
4种鬼伞β-1 3-葡聚糖转移酶的生物信息学分析
2021-05-06 03:16牛鑫柴倩囡冯九海
江苏农业科学 2021年6期
牛鑫 柴倩囡 冯九海

摘要:鬼伞是一类菌柄能在短时间内快速伸长并且伞盖易自溶形成墨汁的蘑菇,菌柄在伸长过程中细胞壁以伸长生长为主,细胞壁组分β-1,3-葡聚糖发生重构修饰,GH72家族的β-1,3-葡聚糖转移酶能够将较低聚合度的寡糖转化生成更高聚合度的糖链。β-1,3-葡聚糖转移酶可能参与鬼伞菌柄细胞壁中β-葡聚糖组分的重构修饰。以完成测序并有注释信息的灰盖拟鬼伞(Coprinopsis cinerea)、拟鬼伞(Coprinopsis marcescibilis)、晶粒小鬼伞(Coprinellus micaceus)和小脆柄菇(Psathyrella aberdarensis)等4种鬼伞的β-1,3-葡聚糖转移酶的氨基酸序列为基础,采用TargetP、WOLF PSORT、SignalP、Sompa、TMHMM 2.0、Big-PI Fungal Predictor、MEME和SISS-MODEL等生物信息学分析工具对其开展蛋白质亚细胞定位、细胞信号肽、二级结构、跨膜螺旋、糖基化磷脂酰肌醇(GPI)锚定位点、基序以及三级结构等进行分析,同时对上述序列开展遗传进化关系分析。研究结果可为进一步深入开展该蛋白在菌柄伸长过程中细胞壁β-葡聚糖组分的重构修饰研究作基础。
关键词:真菌细胞壁;菌柄伸长;β-1,3-葡聚糖;细胞壁重构;多序列比对;生物信息学
鬼伞(coprinoid mushrooms)是一类子实体形成后,菌柄能快速伸长,伞盖易自溶形成墨汁的大型担子菌伞菌[1]。旧的分类系统中鬼伞科(Coprinaceae)包含鬼伞属(Coprinus)、小脆柄菇属(Psathyrella)和斑褶菇属(Panaeolus),依据第10版《真菌词典》最新分子系统学研究表明,鬼伞属已移至蘑菇科(Agaricaceae),小脆柄菇提升到科分类级别,旧的鬼伞属中大部分成员被划分至拟鬼伞属(Coprinopsis)、小鬼伞属(Coprinellus)和近地伞属(Parasola),其与小脆柄菇属(Psathyrella)等属组成小脆柄菇科(Psathyrellaceae),也就是现在的鬼伞科[2-4]。鬼伞菌柄的生长方式是一种类似植物胚芽鞘的顶端生长,其主要特征是细胞主要进行伸长生长,细胞基本不进行分裂[5]。蘑菇菌柄细胞壁的伸长生长理论经历了“水解酶模型”[6-7],“细胞膨压理论”[8]以及“水解酶-膨压折衷理论”[9]等,最新研究表明,特定的内切几丁质酶[10]和内切 β-1,3-葡聚糖酶均能引起灭活菌柄的伸长[11]。
真菌细胞壁主要由葡聚糖、糖蛋白和几丁质组成,对真菌的生存、生长和真菌细胞的形态至关重要[12-13]。尽管它是刚性结构,但它在细胞生长过程中具有可塑性,并能适应环境,因此,真菌细胞壁是一个动态变化的、有个组分相互交联在一起形成一个高强度的具有可塑性的三维复合结构[13]。有4个家族的糖基水解酶或者糖基转移酶已被证明在细胞壁组分的交联中发挥作用[12]。GH16家族水解酶(β-葡聚糖酶)已被证明可能参与葡聚糖和几丁质的交联[14-15]。GH17家族水解酶(β-葡聚糖酶)在葡聚糖之间的交联中起作用[16-17]。GH76家族的甘露聚糖酶在细胞壁的生物发生中起作用[18-19]。