指标与红外线之间的作用特性和关联性,从而探索红外线在物料中的传播规律和光学机理。
1 材料与方法
1.1 试验材料与设备
红森胡萝卜,购自山东省淄博市农贸市场,新鲜,品质优良。
YHG-500-BS型远红外恒温干燥箱,上海跃进医疗器械厂;DHG-0246A型电热恒温鼓风干燥箱,上海精宏实验仪器设备有限公司;TP-114型电子天平(量程220 g,精度0.1 mg),丹佛仪器北京有限公司;物料盘若干;样品厚度可调节刀具;镊子;手套;打孔器;与试验样品直径一致的模具;Quanta250型扫描电子显微镜,FEI香港有限公司;JEOL-1200EX型透射电子显微镜,日本JEOL公司;LKB-V型切片机,瑞典LKB公司;FW100型高速万能粉碎机,天津市泰斯特仪器有限公司;TGL-20M型高速冷冻离心机,湖南湘仪实验室仪器开发有限公司;TA.XT2i/50型质构仪,英国Stable Micro System公司;Labspec 4型全光谱仪(光谱范围350~2 500 nm;400~1 000 nm,分辨率3 nm;1 000~2 500 nm,分辨率6 nm;IRIS-L50型卤素灯(50 W石英卤素灯,可调光,光源可变高及角度);计算机等部件),美国ASD公司。
1.2 试验设计与方法
1.2.1湿基含水率
胡萝卜薄片的切割、红外干燥和湿基含水率测定方法均参照文献[18]。
1.2.2全光谱成像数据的采集与标定
开机,预热30 min,打开R3软件,确认联网;打开“Control”菜单的“Spectrum Save”选项,设置参数;打开“Control”菜单的“Adjust Configuration”选项,设置参数;将光谱仪探头对准白板,点击“OPT”图标优化光谱仪的积分时间,点击“WR”图标,仪器会自动重新采集暗电流,几秒钟之后界面上显示一条反射率为1的平直线,按空格键存储当前的光谱曲线。把光谱仪光纤探头瞄准胡萝卜样品的数据采集部位,此时界面上显示的就是被采集部位相对反射率光谱线,按空格键存储当前的光谱曲线。
1.2.3超微结构观察与测定
胡萝卜的微观结构观察与测定参照文献[19]。
1.2.4可溶性固形物含量测定
取2.5 g胡萝卜切碎加入5 mL离心管中,放入冷冻研磨机以2 000 r/min研磨10 min(12个样可以同时磨),将钢珠取出,倒入3 mL离心管,用石英砂配平,以8 000 r/min离心5 min,用吸管吸取上层悬浮液滴于手持折光仪镜片上,用手持折光仪读数并记录。
1.2.5质构测定
取干燥时间1.5 h无翻动的横切样品,用直径10 mm的打孔器在胡萝卜的髓部和皮层部打孔,形成圆柱体小丁,采用TA.XT2i/50型质构仪,HDP/BSK型探头,设置探头测试前、测试中、测试后的速度分别为2.0、1.0、1.0 mm/s,记录测试过程中的时间-应力曲线。对样品进行全质构分析,触发力5 kg,每组试样重复测定5次。胡萝卜的硬度用第1次压缩变形30%的峰值力(N)表示[20]。
1.2.6干燥能耗
将胡萝卜横向切成厚度3.5 mm的薄片,取样550 g,用红外辐射加热系统(物料表面温度分别为50、55、60、65、70、75、80、85℃,辐照距离30 cm,功率1 200 W)对物料进行干燥。每隔30 min(含水率降至20%后,每隔15 min)取样一次并称量,再将所取样品置于105℃热风恒温箱内,干燥至湿基含水率降至8.7%时,停止试验。每个试验重复3次。
胡萝卜单位能耗表征每蒸发胡萝卜内单位质量的水分需要消耗的电能[21],计算公式为
(1)
式中N——干燥能耗,MJ/kg
W——额定输入功率,kW
t——干燥时间,h
G——去除水分的质量,kg
2 结果与分析
2.1 干制品的外观品质
图1为在干燥温度60℃条件下物料在试验过程中的外观品质。
