
谷子JAZ 家族全基因组鉴定及表达分析
2021-04-26 11:17张琦常国蓉张育铭刘旭张宝俊
山西农业大学学报(自然科学版) 2021年2期
张琦,常国蓉,张育铭,刘旭,张宝俊
(山西农业大学 植物保护学院,山西 太谷030801)
茉莉酸(Jasmonic acid,JA)是植物体内合成的一类重要植物生长激素,在调控植物的生长发育及应答病菌侵染、昆虫为害等环境胁迫方面发挥着重要作用[1]。JAZ(Jasmonate ZIM-domain)蛋白是植物中特有的锌指蛋白,参与植物抵御生物和非生物逆境胁迫信号的响应和调控,在JA 信号转导通路研究中扮演着“抑制子”角色[2]。JAZ 蛋白含有2 个保守结构域,分别位于为N 端的ZIM(又称TIFY)结构域及位于C 末端的Jas(又称CCT_2)结构域。ZIM 结构域参与二聚体的构成及与NINJA等抑制子的结合[3];Jas 结构域主要负责与受体复合体中COI1及MYC2等转录激活物的结合[4]。植物细胞中JA 的浓度较低时,JAZ 蛋白与MYC2结合而受到抑制[5];高浓度的JA 可促进JAZ 蛋白与SCFCOI1结合,使JAZ 蛋白泛素化,被26S 蛋白酶降解[6],解除JAZ 蛋白对MYC2转录活性的抑制,激活JA 早期应答基因的表达[7,8]。此外,JAZ 蛋白还参与赤霉素(GA)、生长素(IAA)、乙烯(ET)和水杨酸(SA)等其他植物激素生物合成或信号传导途径[9]。
JAZ 基因家族成员众多,生物学功能丰富,在玉米、水稻、小麦、陆地棉和粗山羊草中分别发现了23、15、14、30 及9 个JAZ基因[10~14]。拟南芥中缺失Jas 功能结构域AtJAZ1基因能够增强拟南芥对甜菜夜蛾(Spodoptera exigua)的抗性[15];水稻中OsJAZ9作为茉莉酸盐信号转导的转录调节因子,可以调节水稻的盐胁迫或缺钾耐性[16];大豆中GsJAZ2是一个新基因,其过度表达可以增强拟南芥对盐和碱胁迫的耐受性[17]。
JAZ 基因在调节生物和非生物胁迫的适应或防御以维持植物生长方面发挥了重要作用。谷子(Sitaria italica)是起源于我国的粮食作物,在食物多样性和种植业结构调整中具有不可或缺的作用[18]。近年来,随着种植面积的不断扩大,以谷子白发病、谷瘟病、粘虫等为主的谷子病虫害在生产上频发。同时,谷子很容易受到各种非生物胁迫,如盐碱、干旱等都会影响谷子的生长发育。鉴定谷子JAZ 家族基因,探究不同成员在根、茎、穗和叶中的差异表达及其调控网络,有利于深入解析JAZ 家族基因在谷子生长发育及抗病其它代谢通路中发挥的作用,为谷子育种提供一定的科学依据。
1 材料和方法
1.1 谷子JAZ 基因家族鉴定
自Phytozom 数据库(https://phytozome.jgi.doe.gov/pz/portal.html)中获取谷子(Setaria italicav2.2)、水稻(Oryza sativav7_JGI)和拟南芥(Arabidopsis thalianaTAIR10)的全基因组数据文件。分别从TAIR 数据库(https://www.arabidopsis.org/)和水稻基因数据库(http://www.ricedata.cn/gene/)获取到11 条拟南芥和15 条水稻JAZ(Jasmonate ZIM-domain)蛋白序列作为搜索谷子同源基因的探针序列。利用Pfam 数据库(http://pfam.xfam.org/)构建的TIFY(也称为ZIM)(PF06200) 和 Jas( 也称为 CCT_2)(PF09425)结构域的隐马尔科夫模型,在TBtools[19]工具中利用Simple HMM Search 程序,在谷子全基因组蛋白序列中搜索同源序列。选取比对值E-value<0.01。
1.2 谷子JAZ 基因蛋白特性分析
使用Pfam 中 的Batch Search(http://pfam.xfam.org/Siarch#tabview=tab1)对检索到的蛋白序列结构域进行验证。利用ExPASy-ProtParam tool 在线工具(https://web.expasy.org/protparam/)预测谷子JAZ 家族的蛋白序列的氨基酸数目、分子量、等电点和亲水性均值。
1.3 谷子JAZ 基因系统进化分析及保守结构域分析
使用MEGA7[20]对筛选到的拟南芥、水稻和谷子的JAZ 家族蛋白序列进行多序列比对,采用最大似然法(Maximum Likelihood Estimate,MLE)构建系统发育树,其中Boot Strap 参数设置为1000,其它参数选择默认。借助在线工具ITOL(https://itol.embl.de/)进一步优化进化树,利用MEME 在线软 件(http://meme-suite.org/tools/meme)分析谷子JAZ 基因家族蛋白序列保守motif,并运用TBtools 工具作图。
1.4 谷子JAZ 基因家族的启动子分析
将JAZ 家族基因起始密码子上游2 000 bp 的基因组序列作为启动子序列提交到Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)数据库,预测JAZ 家族基因的启动子区顺式作用元件,并运用Tbtools 工具作图。
1.5 谷子JAZ 家族基因的组织特异性表达分析
从Phytozome 数据库获得谷子根、茎、穗、叶不同组织的基因表达量数据。于2020 年5 月在山西农业大学谷子杂粮育种基地,采取随机区组设计方法划分试验小区,小区面积为3 m×2 m。在谷子抽穗开花期取材,样品经液氮速冻后于−80 ℃保存。采用Trizol 试剂(Sangon Biotech)分别提取根、茎、穗和叶的RNA。采用反转录试剂盒(Takara)合成cDNA。使用primer5 设计JAZs 基因特异性引物,以actin为内参(表1),采用2×SG 快速qPCR 混合试剂盒(Takara)进行荧光定量分析。qRT-PCR 反应体系(15 μL):2×SG 快速qPCR 主混合物7.5 μL、上下游引物各0.6 μL、40 ng cDNA模板、ddH2O 3.3 μL。反应条件:95 ℃变性30 s,95 ℃变性5 s,60 ℃退火/延伸30 s,40 个循环,用2-ΔΔCT法分析各个基因在不同组织的相关表达水平,3 次生物学重复。
2 结果与分析
2.1 谷子JAZ蛋白家族筛选、理化性质和基因定位
利用Jas 和TIFY 结构域的HMM 模型在谷子基因组中分别鉴定得到25 个和39 个候选成员,通过Pfam 数据库分析后,最终筛选得到谷子JAZ 家族 基 因21 个,命 名 为SiJAZ1~SiJAZ21,编 码156~424 个氨基酸,平均氨基酸数目为234 个,蛋白分子量介于16.22~44.93 kDa(表2)。该蛋白的GRAVY 值均为负值,预测该家族蛋白为亲水蛋白。在染色体定位中,JAZ 家族基因在Chr9 染色体上的数目最多为11 个,在Chr3 和Chr6 染色体上的数目各仅有1 个。

