
急性心理应激对协同运动知觉的影响及其机制:基于ERP的证据
2021-04-24 02:58王积福贺梦阳漆昌柱
体育科学 2021年2期
王积福,贺梦阳,漆昌柱*
(1.长江大学 教育与体育学院,湖北 荆州 434023;2.武汉体育学院 心理学系,湖北 武汉 430079)
急性心理应激是指不可控制且不可预期的外在环境要求短时间内超出了身体的调节能力时,有机体做出的一种非特异性反应(Koolhaas et al.,2011)。由于急性应激是有机体短时内接受超出身体承受范围的外在刺激,具有持续时间短、无躯体明显痛苦以及强度大等特点。前期研究发现,急性应激会增加人们的警觉性和感官信息输入(Shackman et al.,2011)。研究还发现,在高强度心理唤醒状态下,人们对生物运动知觉任务的反应时间较短(Niederhut,2009)。在神经水平上,急性应激通过增加多巴胺、去甲肾上腺素和糖皮质激素介导的信号传导来影响依赖前额叶皮层(prefront cortex,PFC)的认知功能(Arnsten,2009;Sänger et al.,2014)。研究表明,应激会削弱需要PFC参与的任务绩效,依赖于基底神经节回路的行为习惯方式以保留或增强(Arnsten,2009;Sandi et al.,2007)。在一些简单的任务中,或者当认知负荷不过度时,急性应激倾向于促进个体的认知功能(Qi et al.,2017)。协同运动知觉(coherent motion perception)作为认知功能领域中的一个重要组成部分,同时也是评价个体整体-局部运动知觉能力的重要指标之一(Robertson et al.,2014)。
协同性运动是指运动的点或客体之间相互独立,没有相同的运动规律,但其中一定数量客体的运动存在知觉特性中的“共同命运”原则,即有着相同的运动方向,具有知觉整体性的特点(Newsome et al.,1988)。当这些“共同命运”的点达到一定的数量后,被试则会察觉这些点会朝着同一个方向运动。“共同命运”的点占全部散点的比例称为协同性或协同性水平,如100个点中有50个点的运动方向是一致的,则协同性水平为50%。一般可通过被试按键判断点刺激的运动方向(向左、向右、向上或向下)作为判断被试对协同性水平感受力的途径。研究发现,感受力大小与协同性阈值大小成反比,即当被试感受力越高,则只需要较少的点(共同命运)就能判断出运动方向(胡奂,2013)。关于协同运动知觉的研究范式主要采用平面上的光点刺激任务来实现,并控制协同性水平(0、50%、100%等),如 RDK 任务(random-dot kinematogram)(Braddick,1974)、GDM 任 务(global dot motion task)(Newsome et al.,1988)等,点的运动包括水平运动与扩展运动。本研究采用RDK任务中的水平运动来实现协同运动知觉的判断任务。
协同运动知觉受到多种因素的影响。有研究发现,其影响因素主要包括自闭症、情绪状态、年龄、认知风格、智力、刺激特征(刺激类型、呈现时间)等(Hallen et al.,2019)。Koldewyn等(2011)研究发现,自闭症患者的协同运动知觉阈限显著高于健康群体,即自闭症患者只能判别出协同性水平较高的光点运动方向。自闭症患者的严重程度与其协同运动知觉阈限的大小成正比例相关,即自闭症患者程度越低,协同运动知觉阈限越低(Grinter et al.,2009)。Kunchulia等(2019)研究发现,与年轻人相比,老年人的协同运动知觉感受力有所下降,即协同运动知觉阈限增加。在这些影响因素当中,鲜见考察急性心理应激对协同运动知觉影响效果的研究。但也有研究发现急性心理应激对其他认知活动的影响,Qi等(2020)发现,急性心理应激会促进选择性注意过程中的注意控制。另一研究发现,急性心理应激可以提升深度运动知觉任务的行为绩效,以及增强其注意资源的投入(Wang et al.,2020)。从研究现状来看,急性心理应激与协同运动知觉的关系尚不明确,但有关情绪与认知之间的理论假说认为,急性心理应激可能会对个体的协同运动知觉能力产生一定程度的影响。
