
香稻品种OsBADH2突变位点分析及其功能标记的开发
2021-05-14 06:01:16孙平勇张武汉舒服何强张莉彭志荣邓华凤
生物技术通报 2021年4期
孙平勇 张武汉 舒服 何强 张莉 彭志荣 邓华凤,3
(1. 湖南杂交水稻研究中心杂交水稻国家重点实验室,长沙 410125;2. 湖南省核农学与航天育种研究所,长沙 410125;3. 湖南省农业科学院,长沙 410125)
水稻是重要的粮食作物之一,杂交水稻的发展为解决我国人民的温饱问题作出了巨大贡献。随着国家经济的飞速发展、农业生产力的不断提升,我国居民实现了由过去吃不饱到现在吃得好的历史性转变。香稻不仅具有浓郁的香味,而且富含多种维生素和芳香氨基酸,有很高的营养价值[1],深得国内外消费者的喜爱。传统的香稻品种产量低,导致香米的价格是普通大米的2-3倍[2-3]。因此,香味作为水稻的重要品质性状,越来越受到育种专家的重视。明确水稻香味遗传的分子机理有助于培育香稻新品种。
近年来,测序技术、基因编辑和分子生物学的飞快发展,促进了水稻香味基因的克隆及其分子机理的解析。水稻OsBADH2(LOC_Os08g32870)是一个重要的香味调控基因。早在1992年,康奈尔大学的Tanksley团队将香味隐性基因OsBADH2初步定位在水稻第8染色体上,与RFLP标记RG28的遗传距离为4.5 cM[4],后来通过不同的遗传群体将其定位在标记RG28的附近[5-7]。直到2005年,澳大利亚Bradbury才将OsBADH2精细定位在SSR标记RM515和SSRJ07之间386 kb的区间,并通过测序分析成功预测到候选基因[8]。OsBADH2的编码蛋白为甜菜碱醛脱氢酶,香稻OsBADH2第7外显子存在8 bp的缺失和3 bp的替换,导致其蛋白功能丧失,从而使籽粒积累香味物质2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline,2AP)[8]。功能互补和 RNAi等转基因方法证实了OsBADH2的功能[9-10]。研究表明,OsBADH2第7外显子的8 bp缺失是主要的变异类型[11],之后在第 1、2、10、12、13和 14外显子以及第4和第5外显子之间、第1外显子和第1内含子的剪接位点、5′UTR区发现一系列新的等位突变类型[3,12-17]。这些等位基因的发现,为培育优质香稻品种提供了重要的基因资源。
目前,OsBADH2不同的等位变异相继被报道,但通过质谱仪定量比较它们之间香味表型差异的甚少。本研究通过对10份香稻OsBADH2编码区进行测序,发现其中9份香稻第7外显子缺失8 bp和替换3 bp、1份香稻第14外显子插入1个碱基G。对香味物质2AP含量进行测定,表明OsBADH2不同变异类型的香味有强弱之分。根据第7外显子的变异开发功能标记E7,并对香稻资源和育种群体进行鉴定,为利用OsBADH2培育优质香稻新品种奠定基础。
1 材料与方法
1.1 材料
香稻品种(品系)10份:巴斯马蒂、香杂1号、玉油香占、象牙香占、花香B、农香24、稻花香、香粳、明香10S和xiao7。非香稻品种(品系)2份:魔王谷和02428。香稻育种群体:泰丰B/明香10S//泰丰B和粳944/稻花香//粳944。
供试材料于2018年和2020年正季种植在湖南杂交水稻研究中心试验田。
1.2 方法
1.2.1 DNA提取、PCR扩增和电泳检测 采用CTAB法提取水稻叶片的基因组DNA。利用擎科生物公司的T3 Super Mix体系进行PCR扩增:98℃ 2 min ;98℃ 10 s,55℃ 10 s,72℃ 20 s,35 个循环 ;72℃ 2 min;4℃冷却。反应产物利用1.5%的琼脂糖凝胶检测;或者通过8%的聚丙烯酰胺电泳检测,PCR产物送擎科生物公司测序。扩增OsBADH2编码区的9对引物序列参考Kovach等[11]。