由图1a可以看出,在干燥过程中,每隔0.5 h对物料翻动1次,干物料平整、舒展,皱缩度低,未经翻动的样品髓部向上突出,皮层部向下突出。由图1b可以看出,热风干燥的物料褐变不明显,红外干燥的物料褐变度较高,且髓部的褐变度高于皮层部。由图1c可以看出,干物料的褐色在复水后褪去,说明褐变产物溶于水。由图1d可以看出,干燥过程中,物料髓部逐渐向上突出,皮层部逐渐向下凹陷,说明在同一时间点上,髓部下表面含水率下降快,收缩应力高于上表面,皮层部恰好相反,上表面含水率下降快,收缩应力高于下表面,在两表面不均衡应力的作用下,薄片发生扭曲变形。干燥时间为4 h时,出现明显褐变现象,测定此时物料的湿基含水率约为20%,具有典型的美拉德反应特征[22]。
2.2 干燥过程中物料的质构特征
取干燥时间1.5 h无翻动的物料,对髓部和皮层部分别做3组重复的全质构分析(TPA)试验,得表1所示数据。在TPA各项指标中,与含水率相关的硬度指标存在显著性差异。由于新鲜果蔬中含水率越高,硬度越大[8],因此,在干燥时间为1.5 h时,髓部上表面的含水率高于下表面,皮层部下表面的含水率高于上表面。

表1 红外干燥1.5 h时胡萝卜样品的TPA指标Tab.1 TPA index of carrot sample after infrared drying for 1.5 h
2.3 光谱特征
对样品进行全光谱数据采集,得到图2所示的反射率图谱。
由图2a可知,在含水率相同的条件下,髓部对任意波段光的反射率都低于皮层部,说明髓部和皮层部表面结构的差异对光反射率的影响比较明显;在相同部位,含水率越高,物料表面对任意波段光的反射率越低;物料对600~1 200 nm波段比对1 800~2 500 nm波段光的反射率高,说明短波光与长波光在物料表面的作用机理存在差异。由图2b可以看出,无论是髓部还是皮层部,横切样品在相同区域对光的反射率都高于纵切样品;无论是采用横切方式还是纵切方式,髓部对光的反射率都低于皮层部。
研究资料表明:水对980 nm波长光谱的吸收能力较其它物质敏感,对1 100、1 380、1 870 nm波长附近红外线有更高的吸收率系数[23]。从图2的反射率谱线可以看出,物料在950、1 200、1 400、1 800 nm附近的近红外波段存在吸收峰,这几个吸收峰的位置与水的吸收特征谱线具有较高的吻合度,因此可以认为胡萝卜内部成分对红外辐射吸收占主导作用的是水分。
2.4 超微结构
若研究物料与红外线的作用机制,需对物料的超微结构进行分析。在表面温度60℃条件下经红外干燥后的物料髓部和皮层部的扫描电子显微镜图像(SEM)和鲜物料透射电子显微镜(TEM)图像分别如图3、4所示。
由图3可以看出,相对于皮层部,髓部干样品孔隙壁平整度高,孔隙排列规则,结构松散,孔隙内部空间大,闭合孔隙数量少;髓部孔隙更好地保留了原生态的空间架构,孔隙形状大致呈方管或圆管状直通状态,孔隙迂曲度低,孔隙尺寸和比面都比较大,通透度高;与横切样品相比,纵切样品的表面平整,结构致密,仅在髓部有少量孔隙,皮层部孔隙结构不明显。
由图4可以看出,髓部细胞内细胞器较少且发育不完全,整个细胞膜内空间几乎被液泡占满,细胞器被细胞膜和液泡膜限定于紧邻细胞壁的狭小区域内,细胞在总体上呈水球形态;皮层部细胞内细胞器清晰可见,细胞器占空比较大,呈颗粒状,较小颗粒的直径约为0.1 μm,较大颗粒物直径在1 μm以上,细胞呈水球中掺杂大量杂质的形态。
2.5 可溶性固形物含量
尽管细胞结构复杂,内容物对光线的折射系数差异影响较大,不易对其进行精准测量,但水溶液所含溶质的浓度会影响溶液的折光率[24],由于胡萝卜所含物质除水外,主要是蛋白质、淀粉、葡萄糖、纤维素及其它碳水化合物,无机盐类物质含量较少,所以,胡萝卜中水溶液的溶质主要是以可溶性糖类为主的固形物。对新鲜胡萝卜的髓部和皮层部所含可溶性固形物质量分数进行测量,结果如表2所示。