卡尼曼(Kahneman)提出的认知资源占用学说认为,个体的认知资源有限,人们在特定时间内只能对有限资源进行分配,认知资源的本质是将个体可用于完成认知任务时的注意看作是一种有限的资源(王甦等,2006;Kahneman et al.,1979);Pessoa(2009)提出的双竞争理论模型认为,认知加工与情绪刺激加工同时进行时,它们会对有限的认知资源进行竞争。急性心理应激属于情绪刺激,而协同运动知觉任务属于认知加工过程,二者在先后加工过程中一定会出现相互影响。Qi等(2017)研究发现,人们在应激条件下go/nogo任务中表现出P2波幅减小而N2波幅增加,说明人们在急性心理应激状态下可以通过减少早期选择性注意过程和增强认知控制过程来改变反应抑制过程。本研究结合行为学和EEG指标,进一步探究急性心理应激对协同运动知觉的影响及其内在作用机制。
关于协同运动知觉的ERP成分,Taroyan等(2011)采用3种水平的协同运动任务(10%、25%和40%)测试了9名患有阅读障碍的青少年的协同运动知觉能力,结果发现,早期ERP成分(P1、N1、P2)或晚期ERP成分峰波幅与峰潜伏期之间差异不显著。为满足ERP实验的要求,本研究选取改良后的乘法估算任务(MIST任务)作为实验室急性心理应激诱发的手段(Dedovic et al.,2005),改良后的MIST任务中主要包括两种急性心理应激情境:不可控性(应激条件下呈现时间很短)与社会性评价威胁(负反馈)。选取经典的RDK任务范式(Braddick,1974)作为评价协同运动知觉能力的任务。旨在通过采用具有高时间分辨率的ERPs实验技术探究急性心理应激对协同运动知觉影响的深层次电生理学特点。依据相关理论基础,研究假设:若应激条件与控制条件下被试在协同运动知觉任务中行为学表现存在差异,则协同运动任务中P1、P2、N2成分及晚期正成分(late positive potential,LPP)平均波幅会存在显著性差异,且随着协同性水平的逐渐升高,被试的反应时逐渐缩短、正确率逐渐升高。
1 研究方法
1.1 被试
参照Qi等(2017)的研究,并通过G*power 3.1.9计算样本量大小(Faul et al.,2007),探测重复测量方差分析中的主效应及其交互作用(组内设计),使其统计检验力达到0.8及中等效应量(d=0.25)所需要的总样本数为19。随机选取某院校25名在校大学生,并采用贝克抑郁量表(Beck,1967)、情绪状态评价量表(漆昌柱 等,2007)、UCLA孤独量表(Russell,1996)以及镶嵌图形测验(邓铸等,2008)对被试的情绪状态、孤独状态及空间思维能力进行筛查,发现25名被试均不处于抑郁状态和自闭状态,同时所有被试情绪状态也未出现过度兴奋或过度负性状态;镶嵌图形测验中所有被试的认知风格均为场独立型,都处于同一基线水平。因此,排除了相关被试额外变量对实验结果的影响。所有被试矫正视力正常,且均为右利手。由于有2名被试脑电伪迹过多而被剔除(伪迹试次数量大于总试次的50%而造成叠加次数过少),最后实际纳入行为学与ERP数据分析的被试有23名(12男,11女,平均年龄19.78岁)。实验过程得到学校人体实验伦理委员会的批准。
1.2 实验设计
该研究为两因素被试内实验设计,自变量为应激水平2(应激条件与控制条件)×协同性水平3(50%、75%、100%),其中为提升被试的自我卷入程度增加协同性水平为0%的刺激材料,两个自变量均为组内变量。因变量为被试反应时(刺激呈现到被试按键反应)、反应正确率、ERP相关成分(P1、P2和N2)的峰波幅值与峰潜伏期,以及LPP的平均波幅值。
1.3 材料
实验材料主要包括两个部分:
其一,急性心理应激诱发参照改良后的MIST任务范式,为160个乘法心算题目(如4.78×2.16等),被试需要判断所乘结果是大于10还是小于10,若小于10则需按键盘上的F键,若大于10则需要按键盘上的J键。