1.2.2 OsBADH2编码区的比对分析 以野生型非香稻日本晴OsBADH2编码区的序列为参考,其全长下载自RGAP数据库(http://rice.plantbiology.msu.edu/)。通过Sequencher软件对测序结果进行比对分析,最终获得供试香稻OsBADH2编码区的变异类型。
1.2.3 香味物质2AP含量的测定 委托武汉普奈斯生物科技有限公司测定香味物质2AP的含量。取水稻成熟籽粒并用干冰保存,种子去壳后,于液氮中研磨粉碎,以1∶1的无水乙醇和氯仿萃取样品中的2AP,通过Thermo TSQ 8000 EVO质谱仪进行测定。每个材料设3次生物学重复,通过t测验计算P值分析品种间的显著性差异。
1.2.4 OsBADH2功能标记的设计与合成 根据香稻OsBADH2第7外显子8 bp的缺失,通过Primer Premier5软件设计1对能检测水稻是否具有香味的功能标记E7,其引物序列为E7F:5′-ACCCCATCAATGGAAATG-3′,E7R:5′-CAGGATAGAACTGGCTAC-3′。引物由擎科生物公司合成。
2 结果
2.1 香稻OsBADH2变异类型的分析
为了明确供试香稻的香味是否由OsBADH2的变异造成,对其编码区进行测序分析。利用9对覆盖OsBADH2的引物对样品DNA进行PCR扩增,其产物通过琼脂糖电泳检测。以农香24和稻花香的扩增结果为例,除了第4对引物的扩增产物有非特异性条带,其他产物均为符合目标大小的单一清晰条带(图1),因此适合测序。
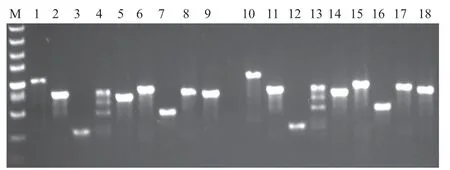
图1 OsBADH2 编码区的扩增Fig. 1 Amplification result from coding regions of OsBADH2
测序结果显示,巴斯马蒂、香杂1号、玉油香占、象牙香占、花香B、农香24、稻花香、香粳和明香10S这9个香稻OsBADH2的第7外显子均有8 bp的缺失和3 bp的替换。香稻xiao7的OsBADH2第7外显子与非香稻日本晴的一致,而在第14外显子有1个碱基(G)的插入突变(图2)。
2.2 基于第7外显子8 bp的缺失开发OsBADH2的功能标记
为了通过分子标记辅助选择提高育种效率、培育香稻品种,根据OsBADH2第7外显子的变异开发了功能标记E7,E7的PCR扩增产物覆盖了8 bp的缺失位点(图3)。根据设计原理,利用E7对香稻(有8 bp的缺失)和非香稻的DNA进行扩增时,其产物大小分别为267和275 bp;如果是杂合型,扩增产物则有267和275 bp条带。因此,只需通过1次PCR扩增和电泳检测,即可快速鉴定出OsBADH2的3种基因型。
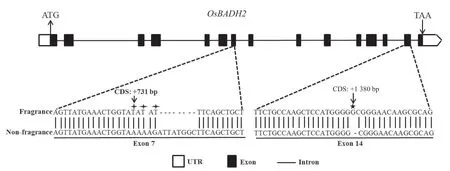
图2 OsBADH2 的基因结构和香稻的变异类型Fig. 2 Genetic structure of OsBADH2 and allelic variation types in fragrant rice

图3 功能标记E7 在OsBADH2 上的位置Fig.3 Location of functional marker E7 in the OsBADH2