GH72家族葡聚糖转移酶已被证明是形成正常细胞壁必不可少的,能够与β-1,3-葡聚糖交联在一起。这些细胞壁重构酶中,尤其是GH72家族的β-1,3-葡聚糖转移酶为细胞壁提供了可塑性[20]。这些酶主要参与真菌细胞壁骨架结构多糖 β-1,3-葡聚糖的重构修饰,它们能够水解至少含10个葡萄糖单元的昆布寡糖中的β-1,3-糖苷键,并且把新形成的带还原性末端的供体(含有5个以上葡萄糖单元)转移至另一个β-1,3-寡聚糖的非还原性末端(受体)形成聚合度更高的寡糖。除 β-1,3-葡聚糖合成酶合成延长葡聚糖链外,这种转糖苷酶反应产生新的β-1,3糖苷键为β-1,3葡聚糖链的延长提供了另一种作用或者是协同作用机制[21]。迄今为止,这类酶在酿酒酵母(Saccharomyces cerevisiae)、粟酒裂殖酵母(Schizosaccharomyces pombe)、烟曲霉(Aspergillus fumigatus)、白色念珠菌(Candida albicans)等酵母和丝状真菌中具有广泛的研究,酿酒酵母中这类蛋白被命名为Gasp(表面锚定糖酯类蛋白),烟曲霉中为Gelp(葡聚糖延伸蛋白),白色念珠菌中则为Phrp(pH值响应蛋白)[22-23]。在酵母菌中,这些糖基转移酶在孢子和菌丝的细胞壁组装中具有重要作用,并在菌丝营养生长条件下维持细胞壁的完整性,是粟酒裂殖酵母生存不可缺少的[24],烟曲霉中编码Gel4蛋白的基因也是必需基因[21],但在大型傘菌中对该基因的研究较少。
本研究选取已完成基因组测序且有注释信息的4种鬼伞:灰盖拟鬼伞(Coprinopsis cinerea)、拟鬼伞(C. marcescibilis)、晶粒小鬼伞(Coprinellus micaceus)和小脆柄菇(Psathyrella aberdarensis),对其β-1,3-葡聚糖转移酶进行生物信息学和遗传进化分析。
1 材料与方法
1.1 试验材料
根据烟曲霉中已报道的β-1,3-葡聚糖转移酶Gel1蛋白序列(登录号:XP_749253.1),在美国国家生物技术信息中心(national center for biotechnology information,简称NCBI)数据库中的组装库(Assembly)中分别检索“Coprinopsis”“Coprinellus”“Psathyrella”,获得CC3、Copmar1、Copmic2和ASM412641v1等4个基因组组装数据,分别点击“BLAST the assembly”选项进入“BLAST”比对界面,点击切换至tblastn,将Gel1蛋白登录号XP_749253.1粘贴至文本框,点击BLAST,对期望值小于10-3的结果点击GenBank从Feature中查看CDS详情获得“protein_id”。
1.2 试验方法
1.2.1 蛋白信号肽预测 利用SignalP 5.0 (http://www.cbs.dtu.dk/services/SignalP/)在线预测蛋白的信号肽。
1.2.2 蛋白的基本理化性质预测 利用ExPASy中的ProtParam(http://web.expasy. org/protparam/)对蛋白的分子量、等电点、氨基酸组成、不稳定指数、脂肪族系数和总平均疏水指数等基本理化参数进行预测。