其二,协同运动知觉刺激材料采用Newsome等(1988)研究协同运动知觉的RDK范式,协同运动方向为水平流动(向左或向右),其中随机点与协同运动点的比例分别为0%、50%、75%、100%,随机点与协同运动点的数量总计100个[点的颜色为白色;视角:(0.3×0.3)o;密度:1.85点/度2;移动速度:5.0o/s],黑色背景[视角:(50×48)o;亮度:0.1 cd/m2],协同点的运动时间为2 000 ms,实验视频使用Matlab软件制作,一共7个视频,视频格式均为WMV格式,分辨率为800×600 dpi。为增加被试实验过程中的自我卷入程度,加入协同性水平为0的刺激材料,协同性水平为0的刺激是指视频中100个点的运动方向均是随机的,被试不需要做出按键反应,同时也可以作为被试是否认真参与实验的一个指标。其他刺激材料中,当协同运动方向为向右时,被试需要按下键盘上的J键,当协同运动方向为向左时,被试需要按下键盘上的F键。
1.4 过程
1)被试进入实验室后,填写被试知情同意书、贝克抑郁量表(基本信息部分包括利手测验等)、情绪状态评价量表、状态特质焦虑量表、UCLA孤独量表以及镶嵌图形测验,然后并对其结果进行筛查,排除抑郁患者、过度兴奋、负面情绪过多、自闭症以及空间认知能力较差的被试。2)主试为被试佩戴Easycap64导脑电帽,注入导电膏,使所有电极点阻抗均降到5 kΩ以下。3)主试结合具体实验内容以及实验指导语给被试讲解实验的具体任务要求,被试明白实验内容和任务后,身体坐正,并保持眼睛距离电脑屏幕中央约70 cm,并告知实验过程中尽量不要出现摆头、摆腿等大的动作。4)心率调整阶段。屏幕上呈现一张放松图片,被试仔细观看并表象图片里边的内容,想象自己身临其境,调整呼吸,放松时间为60 s。5)练习与正式实验阶段(图1)。两个阶段主要区别在于练习阶段协同运动知觉任务中有反馈,其反馈界面主要呈现被试方向判断的正确与否或者是否反应。心算任务后的反馈内容为被试反应的正确与否以及其反应时与其他大多数人平均反应时(700~800 ms随机)的比较(大于、等于或小于),控制条件下反馈内容为星号。被试练习结束后,将进入正式实验阶段。正式实验阶段包括两个block:其一为应激条件(160个trail);另一个为控制条件(160个trail),共 320个 trail,协同性水平分别为 0%、50%、75%、100%,均包含40个trail(左右各一半),其中每个trail随机呈现。应激条件与控制条件之前均有一个练习block。其中,为排除任务难度的影响,应激条件与控制条件下心算任务的题目是一致的。由于研究为组内设计,为避免顺序效应造成的实验误差,研究采用ABBA设计来进行被试间平衡处理,即应激-控制和控制-应激交替进行,两种条件之间休息5~10 min,以避免两种条件之间的交互影响。此外,在协同刺激视频呈现中会有3种情况:一是所有运动的点均向左或向右(100%条件,向左按键盘上的F键,向右按键盘上的J键);二是所有运动的点均为随机运动(0%条件,不需要进行按键反应);三是部分运动的点向左或向右,其他运动的点均为随机运动(50%、75%条件,向左按键盘上的F键,向右按键盘上的J键)。
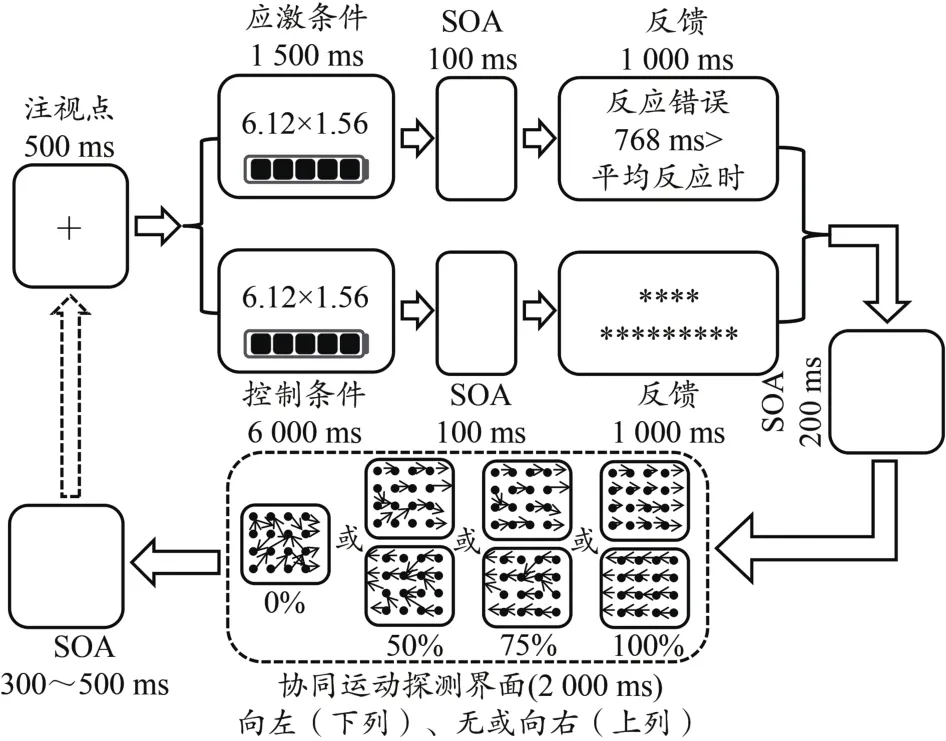
图1 急性心理应激对协同运动知觉影响的正式实验任务流程图Figure 1.Experimental Flow Chart of the Effects of Acute Psychological Stress on Coherent Motion Perception
1.5 行为学数据
行为学数据主要由E-prime 2.0软件采集,并采用Excel 2007软件计算出每名被试应激条件与控制条件下乘法心算任务的反应时与正确率,以及不同协同性水平下的反应时及正确率。采用SPSS 17.0对23名被试在不同条件下的正确率及反应时进行描述性统计分析。在正确率方面,做Chi-square检验。在反应时方面,以应激水平与协同性水平做两因素重复测量方差分析,若主效应显著,则进行事后检验;若交互作用显著,则进行简单效应分析。
1.6 脑电记录与分析