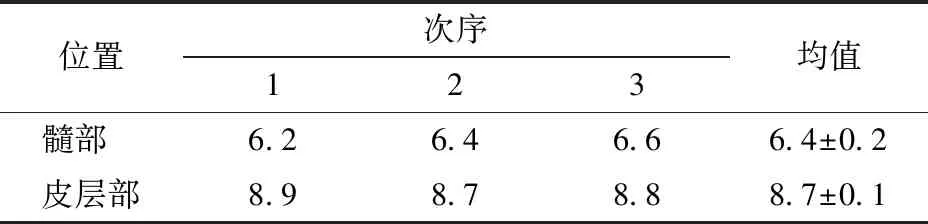
表2 髓部和皮层部可溶性固形物质量分数Tab.2 Mass fraction of soluble solids in medulla and cortex %
由表2中可溶性固形物质量分数差异可以看出,新鲜胡萝卜髓部的可溶性固形物质量分数低于皮层部,说明髓部的水溶液浓度低于皮层部。这与髓部细胞内液泡体积较大和细胞器数量较少、体积较小的特点相吻合。由于浓度高的液体具有较高的折光率,因此,皮层部比髓部对红外线的折光能力强。
2.6 干燥温度对单位能耗的影响
在功率、辐照距离和物料质量均相同的情况下,分别在表面绝对温度为323.15、328.15、333.15、338.15、343.15、348.15、353.15、358.15 K条件下对物料进行红外干燥,运用式(1)进行运算,得到表3所示的单位能耗与干燥温度的关系。
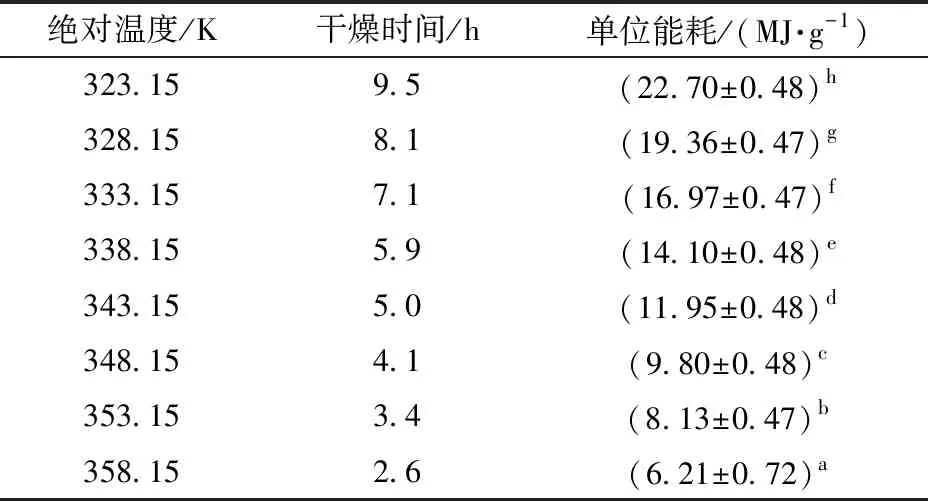
表3 单位能耗与干燥温度的关系Tab.3 Relationship between unit energy consumption and drying temperature
由表3可以看出,干燥温度越高,单位能耗越低,二者存在负相关关系。同时,干燥温度越高,差异越明显,说明随着干燥温度升高,干燥箱体温度升高吸收的能量、干燥箱向外部环境散失的能量均增大,致使测量误差随温度升高而增大。
由黑体辐射的光谱辐射出射度公式[25]可知光谱辐射出射度峰值与绝对温度的5次方成正比,公式为
Mbλm=BT5
(2)
式中Mbλm——辐射出射度峰值,W/(m2·μm)
B——常数,取1.286 7×10-11W/(m2·μm·K5)
T——绝对温度,K
由表3可以得出,红外干燥的单位能耗与物料表面绝对温度的5次方呈线性负相关,在50~85℃范围内,温度越高,干燥速率越快,单位能耗越低,与式(2)相吻合。
3 理论分析与讨论
3.1 散射的影响
根据胡萝卜细胞的结构特点,将细胞及细胞内颗粒物均抽象为球形模型,将红外光场分解为入射场、散射场和球内场,则散射问题可以归结为散射体(细胞及细胞内颗粒状物质)对入射光的衍射、入射光在散射体表面的反射以及光线进入散射体内部形成的折射3部分[24]。
当入射光波长与散射体直径相近时,满足米氏散射条件。根据米氏散射理论,散射光强与入射光强、散射体的关系[26-27]为
(3)
式中I(θ)——散射光强,cd
I0——入射光强,cd
S1(θ)、S2(θ)——散射光的振幅函数
θ——散射角,rad
r——光源与散射体的距离,μm
λ——入射光波长,μm
从式(3)可以看出,米氏散射的散射强度与频率的2次方成正比,前向散射光强度高于后向,后向散射光强度高于侧向,方向性特征明显[28-29];入射光波长越小,散射光能量越集中分布在散射角较小的范围内,相对折射率的变化对散射光分布的影响不大,不同半径散射体的散射光强的分布差异较大[24]。
由图4可知,尽管胡萝卜细胞大小不等,但总体上细胞直径在10 μm左右,细胞内大颗粒物的尺寸在1 μm左右。当入射光波长位于100~1 000 nm之间时,尽管在胡萝卜皮层部的散射具有较强的前向性,但后向的强度同样不容忽视,而该波段光线在照射髓部时,由于缺少散射必需的颗粒物作散射体,因此,该波段光线在髓部的米氏散射较弱,这是图2a光谱曲线上皮层部比髓部在100~1 000 nm波段反射率高的主要原因。