1.2.3 蛋白的亚细胞定位预测 利用WoLF PSORT(https://wolfpsort.hgc.jp/)进行蛋白的亚细胞定位预测。
1.2.4 蛋白二级结构预测 采用Sompa(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_ automat.pl? page=npsa_sopma.html)在线预测蛋白的二级结构。
1.2.5 蛋白质跨膜结构域预测 利用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)对蛋白质的跨膜区结构进行预测。
1.2.6 糖基化磷脂酰肌醇(GPI)修饰预测 利用big-PI Fungal Predictor(http://mendel.imp.ac.at/gpi/fungi _server.html)对蛋白的GPI修饰位点进行预测。
1.2.7 蛋白基序(motif)分析 采用MEME(http://meme-suite.org/tools/meme)对蛋白序列中的基序进行分析。
1.2.8 系统进化树构建 在NCBI中找寻Gel的同源序列,利用MEGA 7中的Clustal X进行多重比对分析,然后以邻接法构建系统进化树。
1.2.9 蛋白三级结构预测 利用SWISS-MODE(http://swissmodel.expasy.org/)对蛋白进行同源建模预测分析。
2 结果与分析
2.1 Gel蛋白序列及其信号肽、基本理化特性和亚细胞定位分析
通过烟曲霉Gel1对灰盖拟鬼伞、拟鬼伞、晶粒小鬼伞、小脆柄菇的基因组数据CC3、Copmar1、Copmic2和ASM412641v1进行tblastn比对,结果如表1所示。结果表明,灰盖拟鬼伞有2个Gel蛋白,拟鬼伞有4个Gel蛋白,晶粒小鬼伞和小脆柄菇各仅含有1个Gel蛋白。
由表1可知,各鬼伞Gel蛋白均含有信号肽,除晶粒小鬼伞和小脆柄菇的信号肽为N端前24个氨基酸,其他均为前20个氨基酸,结合WoLF PSORT 亚细胞定位分析,Gel蛋白均分泌到胞外。其氨基酸组成数量在520~550个之间,分子量为55 ku左右,等电点呈酸性,在4~5之间,除拟鬼伞的Gel(TFK21136.1和TFK21135.1)为不稳定蛋白,其他均为稳定蛋白,灰盖拟鬼伞Gel(XP_001831707.1)的脂肪族指数最低,为69.89,而晶粒小鬼伞Gel(TEB39055.1)的脂肪族指数最高,为87.09。除晶粒小鬼伞Gel为疏水蛋白外,其他均为亲水性蛋白。此4种鬼伞Gel蛋白的优势氨基酸均为丝氨酸(Ser)、丙氨酸(Ala)和甘氨酸(Gly)。
2.2 Gel蛋白的二级结构、跨膜螺旋以及GPI修饰位点预测
4种鬼伞的Gels蛋白中二级结构以无规则卷曲和α-螺旋为主,其次是延伸链和β-转角。α-螺旋比例在30%~36%之间,灰盖拟鬼伞Gel蛋白 XP_001831707.1 的最高,为35.28%,拟鬼伞中TFK30742.1的比例最低,为3096%;延伸链的占比在13%~17%之间,拟鬼伞中TFK30742.1的比例最高,为1655%,而TFK21138.1的最低,为1369%;β-转角占比最低,约为5%左右;无规则卷曲占比最高,在44%~50%之间。除拟鬼伞中TFK21138.1和小脆柄菇RXW23863.1在C端具有跨膜区域,其他Gel蛋白均不含跨膜结构。所有Gel蛋白均含有GPI修饰,主要修饰位点发生在C段区域的丝氨酸残基上(表2)。