表1 Si JAZs 基因序列扩增qRT-PCR 引物Table 1 qRT-PCR primers of Si JAZs gene sequence amplification
2.2 谷子JAZ 家族蛋白序列系统发育分析及保守结构域分析
构建的谷子JAZ 蛋白序列系统发育树显示,谷子JAZ 家族基因被分为4 大类(图1)。其中在绿色分支中有6 个谷子JAZ 家族成员与5 个拟南芥JAZ 家族成员同源性较高;红色分支中有1 个谷子JAZ 家族成员与2 个拟南芥JAZ 家族成员同源性较高;在蓝色分支中,有2 个谷子JAZ 家族成员与2个水稻JAZ 家族成员和4 个拟南芥JAZ 家族成员同源性较高;在黄色分支中,有10 个谷子JAZ 家族成员与12 个水稻JAZ 家族成员同源性较高,有且仅有水稻中的JAZ 家族均分布在此分支中。黄色分支中的谷子JAZ 家族基因与水稻存在较近的亲缘性。
21 个JAZ 蛋白序列保守基序分析共鉴定到4个基序,分别命名为motif 1~motif 4(图1)。结果表明,所有成员都含有TIFY motif 和Jas motif,其中motif 1 与motif 2 最为 保守,在21 条JAZ 蛋白 序列中均存在,且分布顺序均为motif 1 在前,motif 2在后,motif 3、motif 4 在大部分的JAZ 家族基因中都存在。保守基序的序列分析表明,其中谷子motif 1 中较为保守的序列为APAQLTIFYGGRV-LVFDDVPA,motif 2 中较为保守的序列为 RKASLQRFLEKRKDR(图2)。