当入射光波长达到甚至超过颗粒物直径的10倍时,由于颗粒物的直径远小于波长,散射体内的球内场在相位上与球外场的区别可以忽略,光波通过散射颗粒所需要的时间积累可以忽略,散射微粒形成电偶极子,振荡频率与入射光相同,所发射子波即为散射波,此时,米氏散射可近似为瑞利散射,近似散射光强分布[24]为
(4)
式中a——细胞器颗粒半径,μm
m——细胞器颗粒与细胞质液体之间的相对折射率
由式(4)可以看出,瑞利散射光强的角分布特征对所有大小和折射率的颗粒都相同,前向散射与后向散射呈对称性;散射光强与颗粒粒径的6次方成正比;散射光强与入射波长4次方成反比。
对照图4可知,波长在1~10 μm范围内的红外线在物料中会存在米氏散射,波长在10~100 μm范围内的红外线在物料中主要表现为瑞利散射。由于皮层部颗粒物数量多,因此,波长在1~10 μm之间的光在皮层部的瑞利散射比髓部明显,表现为图2a光谱曲线上皮层部比髓部对1 μm以上波段光反射率高。
由于瑞利散射在不同方向上均有散射光线,因此,在垂直于光的传播方向上的散射光仍然具有较强的能量。因此,两种散射方式相比,发生在长波段的瑞利散射作用效果更明显,表现为散射光的大部分散射能量最终会被皮层部上表面附近的物料吸收,转化为该部位物料的内能,皮层部上层获得的能量高于下层。当上层水分与物料分离的速率高于下层水分向上传递的速率时,下层含水率就会高于上层,表现为表1中皮层部下表面硬度高于上表面。
由于髓部颗粒物含量较少,两种散射的影响都较弱,红外线在髓部表现出更强的穿透性。在水分迁移过程中,当上层水的散失量与下层水的输入量之差小于下层水向上的迁移量时,下层的含水率就低于上层,此时表现为上表面的硬度高于下表面。
3.2 水对红外线的吸收作用
3.2.1水的光谱吸收特征
由于新鲜胡萝卜的湿基含水率高达89%~91%[30],需要分析红外线与水作用的机理,充分发挥水对红外线的广谱吸收作用,提高胡萝卜干燥效率和提升干制品的品质。文献[31]发现在3 900~3 600 cm-1(伸缩振动)和1 800~1 400 cm-1(弯曲振动)区间,水对垂直偏振光吸收较强,对水平偏振光吸收较弱。文献[32-33]发现水和离子溶液在低频范围内的特征峰包括60 cm-1氢键弯曲、200 cm-1氢键伸缩和600 cm-1分子摆动,在液体状况下这些氢键还以皮秒量级的速度不停地断裂-形成,与此同时,水分子在液体状态下还以皮秒量级的速度在旋转,此外氢原子的量子效应也在其中发挥着作用。由图5[25]可以看出,除去300~900 nm波段的光存在一个吸收低谷外,其余波段的光波均能够被水强烈吸收。在水中大量氢键的作用下,水的比热容较大,气化热值较高,物料在脱除自由水阶段会吸收大量的能量而温度保持相对稳定,因此,在干燥前期可采用较高的辐射功率,既能提高干燥速率,又可避免营养物质因温度过高而被破坏。
3.2.2水的活性与美拉德反应
胡萝卜中的水分子是以团簇的形式存在的,团簇的大小与水中溶解的溶质颗粒大小和分子极性有关。由表2可知,胡萝卜所含可溶性固形物较多,胡萝卜中的水分子与溶液中的有机溶质相互作用时,在数量庞大的弱氢键作用下,能够形成活性较低的水化层结构,溶解的颗粒在表面不饱和键力或极性作用的影响下,吸引偶极水分子,使极性水分子在颗粒表面形成定向、密集、有序排列的水分子层[34-35],这就极大地降低了水的活性。红外线可以通过破坏水分子氢键的方式,减少水分间的缔合力,使大的水分子团簇分解为小的水分子团簇[36],提高水的活性。