脑电数据主要通过Recorder软件采集,采用BrainVision Analyzer 2软件进行离线叠加分析与处理。实验所采用BP设备的64导电极帽按照国际10-10系统安置电极,AFz点为接地电极。在线记录的参考电极为FCz点,后期离线处理时转换为平均参考。垂直眼电贴于右眼正下方1 cm处,水平眼电贴于左眼眼角外1 cm处。信号采样频率为1 000 Hz,滤波范围0.01~100 Hz。离线脑电数据处理时滤波范围0.01~35 Hz,采用独立成分分析法(independent component analysis)识别并去除眼动伪迹,同时去除波幅超过±80 μV的伪迹(artifact rejection)。为探究不同应激水平下被试协同运动知觉的ERP特征,依据协同运动知觉刺激呈现时长与被试实际反应结果将EEG叠加时长定为-200~1 000 ms(共1 200 ms),基线为探测刺激呈现前的200 ms。关于ERP的叠加次数,Picton等(2000)认为,由于ERP实验过程中会受到眼跳、肌肉伪迹与反应错误等因素的影响,建议各实验条件叠加次数大于30次。通过统计分析,各实验条件平均叠加次数约为33次,大于30次。
ERP分析分为应激诱发部分与协同运动知觉部分。一方面,在应激诱发部分,根据前期研究结果(Bertsch et al.,2011;Qi et al.,2016),并结合ERPs总平均、差异脑地形图,应激诱发阶段选取Pz和POz作为分析N1成分的电极点,N1的分析时间窗为 80~200 ms(Reinvang et al.,2000);选取PO3、PO4和POz作为分析P2成分的电极点,P2成分的分析时间窗为 150~250 ms(O’toole et al.,2012)。另一方面,在协同运动知觉部分,依据前人研究基础(Manning et al.,2019;Niedeggen et al.,1999),协同运动知觉分析的ERP成分主要包括P1、P2、N2以及晚期正成分(LPP)。结合ERPs总平均和差异脑地形图,各成分的时间窗和电极点具体如下:P1成分的时间窗为刺激呈现后60~160 ms,分析电极点为左右后颞枕区P3、P5、PO7、P4、P6、PO8;P2成分的时间窗为 150~300 ms,分析电极点为P3、P5、PO7、P4、P6、PO8;N2成分的时间窗为刺激呈现后100~300 ms,分析电极点为枕区POz、PO3、PO4;晚期正成分LPP的时间窗为400~-600 ms,分析电极点为P3、P5、P4、P6。采用重复测量方差分析方法对应激诱发部分和协同运动知觉部分产生的ERP成分进行差异检验,包括主效应和简单效应分析。重复测量方差分析中若不满足球形检验,则方差分析的P值采用Greenhouse Geisser法校正,所有主效应的多重比较均通过Bonfferni方法校正。
2 结果
2.1 行为学结果
在应激诱发有效性检验方面,配对样本t检验发现心算任务中应激条件的正性心理唤醒得分(13.86±5.29)显著低于实验前(17.05±4.49),t=3.32,P<0.01;被试在应激条件实验后的状态焦虑得分(43.36±11.70)显著大于实验前的得分(37.50±7.03),t=-2.95,P<0.01(图2)。在心算任务反应时方面,应激条件下心算任务反应时显著短于控制条件(t=-8.81,P<0.01),应激条件下的心算正确率显著小于控制条件(χ2=13.94,P<0.01)。说明被试在应激状态下的心算反应时较快且正确率较低,心算任务起到了应激诱发效果。
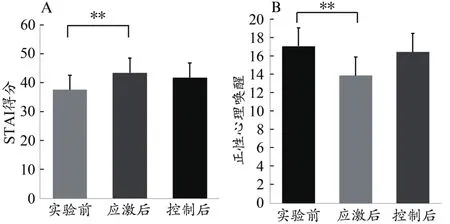
图2 不同应激水平下状态焦虑(A)与正性心理唤醒(B)差异Figure 2.Averaged Subjective Ratings of State Anxiety(A)and Positive Affect(B)in the Stress and Control Conditions
以应激水平(应激与控制)、协同性水平(0.50、0.75、1.0)为被试内变量,对被试反应时进行两因素重复测量方差分析。结果表明,在被试反应时方面:应激水平主效应显著,F(1,22)=10.76,P<0.01,,事后检验发现M应=813.14<M控=906.15;协同性水平主效应显著,F(1.53,33.65)=87.73,P<0.01,,M0.5=1 012.69,M0.75=812.26,M1.0=753.99(图3A),可见随着协同性水平的增加,被试反应时逐渐缩短。事后检验发现,4种协同性水平两两比较均存在显著性差异(P<0.01),即随着协同性水平的增加,被试反应时显著性降低;应激水平与协同性水平交互作用不显著,F(1.44,31.71)=1.97,P>0.05,。在反应正确率方面:以应激水平与协同性水平做交叉表Chi-square检验,结果为χ2=0.26,df=2,P>0.05,说明应激水平与协同性水平两个自变量相互独立,即交互作用不显著。以应激水平做单一变量的 Chi-square检验,结果为χ2=0.04,df=1,P>0.05,说明不同应激水平下被试协同运动方向判断正确率差异不显著。以协同性水平做单一变量的Chi-square检验,结果为χ2=17.84,df=2,P<0.01,说明不同协同性水平下被试判断正确率差异显著(图3B),随着协同性水平的增加,被试反应正确率逐渐增加。