表2 谷子JAZ 家族基因的序列特征Table 2 Sequence characteristics of the JAZ family in S.italica
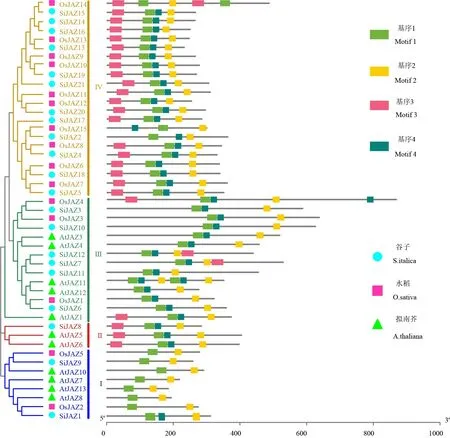
图1 谷子、水稻和拟南芥的JAZ 家族系统进化树及蛋白motif 分析Fig.1 Phylogenetic tree and protein motif analysis of JAZ family of S.italica,Arabidopsis thaliana and Oryza sativa
2.3 谷子JAZ 家族基因启动子顺式作用元件
顺式作用元件分析结果显示,SiJAZ2,Si-JAZ3,SiJAZ10,SiJAZ16和SiJAZ20基因含有防御和胁迫反应响应元件(图3),表明它们可能参与调控谷子对病菌侵染的响应,其中,SiJAZ3基因的顺式调控元件最多,高达48 个;SiJAZ11基因最少,仅含有5 个。21 个JAZ 家族基因中,均具有脱落酸响应元件(abscisic acid responsiveness,ABRE),14 个基因具有赤霉素响应元件(P-box,TATC-box 和 GARE-motif,gibberellin-responsive element),19 个基因具有MeJA 响应元件(CGTCA-motif 和TGACG-motif)和3个基因具有水杨酸响应元件(TCA-element),表明谷子JAZ基因家族与激素信号紧密相关。
2.4 谷子JAZ 家族基因的组织表达分析
谷子JAZ 家族部分基因组织特异性表达结果见 表4。qRT-PCR 分析结果表明,SiJAZ4,Si-JAZ12,SiJAZ14,SiJAZ17,SiJAZ20分别在谷子的根、茎、叶穗部等组织中均有表达,其中SiJAZ4在茎中表达量较高,在根,穗,叶中表达量较低;Si-JAZ12和SiJAZ20在叶中表达量最高,尤其Si-JAZ12表达量可达到0.7,在茎中次之,在根和穗中较低,推测SiJAZ12和SiJAZ20可能在叶中起重要作用;SiJAZ14和SiJAZ17仅在根中表达量高,在其余组织中表达量较低。测定结果表明,谷子JAZ 基因家族的表达存在组织特异性表达。

图2 谷子JAZ 家族氨基酸序列保守基序Fig.2 Conserved motifs in the amino acid Sequences of JAZ family in S.italica

图3 谷子JAZ 家族基因启动子顺式作用元件功能Fig.3 Function of promoter cis-acting elements in JAZ family genes in S.italica

图4 谷子JAZ 家族基因(部分)的组织特异性表达Fig.4 Tissue-specific expression of S.italica JAZ family genes(parts)
3 讨论与结论
茉莉酸是一种重要的生物胁迫、非生物胁迫防御胁迫响应激素。在JA 信号转导途径中,抑制因子JAZs 蛋白广泛参与到植物生长发育及抵抗胁迫的过程中,DELLA 蛋白可以促进JA 应答基因的表达,但DELLA 蛋白是一类能够抑制GA 信号传递的重要调控因子,对植物起负反馈作用[21]。研究表明DELLA 阻遏物可能通过与JAZ1的物理相互作用促进JA 信号传导[22],JAZs 与DELLAs 相互协同,调控植物的生长与防御。此外,JAZ 蛋白在茉莉酸和乙烯、生长素和脱落酸的互作中也均有 报 道[23,24]。在 水 稻 中,OsJAZ1通 过JA 与ABA途径调节抗旱性[25];面包小麦中TaJAZ1负调控脱落酸(ABA)介导的种子发芽抑制和ABA 响应基因表达[26];野生葡萄中JAZ4的过表达通过SA 或JA 信号通路提高了对葡萄白粉病的抵抗力[27]。因此,JAZ 在激素信号的传导中具有重要地位。
本研究利用生物信息学分析,共鉴定到21 个谷子JAZ 基因家族成员,均具有TIFY(又称ZIM)和Jas(又称CCT_2)结构域。目前在拟南芥中鉴定到12 个JAZ 蛋白,2 个结构域之间有131~352个氨基酸,而且TIFY 和Jas 的相对位置也有变化[28]。系统发育分析表明,谷子中该家族分为4 个亚家族,且谷子与水稻同源性更高一些,可能因为在进化过程中发生了基因丢失或该家族基因进化速率较快导致基因间差异较大,这种现象在禾本科植物中普遍存在。基因结构和Motif 分析表明,多数基因具有相似的结构及组成,Motif 的组成及排布也有一定的规律,部分Motif 有位置和数量的变化,可能与谷子在病原菌等生物胁迫或非生物胁迫条件下产生的遗传变异有关。组织特异性表达分析显示,SiJAZ12和SiJAZ20在叶中表达量最高;它们可能与叶部发育密切相关。有研究表明拟 南 芥 中AtJAZ1、AtJAZ3、AtJAZ4、AtJAZ9和AtJAZ10基因参与叶片的生长调控[29]。其中Si-JAZ12在叶中的表达量接近1,这表明它可能通过参与谷子叶片的生长发育参与茉莉酸及其它激素调控。SiJAZ4基因在茎中的表达量高于其他组织,这与水曲柳(Fraxinus mandshurica)FmJAZ1基因在茎中表达量最高的结果一致[30]。SiJAZ14和SiJAZ17在根中表达量较高,其余组织中很低,苹果中MdJAZ1基因在根中表达量最高[31]。这表明SiJAZs基因的表达具有组织特异性,且不同的JAZ 家族成员表达的组织部位不同。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
当代陕西(2021年21期)2022-01-19
延河(2018年6期)2018-06-11
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