2.3 Gel蛋白的基序分析、多序列比对及三级结构预测
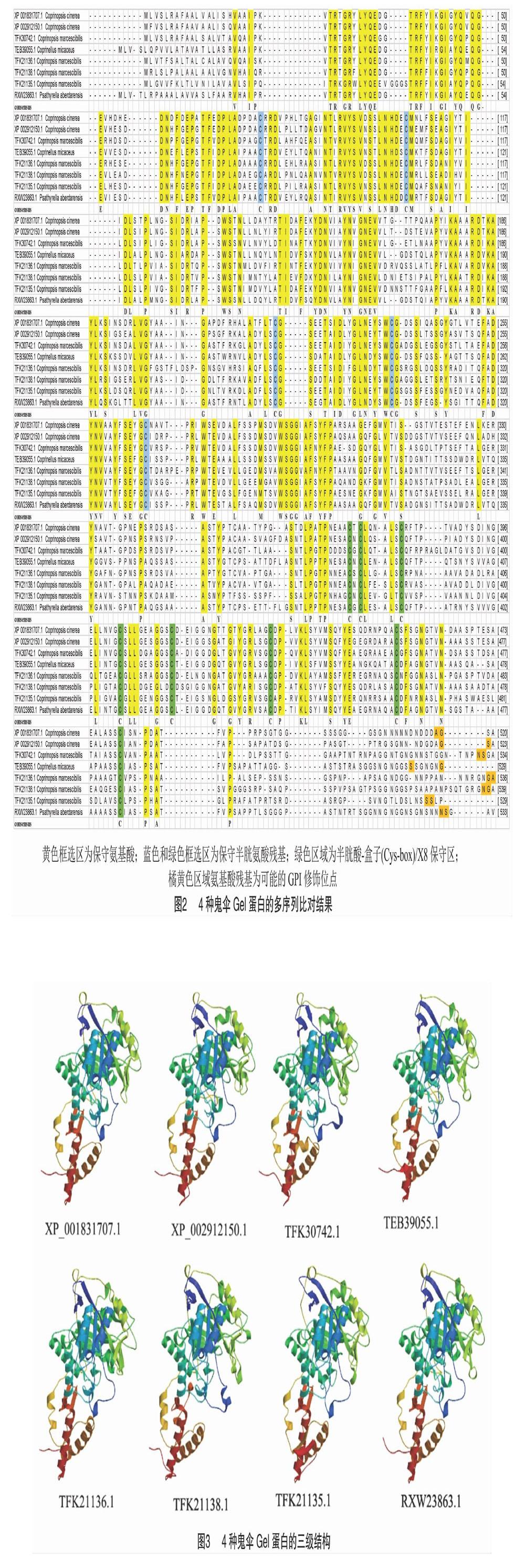
4种鬼伞的Gel蛋白序列中均含有3个基序(图1)。4种鬼伞Gel蛋白具有较高的同源性,同源性矩阵分析结果表明其同源性均在50%以上(表3)。晶粒小鬼伞和小脆柄菇的Gel蛋白同源性最高,将近80%;其次是灰盖拟鬼伞的2个Gel蛋白,其同源性为76.2%;晶粒小鬼伞的Gel与拟鬼伞的Gel(TFK21136.1)同源性最低,但也达到了51.6%。通过Gel蛋白的多序列比对(图2)可以看出,4种鬼伞的Gel蛋白中含有177个保守氨基酸,其中包含甘氨酸(G)和亮氨酸(L)各17个,15个酪氨酸(Y),14个丝氨酸(S),半胱氨酸(C)和脯氨酸(P)各13个,以及11个天冬酰胺(N)等。4种鬼伞的Gel 序列中均包含糖基水解酶超家族 (Glyco_hydrosuper family)和8个半胱酸-盒子(Cys-box/X8)保守域, 在C端的丝氨酸或者甘氨酸或者天冬酰胺上发生GPI修饰。使用酿酒酵母β-1,3-葡聚糖糖基转移酶GAS2的晶体结构(261.1.A)作为模板,采用Swiss-model對4种鬼伞Gel蛋白进行在线同源建模预测分析,结果表明预测结构与模板序列相似度均在35%左右,覆盖度约为80%,各鬼伞Gel蛋白的三级结构十分相似(图3)。
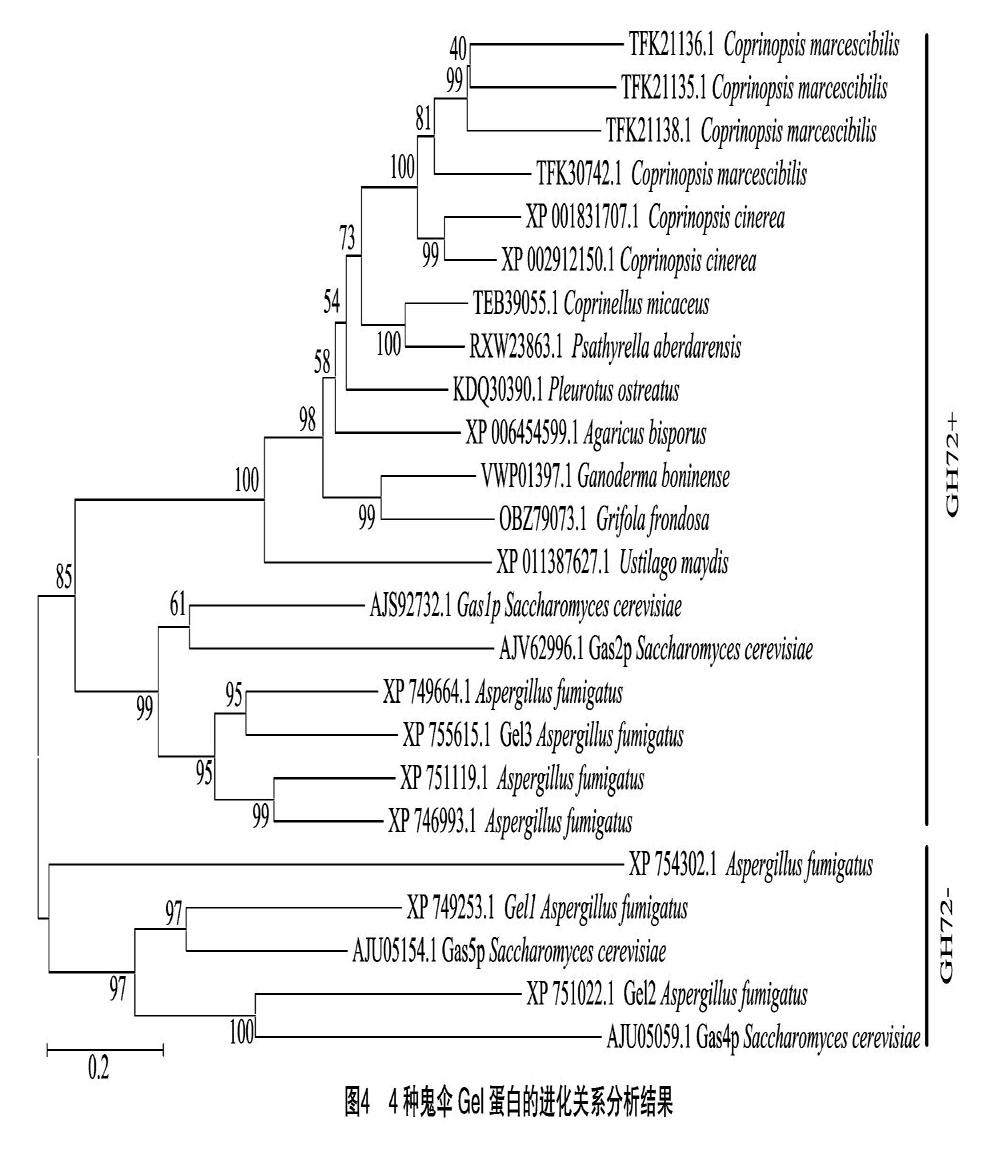
2.4 Gel蛋白的进化分析
结合研究较多的子囊菌中的烟曲霉(Aspergillus fumigatus)和酿酒酵母(Saccharomyces cerevisiae)中的β-1,3-葡聚糖糖基转移酶蛋白以及担子菌中的玉蜀黍黑粉菌(Ustilago maydis)和双孢蘑菇(Agaricus bisporus)、平菇(Pleurotus ostreatus)、灰树花(Grifola frondosa)以及灵芝(Ganoderma boninense)的Gel同源蛋白使用MEGA 7[25]进行演化分析,采用邻接(neighbor-joining)法推导其演化历史[26],得到分支长度和为8.760 563 63的最优树(图4)。Bootstrap 测试设置1 000次重复,进化距离采用基于JTT矩阵的方法[27]计算,以每个位点的氨基酸取代数为单位。分析包括24个氨基酸序列。所有包含间隙和缺失数据的位置被消除。最终数据集中共有347个位置。可以看出,4种鬼伞的Gel蛋白基本按照种属分类关系进行聚类,拟鬼伞属聚类为一组,小鬼伞属和小脆柄菇属聚为一组。根据序列C端是否有半胱氨酸富集结构域,GH72家族可分为2个亚类:GH72+(含CBM43或者X8)和GH72-(没有CBM43或者X8)[28]。所有担子菌的Gel蛋白序列中均含有X8结构域,因此属于GH72+。
3 討论