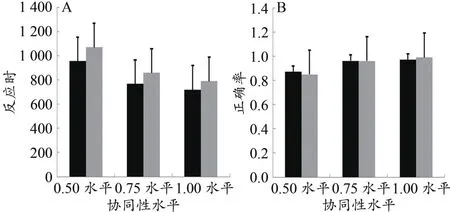
图3 不同应激水平与协同性水平下反应时(A)与正确率(B)结果Figure 3.Results of Reaction Time(A)and Accuracy(B)at Different Stress Levels and Coherent Levels
2.2 电生理学结果
应激诱发部分:在N1峰潜伏期方面,电极位置主效应显著,F(1,22)=8.27,P=0.009<0.01,,事后检验发现POz电极点(137.44±3.31)ms的N1峰潜伏期显著大于Pz电极点(129.87±2.83)ms(P<0.01);应激水平主效应以及电极位置与应激水平交互作用均不显著(P>0.05)。在N1峰波幅方面,电极位置主效应与应激水平主效应均不显著(P>0.05);电极位置与应激水平交互作用显著,F(1,22)=5.42,P=0.03<0.05,,简单效应分析未发现两两之间存在差异。
对于P2峰潜伏期而言,发现应激水平主效应不显著,F(1,22)=0.73,P>0.05,;电极位置主效应不显著,F(2,44)=1.37,P>0.05,;应激水平与电极位置交互作用不存在显著差异,F(2,44)=2.45,P>0.05,。对P2峰波幅而言,应激水平主效应显著,F(1,22)=7.70,P=0.01,,事后检验发现应激水平下P2峰波幅(8.41±0.58)μV显著小于控制条件(9.67±0.59)μV(P<0.05,图4A);电极位置主效应,F(2,44)=1.48,P>0.05,;应激水平与电极位置交互作用不显著,F(2,44)=0.96,P>0.05,。
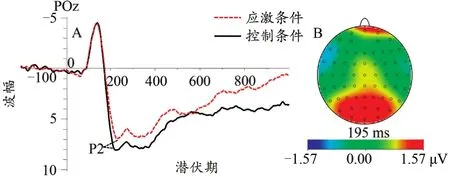
图4 不同应激条件下P2总平均波形图(A)及差异脑地形图(控制-应激,B)Figure 4.The Grand Average Waveform of P2(A)and Topographic Map(B)at Different Stress Levels
协同运动知觉部分。P1成分:对于P1峰潜伏期而言,应激水平主效应不显著,;协同性水平主效应显著,M0.50=109.53 ms,M0.75=123.92 ms,M1.00=116.73 ms,事后检验发现,0.50水平协同运动的P1峰潜伏期显著短于0.75水平协同运动(P<0.01)与1.00水平协同运动(P<0.01);电极位置主效应显著,事后检验发现,P3电极点的P1峰潜伏期显著短于P5电极点(P<0.01),P4电极点的P1峰潜伏期显著短于PO8电极点(P<0.05);电极位置与协同性水平交互作用显著,,进一步做简单效应分析表明,在0.50水平上P5电极点P1峰潜伏期显著大于P4电极点(P<0.01)、P6电极点(P<0.05)和PO8电极点(P<0.05),在PO7与P4电极点0.50水平协同运动P1潜伏期显著小于0.75水平协同运动(P<0.05),在P6电极点0.5水平协同运动P1潜伏期显著小于1.00水平协同运动(P<0.05),在PO8电极点0.50水平协同运动P1潜伏期显著小于0.75水平与1.00水平协同运动(P<0.01),以及0.75水平协同运动P1潜伏期显著大于1.00水平协同运动(P<0.05),其余两两比较之间差异均不显著(P>0.05);其余交互作用均不显著(P>0.05)。
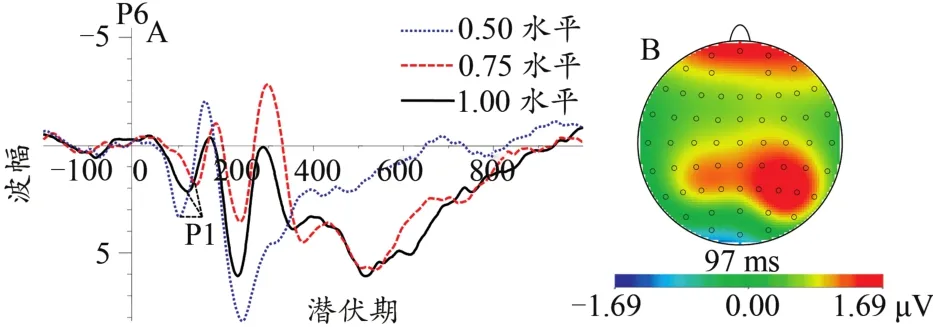