为了验证功能标记E7的准确性,以测序的10份香稻和2份非香稻的基因组DNA为模板进行PCR扩增,并通过8%的聚丙烯酰胺电泳检测。结果显示,在第7外显子有8 bp缺失的香稻巴斯马蒂、香杂1号、玉油香占、象牙香占、花香B、农香24、稻花香、香粳和明香10S的扩增产物大小为267 bp;香稻xiao7在第7外显子没有8 bp的缺失,因此,它的扩增大小为275 bp,与非香稻魔王谷和02428的一致(图4)。PCR扩增结果与预测的结果完全一致,说明开发的功能标记E7能高效检测OsBADH2第7外显子是否有8 bp的缺失。
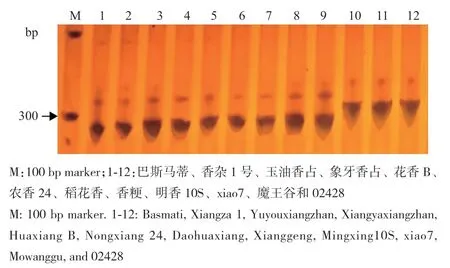
图 4 功能标记E7 对不同品种OsBADH2 基因型的鉴定Fig. 4 Molecular detection of OsBADH2 genotypes by functional marker E7 in different rice varieties
2.3 不同香稻香味物质2AP含量的比较
为了明确不同等位基因型之间香味物质的含量是否有差异,对OsBADH2第7外显子变异(象牙香占)和第14外显子变异(xiao7)的香稻以及非香稻(魔王谷)的香味物质2AP进行测定。结果表明,非香稻魔王谷的2AP含量最低,只有13.08 ng/g;香稻象牙香占的2AP含量适中,浓度为109.95 ng/g;香稻xiao7的2AP含量最高,达147.68 ng/g。香稻象牙香占和xiao7的2AP含量均显著高于非香稻魔王谷2AP的含量,而xiao7的2AP含量显著高于象牙香占的含量(图5)。
2.4 功能标记E7在培育香稻新品种中的应用

图5 不同品种香味物质2AP含量比较Fig. 5 Content of 2AP in different rice varieties
为了利用功能标记E7辅助选择培育优质香稻新品种,以香稻稻花香和明香10S为香味基因的供体,分别与粳944和泰丰B杂交构建了粳944/稻花香//粳944和泰丰B/明香10S//泰丰B群体。通过功能标记E7对这两份育种群体的OsBADH2基因型进行鉴定,结果显示,稻花香和明香10S的PCR扩增产物大小为267 bp,非香稻粳944和泰丰B的产物大小为275 bp,而杂合型扩增产物包含267和275 bp条带(图6)。基因型统计结果表明,粳944/稻花香//粳944群体的74个单株里有38个杂合,其余为非香型纯合,杂合数与纯合数之比接近1∶1;泰丰B/明香10S//泰丰B群体的41个单株中间有21个杂合,杂合数与纯合数之比也接近1∶1,与预期的分离比一致。因此,设计的功能标记E7可以对OsBADH2非香型显性纯合、香型隐性纯合和杂合3种基因型进行快速准确鉴定。
3 讨论
香稻资源遍布全球,泰国的茉莉香105(KDML105)和印度的巴斯玛蒂(Basmati)都是国际上有名的香稻。我国香稻的种植历史也很悠久,玉针香、螃蟹谷、香禾和靖西香糯分别是湖南、云南、贵州和广西的代表性常规香稻品种。传统的常规香稻因抗性差和植株偏高等原因导致产量低,而杂交香稻具有抗逆性强和产量高的特点[18]。1995年周坤炉[19]利用香稻不育系和非香稻恢复系育成了我国首个杂交香稻组合香优63。由于现有的香味基因属于隐性遗传,因此由香稻不育系和非香稻恢复系配组得到的杂交稻米粒只有1/4具有香味[20]。直到2013年,汤俭民等[21]利用均具香味的不育系和恢复系培育出一个全香型杂交稻红香优68。

图6 功能标记E7 对2 份育种群体OsBADH2 基因型的鉴定Fig. 6 Molecular detection of OsBADH2 genotypes by functional marker E7 in two breeding populations