众所周知,GH72家族葡聚糖转移酶是GPI锚定修饰的细胞壁蛋白。该类蛋白在子囊菌中有较多研究,在酿酒酵母中含有5个GAS基因,白色念珠菌中含有5个PHR基因,粟酒裂殖酵母含有4个GAS基因,烟曲霉中含有7个GEL基因,稻瘟病菌中含有5个GEL基因,粗糙脉孢菌中含有5个GEL基因[29]。在本研究中除拟鬼伞外,大部分担子菌中仅有1~2个该类基因,明显少于子囊菌。因此,担子菌葡聚糖转移酶基因的生理功能可能不完全等同于子囊菌。GH72酶可分为2类[30],GH72+酶包含葡聚糖转移结构域和1个来自转移酶结构域的羧基末端第2个碳水化合物结合X8或CBM43结构域。GH72-酶缺乏X8/CB43碳水化合物结合域。4种鬼伞的Gel蛋白均属于GH72+,且均含有信号肽序列,因此,鬼伞的Gel可能在细胞周质空间中对新添加的细胞壁组分β-1,3-葡聚糖进行修饰。Hurtado-Guerrero等首次解析了GH72+与昆布五糖和昆布七糖水解产物有关的ScGas2转糖苷酶的晶体结构,结构表明存在1个(β/α)8的催化核心,紧紧地和C端CBM43葡聚糖绑定结构域相互作用,活性位点位于不常见的半胱氨酸富集的沟槽,有2个谷氨酸作为催化残基。定点突变数据结合ScGas2-o寡糖复合体的晶体结构,表明在受体位点产生绑定在调节水解和转糖苷之间的平衡至关重要[31]。该酶作用的位点包含2个催化残基(Glu176和Glu275),和3个酪氨酸残基(Tyr107、Tyr244、Tyr307),这些位点在GH72家族中均是保守的。去除GPI修饰位点该酶的转糖基活性不受影响,但去除X8结构域后其不再具有转糖基活性[30]。本研究基于已测序并有注释信息的4种鬼伞的Gel蛋白序列,通过对鬼伞Gel蛋白的生物信息学分析,为担子菌中Gel基因的生理功能研究奠定了基础。
参考文献:
[1]Kuo M. Coprinoid mushrooms:the inky caps[Z]. http://www.mushroomexpert.com/coprinoid.html.
[2]Kirk P M,Cannon P F,Minter D W,et al. Ainsworth & bisbys dictionary of the fungi[M]. 10th ed. Wallingford CABI Publishing, 2008:169.
[3]周茂新,文华安. 中国鬼伞属的研究现状[C]//第八届海峡两岸菌物学学术研讨会论文集.长春,2007:10-14.
[4]Redhead S A,Vilgalys R,Moncalvo J M,et al. Coprinus Pers. and the disposition of Coprinus species sensu lato[J]. Taxon,2001,50(1):203-241.
[5]Zhang W M,Wu X X,Zhou Y J,et al. Characterization of stipe elongation of the mushroom Coprinopsis cinerea[J]. Microbiology (Reading,England),2014,160(Pt 9):1893-1902.
[6]Kamada T,Takemaru T,Prosser J I,et al. Right and left handed helicity of chitin microfibrils in stipe cells in Coprinus cinereus[J]. Protoplasma,1991,165(1):64-70.