图5 不同协同性水平下P1总平均波形图(A)及差异脑地形图(0.50~1.00水平,B)Figure 5.The Grand Average Waveform of P1(A)and Topographic Map(B)at Different Coherent Levels
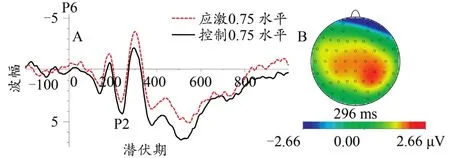
图6 不同应激水平下P2总平均波形图(0.75协同水平,A)及差异脑地形图(0.75协同水平:控制-应激,B)Figure 6.The Grand Average Waveform of P2(0.75 coherent level,A)and Topographic Map(0.75 coherent level,B)at Different Stress Levels
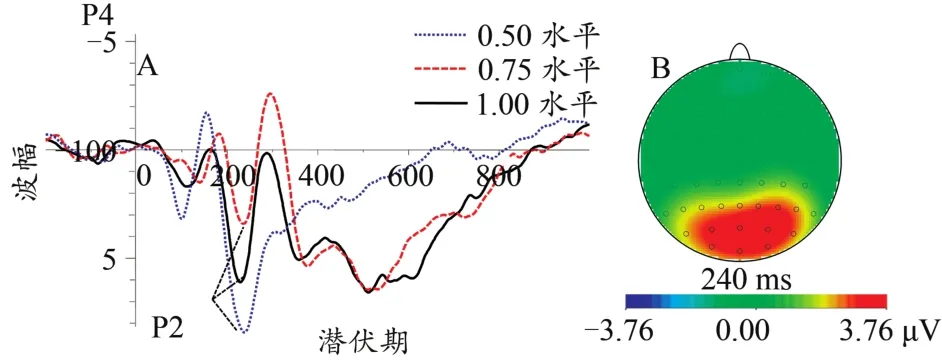
图7 不同协同性水平下P2总平均波形图(A)及差异脑地形图(0.50~1.00水平,B)Figure 7.The Grand Average Waveform of P2(A)and Topographic Map(0.50 to 1.00 coherent level,B)at Different Coherent Levels
N2成分:本研究发现,0.50水平协同运动刺激并未诱发出明显的N2成分,这与前人研究结果具有一致性,即协同性水平较低的运动未能诱发可观察到的视觉诱发电位(Niedeggen et al.,1999;Patzwahl et al.,2000)。因此,本研究纳入高协同性水平运动(0.75与1.00水平)进行重复测量方差分析,并进一步探究应激条件与非应激条件下协同运动知觉的差异机制。
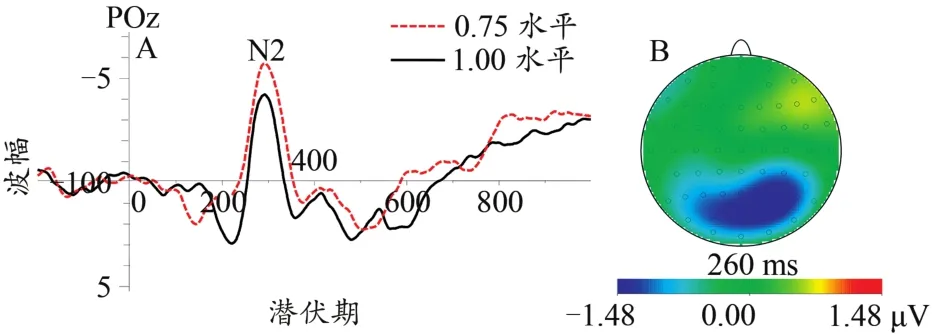
图8 不同协同性水平下N2总平均波形图(A)及差异脑地形图(0.75~1.00水平,B)Figure 8.The Grand Average Waveform of N2(A)and Topographic Map(0.75 to 1.00 coherent level,B)at Different Coherent Levels
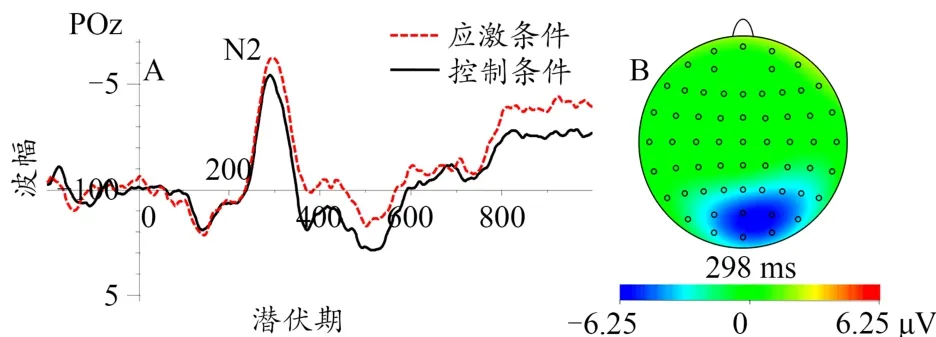
图9 不同应激条件下N2总平均波形图(A)及瞬时脑地形图(应激条件与0.75协同水平,B)Figure 9.The Grand Average Waveform of N2(A)and Instant Topography Map(stress condition and 0.75 coherent level,B)at Different Stress Levels
晚期正成分(LPP/400~600 ms):由于23名被试中有2名被试的EEG数据中LPP的平均值小于0 μV而被剔除,故本部分数据分析共纳入21名被试(10男,11女,平均年龄为19.76岁)。对于LPP而言,应激水平主效应显著,,进一步事后检验发现,应激水平下的LPP平均波幅(3.13±0.57)μV要显著小于控制条件(5.21±0.65)μV(P<0.05,图10A);协同性水平主效应显著=4.98 μV,M1.00=4.83 μV,进一步做事后检验发现,0.50水平协同运动的LPP平均波幅均要显著小于0.75水平(P<0.01)与1.00水平(P<0.05)的协同运动刺激;电极位置主效应显著,进一步做事后检验发现,P3电极点LPP平均波幅显著大于P5电极点(P<0.01),P4电极点LPP平均波幅显著大于P6电极点(P<0.05);电极位置与协同性水平交互作用显著,进一步做简单效应分析表明,在0.75水平协同运动上P5电极点LPP平均波幅显著小于P3与P4电极点(P<0.01)以及P4电极点LPP平均波幅显著大于P6电极点(P<0.05),在1.00水平协同运动上P4电极点LPP平均波幅显著大于P5与P6电极点(P<0.05)以及P3电极点LPP平均波幅显著大于P5电极点(P<0.01),在P4与P6电极点上0.50水平协同运动的LPP平均波幅显著小于0.75水平与1.00水平协同运动(P<0.01);其余两两之间交互作用均不显著(P<0.05)。
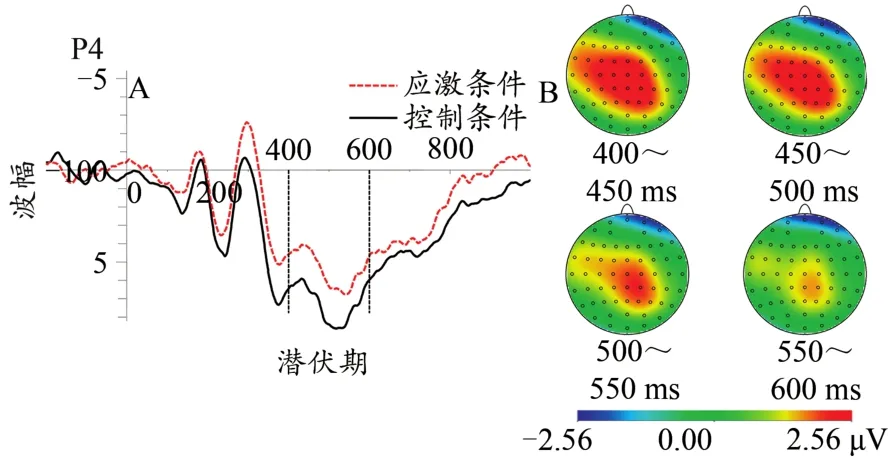