影响美拉德反应的因素除去物料自身的因素之外,还有温度、反应时间、水分活度和pH值、金属离子和化学试剂等外界因子[37]。试验发现,采用相同的物料,在相同的环境温度、湿度、干燥温度和干燥时间条件下,经红外干燥的样品,非酶褐变的程度高于热风干燥,可见,造成美拉德反应加剧的主要原因是红外辐射增加了水的活性。美拉德反应产物具有抗氧化活性,但是其抗氧化能力受多重因素的影响[38-39],另外,尽管美拉德反应的产物有一定的营养价值,但目前对胡萝卜干制品的利用均采用复水后再使用的方式,这就降低了褐变产物的利用率,同时美拉德反应消耗了物料中原有的营养成分,造成了营养流失,降低了胡萝卜干制品的外观品质。由于美拉德反应程度与温度存在正相关关系[37],因此,实际生产中,可以通过降低干燥温度的方式对美拉德反应进行抑制。
3.2.3细胞组织结构对水的影响
胡萝卜具有植物根的基本特征,如图6所示,组织细胞沿植物生长方向伸长[40],这种结构与图3中呈现的横切样品孔隙数量多、深度大的特征相吻合。红外线穿过横切薄片的每层细胞时通过的距离要大于纵切薄片。因此,厚度相同的条件下,纵切薄片比横切薄片层级数量多。层级数量的增加,既降低了红外线与水直接作用的几率,又降低了水的迁移能力。
文献[18]研究发现,胡萝卜在干燥失水过程中,细胞内容物会以固态颗粒物的形式附着于横向的细胞壁上,阻塞该部位细胞壁上的孔隙,降低了细胞的横向通透性,在失水过程中,细胞与邻近细胞之间会发生一系列生物化学反应,形成纵向通透、横向阻隔的网状空间架构。因此,无论是在干燥初始状态,还是干燥中间阶段,横切薄片更有利于对红外辐射能的吸收利用。
由图3a、3b可以看出,在横切方式下,细胞在切面处被截成开放的微小腔体,垂直入射的红外波进入该微腔后易形成谐振特征明显的相干光场,相干光场产生的光学力导致约束它的腔体产生微小形变,在一定条件下的腔体形变会导致空腔边界产生持续不断的机械振动[41],微腔的振动对腔内水分向外迁移形成助推作用,振动产生的机械能又会迅速转化为物料的内能,使物料温度快速上升,腔内水分子运动速率加快,水的迁移能力增强。同时,由于细胞形成的微腔结构对光场具有束缚作用,使得腔内光场因具有极高的功率密度而产生级联混频效应,在腔内产生一系列宽带的梳状光谱[42],提高了光能向内能转变的速率,使空腔中水的迁移能力迅速提高。空腔结构对光的作用表现为:粗糙多孔的横切面比光滑平整的纵切面在图2b谱线上呈现出较低的反射率。表面空腔快速脱水后对物料内部相邻空腔形成负压,提高了相邻空腔内水分向外迁移的能力,相邻腔体的脱水速率增大,逐层依次向内传递,形成由表及里的连锁效应,使物料内部水分在整体上向表面迁移的速率加快。
3.2.4水溶液浓度对光吸收的影响
水溶液浓度越高,对水的折射作用越明显[43-44],由表2可知,胡萝卜皮层部可溶性固形物质量分数高于髓部。因此,皮层部具有更高的折光率,红外线自物料上表面射入后,光入射到皮层部后会产生更大的偏转角,降低了光传播到下表面的几率,使下表面能够接收到的辐射能量降低。因此,与髓部相比,物料皮层部下表面水的迁移能力较弱。
4 结论
(1)采用横切方式既能增大胡萝卜薄片中水与红外线直接作用的几率,发挥孔隙对红外线的吸收能力,削弱红外线在物料表面的散射作用,提升物料对红外辐射吸收的均匀度,又可减少薄片的层级数,降低水的迁移阻力。
(2)在干燥前期应采用较高的辐射功率,以提高自由水的脱除速率,并降低能耗;在干燥后期应采用较低的温度,对美拉德反应进行抑制。
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
中南林业科技大学学报(2022年7期)2022-09-26
医学概论(2022年3期)2022-04-24
农业科技与信息(2021年24期)2022-01-05
首都体育学院学报(2019年5期)2019-10-18
发明与创新·小学生(2018年7期)2018-08-06
中国当代医药(2017年17期)2017-07-25
支部建设(2016年18期)2016-11-28
烟草科技(2011年12期)2011-01-16
东北林业大学学报(2010年6期)2010-08-08
