
长链非编码RNA MEG3对乳腺癌细胞侵袭及迁移的影响
2021-04-28 05:37张明亮张立功郭晨旭
赣南医学院学报 2021年3期
许 睿,钱 军,张明亮,张立功,郭晨旭,谢 强
(蚌埠医学院第一附属医院肿瘤外科-中德乳腺VIP病区,安徽 蚌埠 233000)
最新调查显示,乳腺癌的发病率在女性癌症中逐年上升,成为世界上最常见的癌症之一[1]。虽然近年来乳腺癌患者的总体生存和预后有所改善,但转移仍是患者死亡的首要原因。乳腺癌转移患者5年生存率仅26%,而非转移患者通过治疗后总生存率可以达到90%[2-3]。因此,确定乳腺癌转移的潜在分子机制和开发新的治疗靶点成为乳腺癌治疗的必要途径。
长链非编码RNA(Long non-coding RNA,lncRNA)是一组长度超过200 个核苷酸的非编码RNA[4]。在最近的研究中证实,lncRNA 在基因表达调控方面发挥着极其重要的作用,并参与了肿瘤的形成、增殖、侵袭、转移、耐药性等[5]。LncRNA MEG3 定位于人类染色体14q32,属于DLK1-MEG3 基因座[6]。最近几年引起了人们的广泛关注,已有学者表明lncRNA MEG3可作为抑癌基因调控癌细胞的增殖和转移[7],但在侵袭和迁移方面的机制并无具体研究。因此,本研究主要在细胞水平上通过在乳腺癌细胞系中过表达lncRNA MEG3,检测细胞迁移及侵袭能力的改变,并探讨其潜在机制。
1 材料与方法
1.1 细胞及主要试剂乳腺癌细胞MDA-MB-231、MCF-7、SKBR-3 均由中国科学院上海细胞库提供;pcDNA3.1-vector和pcDNA3.1-MEG3由上海吉凯基因公司合成;RPMI-1640 培养基及胎牛血清由美国Gibco 公 司 提 供;Lipofectamine 2000 转 染 试 剂、TRIzol®Plus RNA Purification Kit 和Revert Aid First Strand cDNA short kit 由美国Invitrogen 公司生产;Power SYBR®Green PCR Master Mix 由瑞士Roche 公司提供;兔抗人E-Cadherin、N-Cadherin、Vimentin 及β-actin 单克隆抗体及鼠抗兔IgG 二抗均购自英国Abcam 公司;结晶紫溶液及BCA 蛋白浓度测定试剂盒由上海碧云天公司生产;Transwell 小室购于美国Corning 公 司;matrigel 基 质胶 由 美 国Sigma 公 司 生产;LncRNA MEG3 引物序列由上海吉凯基因提供,引 物 序 列 如 下 : MEG3-F: 5'-ATCATCCGTCCACCTCCTTGTCTTC-3',MEG3-R:5'-GTATGAGCATAGCAAAGGTCAGGGC-3'miRNA-21-F:5'-TTTTGTTTTTGCTGGTCTTAG-3';miRNA-21-R:5'-AGCAGACAGTCAGGCAGGAT-3';GAPDH-F:5'-TGGTATCGTGGAAGGACTCAT-3',GAPDH-R:5'-GTGGGTGTCGCTGTTGAAGTC-3'。