图10 不同应激条件下LPP总平均波形图(0.75协同性水平,A)及差异脑地形图(控制-应激,B)Figure 10.The Grand Average Waveform of LPP(0.75 Coherent level,A)and Topographic Map(B)at Different Stress Levels
3 讨论
本研究旨在通过双任务范式探究急性心理应激对协同运动知觉的影响及其机制的ERP特征,乘法估算任务(包括不可控制性与社会威胁性)作为先行任务,用于诱发被试的急性心理应激状态;协同运动知觉中RDK光点任务作为后行任务,用于探测被试的协同运动知觉能力大小。研究发现,乘法估算任务成功地诱发了被试的急性心理应激反应,同时急性心理应激状态下被试在协同运动知觉任务中加工速度加快、注意控制能力增强;在协同运动知觉加工早期阶段注意资源投入较早,在晚期阶段枕区表现出持续性抑制减弱现象。
在应激诱发效果方面,应激条件下的心算任务反应时要短于控制条件且正确率要小于控制条件。这与Qi等(2016)研究结果一致,即应激状态打破了被试的速度准确性之间的平衡状态,在应激与控制条件中被试可能采用了两种认知加工策略,即应激条件下更倾向于速度优先型,而控制条件下更倾向于准确率优先型。在ERP指标方面,应激条件下的P2成分峰波幅小于控制条件。Bertsch等(2011)研究发现,个体注意加工过程中若出现反应较快且准确率下降,则P2成分的波幅值会表现出下降的现象。Qi等(2016)采用乘法估算任务诱发被试的应激状态,发现相比较于控制条件,应激条件下诱发的P2峰波幅值也小于控制条件。这些研究结果与本研究基本一致,说明P2成分与注意加工过程有紧密关联,即在视觉任务中若注意资源分配增加则会造成P2波幅值的显著增加(Lenartowicz et al.,2014;Löw et al.,2015)。此外,有研究发现,应激对个体的注意加工过程具有削弱功能,即影响了注意资源的合理分配(Dambacher et al.,2014;Sänger et al.,2014)。因此,本研究采用改良后的MIST任务诱发被试的急性心理应激状态,并辅以时间压力、社会评价压力以及任务绩效压力等多重压力下削弱了被试将更多的注意资源投入到乘法估算任务。结合前人研究结果,可以发现本实验中乘法估算任务较为成功地诱发了被试的急性心理应激状态。
在急性心理应激对协同运动知觉影响的行为学层面,应激与控制条件下随着协同性水平的增加,被试的反应时逐渐缩短而正确率逐渐升高,支持了研究假设。这一结果与Robertson等(2014)的研究基本一致,其研究发现,随着协同性水平的升高,自闭症患者与正常人在协同性知觉任务中表现出的正确率逐渐增加且反应时逐渐缩短。Britten等(1992)将协同性水平分为1%、10%、100%,发现随着协同性水平的增高,被试的判断正确率也随之升高。Siegel等(2007)的研究也发现了这一类似现象。可见,随着协同性水平的增高,被试越容易做出判断,进而会降低任务反应时与增加正确率。相比较于控制条件,应激条件下被试协同性知觉判断的反应时较短,支持了假设。Dambacher等(2014)研究发现,时间压力下个体的注意焦点缩小,进而影响有机体的知觉加工过程的效率,表现出速度快而正确率较低的现象。Bertsch等(2011)研究发现,有机体在应激状态下知觉加工的速度加快,但是其准确性会有所下降。因此,在本实验应激条件下被试协同性知觉判断的反应时缩短,说明急性心理应激提升了有机体警觉性水平,缩小了注意范围,进一步促进了视觉信息内在辨别的过程,加快了有机体快速做出反应,即急性心理应激状态下有机体的自身适应性行为反应。