[7]Kamada T,Fujii T,Nakagawa T,et al. Changes in (1→3)-β-glucanase activities during stipe elongation in Coprinus cinereus[J]. Current Microbiology,1985,12(5):257-259.
[8]Mol P C,Vermeulen C A,Wessels J. Diffuse extension of hyphae in stipes of Agaricus bisporus may be based on a unique wall structure[J]. Mycological Research,1990,94(4):480-488.
[9]Bartnicki-García S. Glucans,walls,and morphogenesis:on the contributions of J. G. H. Wessels to the golden decades of fungal physiology and beyond[J]. Fungal Genetics and Biology,1999,27(2/3):119-127.
[10]Zhou J S,Kang L Q,Liu C C,et al. Chitinases play a key role in stipe cell wall extension in the mushroom Coprinopsis cinerea[J]. Applied and Environmental Microbiology,2019,85(15):519-532.
[11]Kang L Q,Zhou J S,Wang R,et al. Glucanase-induced stipe wall extension shows distinct differences from chitinase-induced stipe wall extension of Coprinopsis cinerea[J]. Applied and Environmental Microbiology,2019,85(21):1319-1345.
[12]Free S J. Fungal cell wall organization and biosynthesis[J]. Advances in Genetics,2013,81:33-82.
[13]Latgé J P. The cell wall:a carbohydrate armour for the fungal cell[J]. Molecular Microbiology,2007,66(2):279-290.
[14]Cabib E,Blanco N,Grau C,et al. Crh1p and Crh2p are required for the cross-linking of chitin to β(1-6)glucan in the Saccharomyces cerevisiae cell wall[J]. Molecular Microbiology,2007,63(3):921-935.