1.2 乳腺癌细胞株RNA 提取及qRT-PCR 反应取相应的对数生长期乳腺癌细胞株,通过Trizol−氯仿−异丙醇法提取总RNA,继而逆转录成合成cDNA。以cDNA 为模板,进行PCR 扩增,GAPDH 为内参,采用2-△△Ct方法计算LncRNA MEG3的表达量。反应条件:①变性95 ℃25 s;②退火60 ℃25 s;③延伸72 ℃30 s。共40个循环。
1.3 细胞培养与转染人乳腺癌细胞株由10%胎牛血清及1%青链霉素溶液配置成的RPMI-1640 培养基进行培养,置于37 ℃,5%CO2的细胞培养箱中。当细胞处于对数生长期时,取适量细胞接种于六孔板中,按10 μL的Lipo2000™混合4 μg质粒置入500 μL不含血清的培养基中,细胞培养6 h 后更换不含双抗的10%胎牛血清培养基,48 h后继续进行实验。
1.4 Western blot 检测乳腺癌细胞中EMT 相关蛋白表达变化取对数生长期乳腺癌细胞,加入裂解液,提取总蛋白,BCA 试剂盒进行蛋白定量。以每孔15~20 μg 总蛋白量进行SDS-PAGE 凝胶电泳,转膜后以5%脱脂牛奶封闭2 h,T-TBS漂洗。E-Cadherin、N-Cadherin、Vimentin(英国Abcam 公司;ab231303,ab76011,ab16700)一抗以1∶1 000稀释,β-actin一抗以1∶2 000稀释,冰箱4 ℃孵育过夜。二抗以1∶5 000稀释,室温摇床孵育1 h,T-TBS 漂洗,ECL 显色。以β-actin(英国Abcam;ab6276)为内参,用Image J 分析目的条带与内参条带灰度值的比值来计算目的蛋白相对表达量。
1.5 划痕实验取对数生长期细胞,以细胞数4×105个/孔接种于六孔板中。待细胞铺满板底后,使用10 μL体积的移液器吸头在板底中心进行笔直刮擦。用PBS(磷酸盐缓冲盐溶液,HyClone,美国)洗涤后,添加无血清培养基,24 h后进行显微镜观察和成像。
1.6 Transwell 实验提前24 h 配置matrigel 基质胶,均匀铺于transwell 小室底部。取对数生长期细胞,胰酶消化后,将每组细胞悬浮在无血清培养基中,配置成2×105个·mL-1细胞浓度,取200 μL位于上室,将完全培养基(700 μL)置于下室,培养24 h。然后用甲醇处理20 min,结晶紫染色。最后,使用倒置显微镜任意捕获5个视野,在20倍镜下进行细胞计数。
1.7 统计学分析采用SPSS 21.0 及Graph Prism 7.0 软件进行分析及统计绘图,配对t检验用于两组间比较,实验数据以均值±标准差进行表示。所有实验均重复3次,P<0.05为差异有统计学意义。
2 结 果
2.1 qRT-PCR 检测三种乳腺癌细胞株LncRNA MEG3 表达水平通过qRT-PCR 检测得出,三种乳腺癌细胞株中,MCF-7细胞中LncRNA MEG3(0.49±0.037)表达水平较MDA-MB-231(0.92±0.049)与SKBR-3(0.67±0.023)表达更低,差异有统计学意义(P<0.05)。因此选取细胞株MCF-7 进行质粒转染,并进行接下来的实验,见图1。