快速准确地对香味性状进行鉴定是开展香稻育种的前提。传统育种一般通过咀嚼法和KOH浸泡法来鉴定香味[22],由于香稻的香味受环境条件的影响很大,加之人为因素,增加了其鉴定的难度[23]。此外,由于杂合体不具有香味,以往许多含有香味基因的优良杂合体被淘汰,不利于培育优质香稻新品种。因此急需一种简便、高效和准确鉴定香味基因型的方法。近年来,随着功能基因组学和测序技术的发展,分子标记辅助选择已广泛应用于香稻的遗传育种,大大提高了选择效率、加快了育种进程,许多香稻品种已得到大面积推广[24]。研究人员根据OsBADH2第12外显子3 bp的插入突变,设计了功能标记FME12-3,并利用2 个F2群体证明了FME12-3可以准确快速地鉴定OsBADH2的不同基因型[15]。沈雨民等[25]通过分子标记辅助选择培育出米质优、抗稻瘟病、异交率高和配合力好的不育系赣莲A,具有很好的应用前景。
利用分子标记辅助选育香稻新品种,首先要明确供体香味基因的突变类型,然后根据突变位点开发相应的功能标记。本研究对10份香稻OsBADH2编码区进行测序,发现其中9个香稻OsBADH2的第7外显子均为8 bp的缺失和3 bp的替换,只有1份香稻在第14外显子有1个碱基(G)的插入突变,这两种变异均为以前报道的类型[8,15]。研究表明第7外显子的缺失为主要突变类型,Kovach等[11]在鉴定的117个香稻资源中,有93份在OsBADH2第7外显子缺失8 bp。杨国峰等[26]对12个香稻进行测序,发现其中10个材料在第7外显子具有同样的突变类型。张江丽等[27]鉴定出86份具有香味的水稻资源,其中80份在第7外显子存在缺失。为了明确第7外显子的变异与其他类型的变异是否会导致香味表型的强弱差别,对第7外显子变异(象牙香占)和第14外显子变异(xiao7)的香稻以及非香稻(魔王谷)的香味物质2AP进行测定。结果表明,香稻象牙香占和xiao7的2AP含量均显著高于非香稻魔王谷2AP的含量,而xiao7的2AP含量显著高于象牙香占的含量。这一结果说明不同变异类型香稻的香味表型有强弱之分,同时可以推测第7外显子的突变作为应用最广泛的等位基因型,不是由于该等位基因型的效应最大(即香味物质的积累最多),而是因为这些香味品种具有相同的OsBADH2来源,在育种的过程中被大量选择保留。为了通过分子标记辅助选择提高育种效率、培育香稻品种,本研究根据OsBADH2第7外显子8 bp的缺失开发了功能标记E7,通过对香稻资源和育种群体进行鉴定,证明E7可以对OsBADH2非香型显性纯合、香型隐性纯合和杂合3种基因型进行快速准确鉴定,为利用OsBADH2进行分子育种、培育优质香稻新品种奠定了重要基础。
4 结论
象牙香占、花香B、农香24等9份香稻均在第7外显子发生8 bp的缺失和3 bp的替换,只有香稻xiao7在第14外显子有1个碱基(G)的插入突变。根据香稻OsBADH2第7外显子8 bp的缺失设计的功能标记E7,能快速准确鉴定OsBADH2的纯合显性、纯合隐性和杂合3种基因型。
猜你喜欢
农业与技术(2024年7期)2024-04-19 00:54:46
农业科技通讯(2023年1期)2023-02-12 07:09:58
电子科技大学学报(2022年5期)2022-10-29 01:57:52
福建稻麦科技(2022年3期)2022-09-14 02:17:10
今日农业(2021年18期)2021-11-26 00:54:08
小读者(2021年2期)2021-03-29 05:03:38
中国生殖健康(2020年4期)2021-01-18 02:58:10
文苑(2020年11期)2021-01-04 01:53:04
中国生殖健康(2018年4期)2018-11-06 07:12:16
热带农业科学(2015年8期)2015-09-10 12:54:53