[15]Pardini G,De Groot P W,Coste A T,et al. The CRH family coding for cell wall glycosylphosphatidylinositol proteins with a predicted transglycosidase domain affects cell wall organization and virulence of Candida albicans[J]. The Journal of Biological Chemistry,2006,281(52):40399-40411.
[16]Goldman R C,Sullivan P A,Zakula D,et al. Kinetics of β-1,3 glucan interaction at the donor and acceptor sites of the fungal glucosyltransferase encoded by the BGL2 gene[J]. European Journal of Biochemistry,1995,227(1/2):372-378.
[17]Amandine G A,Mouyna I,Simenel C,et al. Characterization of a new β(1-3)-glucan branching activity of Aspergillus fumigatus[J]. The Journal of Biological Chemistry,2010,285(4):2386-2396.
[18]Kitagaki H,Ito K,Shimoi H. A temperature-sensitive dcw1 mutant of Saccharomyces cerevisiae is cell cycle arrested with small buds which have aberrant cell walls[J]. Eukaryotic Cell,2004,3(5):1297-1306.
[19]Maddi A,Fu C,Free S J. The Neurospora crassa dfg5 and dcw1 genes encode α-1,6-mannanases that function in the incorporation of glycoproteins into the cell wall[J]. PLoS One,2012,7(6):e38872.
[20]Cabib E,Arroyo J. How carbohydrates sculpt cells:chemical control of morphogenesis in the yeast cell wall[J]. Nature Reviews Microbiology,2013,11(9):648-655.
[21]Amandine G A,Fontaine T,Latgé J P,et al. β(1-3)glucanosyltransferase gel4p is essential for Aspergillus fumigatus[J]. Eukaryotic Cell,2010,9(8):1294-1298.
[22]Delso I,Valero-Gonzalez J,Gomollón-Bel F,et al. Inhibitors against fungal cell wall remodeling enzymes[J]. ChemMedChem,2018,13(2):128-132.
[23]Luo Z,Zhang T,Liu P,et al. The Beauveria bassiana gas3 β-glucanosyltransferase contributes to fungal adaptation to extreme alkaline conditions[J]. Applied and Environmental Microbiology,2018,84(15):1018-1086.
[24]Medina-Redondo M M,Arnáiz-Pita Y,Clavaud C,et al. β(1,3)-glucanosyltransferase activity is essential for cell wall integrity and viability of Schizosaccharomyces pombe[J]. PLoS One,2010,5(11):e14046.
[25]Kumar S,Stecher G,Tamura K. MEGA7:molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology and Evolution,2016,33(7):1870-1874.
[26]Saitou N,Nei M. The neighbor-joining method:a new method for reconstructing phylogenetic trees[J]. Molecular Biology and Evolution,1987,4(4):406-425.
[27]Jones D T,Taylor W R,Thornton J M. The rapid generation of mutation data matrices from protein sequences[J]. Computer Applications in the Biosciences,1992,8(3):275-282.
[28]Ragni E,Fontaine T,Gissi C,et al. The gas family of proteins of Saccharomyces cerevisiae:characterization and evolutionary analysis[J]. Yeast,2007,24(4):297-308.
[29]Kar B,Patel P,Ao J,et al. Neurospora crassa family GH72 glucanosyltransferases function to crosslink cell wall glycoprotein N-linked galactomannan to cell wall lichenin[J]. Fungal Genetics and Biology,2019,123:60-69.
[30]Popolo L,Degani G,Camilloni C,et al. The PHR family:the role of extracellular transglycosylases in shaping Candida albicans cells[J]. Journal of Fungi,2017,3(4):59.
[31]Hurtado-Guerrero R,Schüttelkopf A W,Mouyna I,et al. Molecular mechanisms of yeast cell wall glucan remodeling[J]. The Journal of Biological Chemistry,2009,284(13):8461-8469.楊艳北,许 晶,李袁飞,等. 沼泽红假单胞菌LuxR家族调控蛋白的生物信息学分析[J]. 江苏农业科学,2021,49(6):40-45.
猜你喜欢
浙江工业大学学报(2017年5期)2018-01-22
广东农业科学(2017年5期)2017-08-29
现代农业(2016年6期)2016-02-28
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
现代检验医学杂志(2014年3期)2014-02-02
食品科学(2013年13期)2013-03-11
军营文化天地(2010年1期)2010-04-04