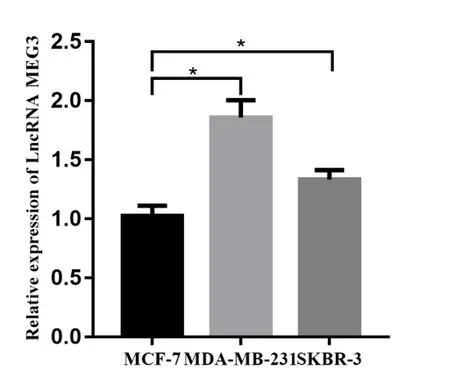
图1 qRT−PCR检测三种乳腺癌细胞中LncRNA MEG3基因相对表达水平(P<0.05)
2.2 qRT-PCR 验 证pcDNA3.1-MEG3 重组质 粒转染效率通过qRT-PCR 检测得出,经过pcDNA3.1-MEG3 转染的MCF-7 乳腺癌细胞株(12.7±0.750)与转染空载体(1.19±0.096)和未转染(1.3±0.202)的细胞株相比,LncRNA MEG3 表达水平升高(P<0.05),差异有统计学意义(P<0.05),见图2。
2.3 qRT-PCR 检测转染后乳腺癌细胞中miRNA-21 的表达量变化通过qRT-PCR 检测得出,经过pcDNA3.1-MEG3转染的MCF-7乳腺癌细胞(2.947±0.436 3)与转染空载体(5.663±0.331 9)和未转染(6.617±0.291 6)的细胞株相比,miRNA-21的表达量降低,差异有统计学意义(P<0.05),见图3。
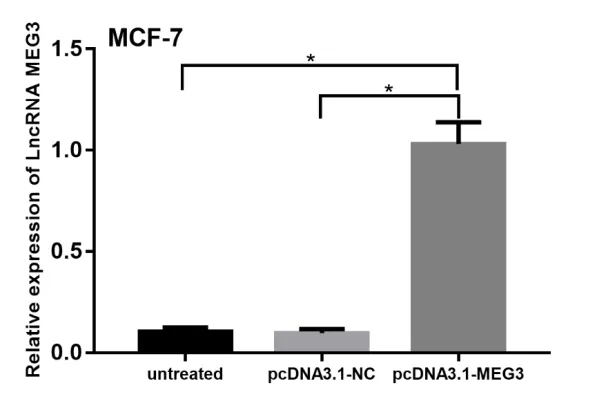
图2 转染后检测LncRNA MEG3在MCF−7细胞中表达水平(P<0.05)
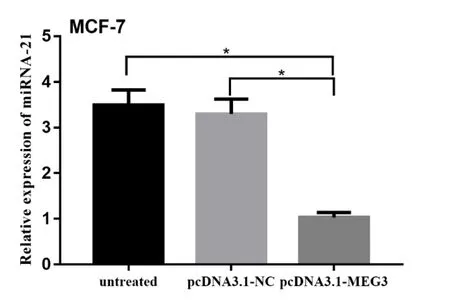
图3 LncRNA MEG3 过表达抑制MCF−7 细胞miRNA−21 的表达量(P<0.05)
2.4 LncRNA MEG3 过表达抑制乳腺癌细胞MCF-7 迁移划痕实验结果显示,过表达组细胞迁移率为(17.6±1.323),而转染空载体组与未转染组迁移率分别为(35.7±1.572)与(31.27±2.085),过表达组迁移率低于空载体与未转染组,差异有统计学意义(P<0.05),见图4。
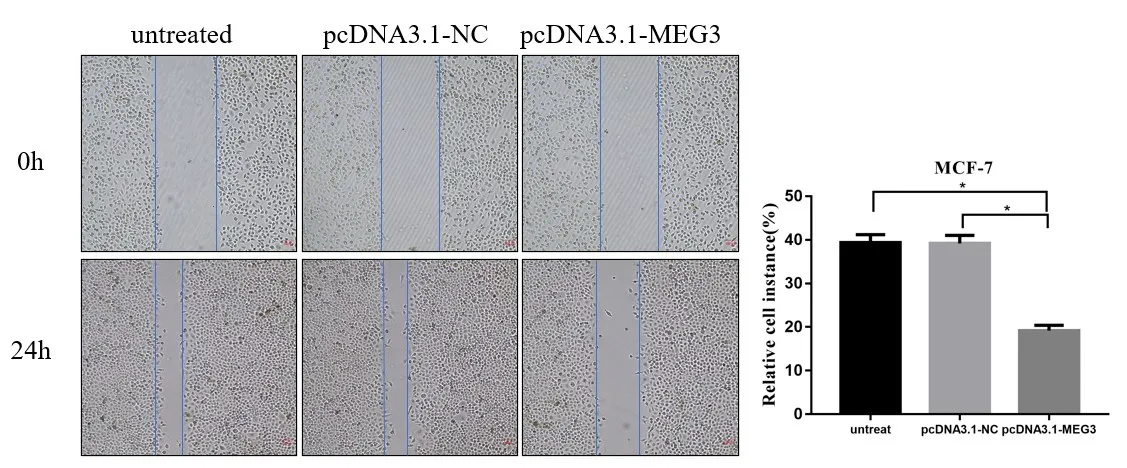
图4 LncRNA MEG3过表达抑制MCF−7细胞迁移能力
2.5 LncRNA MEG3 过表达抑制乳腺癌细胞MCF-7 侵袭Transwell 实验结果得出,过表达组细胞穿膜率为(40.73±4.087),而转染空载体组与未转染组迁移率分别为(72.12±1.898)与(71.67±1.846),过表达组侵袭能力低于空载体与未转染组,差异有统计学意义(P<0.05),见图5。
2.6 LncRNA MEG3 影响MCF-7 乳腺癌细胞EMT相关蛋白表达Western blot实验结果显示,相较于空载体组与未转染组,过表达组中E-Cadherin相对表达量升高,而N-Cadherin 与Vimentin 相对表达量则降低,差异均有统计学意义(P 均<0.05),见图6。
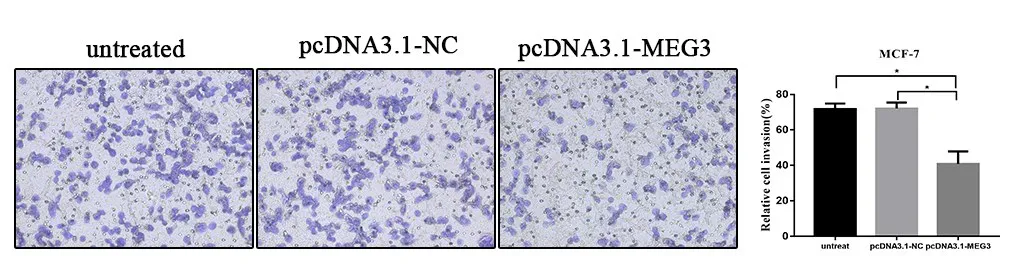
图5 LncRNA MEG3过表达抑制MCF−7细胞侵袭能力
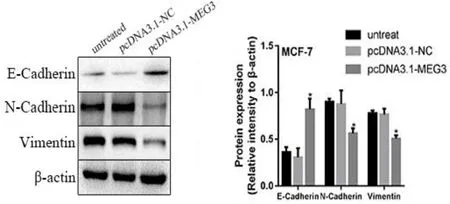
图6 LncRNA MEG3过表达的MCF−7细胞EMT相关蛋白的变化情况
3 讨 论
乳腺癌是全世界妇女最常见的恶性肿瘤之一,近年来,我国乳腺癌发生率及死亡率逐年上升,并呈年轻化发展趋势[8]。乳腺癌的根本治疗手段为手术治疗,但手术治疗对于晚期转移患者的治疗效果并不理想。所以乳腺癌患者的早发现,早诊断,早治疗对于病情的控制非常重要。因此,寻找乳腺癌相关标志物及获取相关生物学指标对于早期的癌症筛查工作具有重要意义。
已有研究证实,LncRNA在基因表达调控方面发挥着极其重要的作用,WANG 的研究表明lncRNA SNHG7 促进胃癌细胞增殖,抑制胃癌细胞凋亡[9]。ZHANG 等发现lncRNA Linp1 在人类三阴性乳腺癌中被过度表达,并加强细胞DNA 的修复[10]。这些研究表明,LncRNA 可能成为恶性肿瘤早期诊断筛查的特异性标志物。且最新研究发现LncRNA MEG3可通过多种机制参与细胞蛋白表达的调控,DAN 等学者研究发现LncRNA-MEG3 在胃癌中通过调节miRNA-21 抑制增殖和转移[11]。LING 等学者指出LncRNA-MEG3 抑制细胞生长并以miR-93 为靶点,抑制PI3K/AKT信号传导通路[12]。但LncRNA-MEG3在乳腺癌中的作用尚不清楚,本研究通过过表达MCF-7细胞中LncRNA-MEG3,检测MCF-7细胞侵袭及迁移的变化情况,并联系EMT 相关蛋白变化情况,探讨相关机制研究。
本研究发现,通过pcDNA3.1-MEG3 重组质粒成功过表达MCF-7 细胞中LncRNA-MEG3,并通过qRT-PCR 发现miRNA-21 的表达明显受到抑制。我们采用划痕实验及Transwell 实验,分别检测了MCF-7 细胞迁移及侵袭能力的改变情况,发现LncRNA-MEG3 能够有效的抑制MCF-7 细胞的迁移及侵袭能力。并通过Western blot实验,检测了EMT相关蛋白表达情况,过表达后的MCF-7细胞E-Cadherin蛋白表达量上调而N-Cadherin及Vimentin蛋白表达量下调。
综上所述,LncRNA-MEG3 的表达能够调控miRNA-21的表达变化,进而改变乳腺癌细胞MCF-7侵袭及迁移能力,且LncRNA-MEG3 表达水平越高,miRNA-21 表达水平越低,细胞侵袭及迁移能力就越弱,且这种调节机制可能与EMT 相关蛋白调控有关。但本文只是对细胞学方面做了一部分探讨,没有真正调查临床患者的实际情况,存在一定的局限性,下一步准备收集病患资料,研究LncRNA-MEG3 是否会影响患者生存率。总之,本研究是首次报道LncRNA-MEG3 与乳腺癌细胞之间的相关性,我们希望能够作为一个新的思路运用到乳腺癌的相关生物学指标及靶向治疗当中去。
猜你喜欢
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
广西林业科学(2016年2期)2016-03-20
中国医药生物技术(2015年4期)2015-12-26
广州大学学报(自然科学版)(2015年4期)2015-12-23
医学研究杂志(2015年11期)2015-06-10
中国当代医药(2015年16期)2015-03-01
中国医药导报(2015年27期)2015-02-28
中国酿造(2014年9期)2014-03-11