在急性心理应激对协同运动知觉影响的ERP特征方面:对P1成分而言,本研究发现,0.50水平协同运动的P1成分出现时间早于0.75与1.00水平,左侧枕颞叶中0.50水平协同运动诱发的P1成分出现时间慢于右侧枕颞叶,以及0.50水平协同运动的P1成分波幅值大于0.75与1.00水平。前期研究发现,P1成分是一种与感觉分析器对外在物理刺激进行早期视觉信息加工有关的成分,即与视觉外在信息刺激的初级特征编码有关,同时P1成分还与个体对无意识信息感觉偏向的抑制相关(Luck et al.,2011),且它较易受到外在刺激的对比度与亮度大小的影响(Kubová et al.,1995)。外在刺激信息进入视觉系统后,感觉分析器首先会对物理刺激的形状、颜色等视觉特征进行分析加工,这种加工属于较低水平的加工过程,主要位于颞枕区。在P1潜伏期方面,0.50水平协同运动较早地诱发出了有机体的P1成分,从3种协同运动知觉任务来看,0.50水平协同运动的任务难度较难,在屏幕上随机运动的点较多,更容易吸引有机体的注意力,故被试需要过早地分配注意资源去辨识其共同运动点的方向。此外,由于相较于0.75水平与1.00水平协同运动,0.50水平协同运动的辨识过程较长,任务难度较大,被试需要分配更多的注意资源去辨识出相应协同运动点的方向,所以0.50水平下被试的P1成分波幅值较大。
在P2成分方面,本研究发现,随着协同性水平的升高,P2成分的潜伏期逐渐缩短。在视觉任务中若注意资源分配增加则会造成P2波幅值的显著增加(Lenartowicz et al.,2014)。P2成分还与有机体工作记忆有紧密关系的ERP成分(Finnigan et al.,2011)。工作记忆也称为短时记忆,涉及模式识别,即将外在刺激信息与长时记忆储存的信息进行匹配并找出最佳的匹配选项。也就是说,随着协同性水平的增高,协同方向的判断变得更加容易,被试可以较快地进行模式识别,并与脑海中所储存的方向模式进行匹配,最后快速做出判断反应,所以P2成分的潜伏期会随着协同性水平的增加而逐渐缩短。此外,0.50水平协同运动的P2波幅值要大于0.75与1.00水平,且0.75水平协同运动的P2波幅值要小于1.00水平。P2成分的波幅值大小与个体的注意资源分配具有一定的关系,0.50水平协同运动的方向识别需要被试付出较多的认知资源,故P2波幅值较大。相比较于控制条件,应激条件下协同运动知觉的P2成分出现较早。研究发现,急性心理应激可以使个体处于高唤醒与高警觉的精神状态(Qi et al.,2020;Wang et al.,2005),心理应激会使有机体的感觉信息输入与早期视觉加工过程变得更加敏感(Davis et al.,2001)。可见,心算任务诱发了被试的急性心理应激状态,个体在心算任务中的高警觉状态延续到了协同运动知觉任务中,高警觉状态促使有机体集中注意力于当前任务,故应激条件下被试会过早地调用注意资源来辨别协同运动方向。
对于N2成分而言,本研究发现,POz电极点上的N2波幅值最大,0.75水平协同运动的N2波幅值要显著大于1.00水平。有研究发现,N2是运动特定的一种成分,与早期P1成分(与模式特点识别有关)不同,其关键区别在于运动特定机制捕获关于运动方向的信息,而不仅仅是亮度的动态变化信息(Clifford et al.,2002)。Kuba等(2007)在述评运动刺激诱发的脑电位活动特征中发现枕叶存在两个与运动有关的ERP成分:其一是运动开始后约130 ms的正成分P1,其二是150~200 ms的负成分N2。从这两项研究来看,N2成分可以作为运动刺激诱发的一个电位活动。关于N2的脑区,Meier等(2018)研究发现,对运动刺激比较敏感的背部流区域主要包括V3A、背侧枕叶沟中的V6以及沿着顶内沟的前部和下部区域。可见脑部枕叶参与运动刺激辨别的认知过程。在协同运动知觉任务中,Niedeggen等(1999)在协同运动知觉任务中发现N2出现于枕叶电极,且其潜伏期约在300 ms达到峰值,伴随着与知觉决策有关的正向偏移慢成分。研究发现,协同运动知觉任务诱发的N2成分波幅值与协同性水平有一定关系,且协同性水平较低的运动未能诱发可观察到的视觉诱发电位(Niedeggen et al.,1999;Patzwahl et al.,2000)。这些研究表明,或许由于刺激物理信息的原因,协同运动知觉过程中会在枕叶出现一个较晚的N2成分,且波幅值与协同性水平有关,同时较低的协同性水平未能诱发出视觉诱发电位,这与本研究结果基本一致。Manning等(2019)采用ERP技术研究发现,被试在协同运动知觉任务中会在枕区中出现潜伏期约300 ms的一个负成分,如同前期研究中有关协同运动知觉诱发的N2成分。可见,协同性知觉任务中枕叶区域确实存在一个潜伏期约300 ms的N2成分。除了N2成分代表运动特征信息外,Olson等(2016)认为,N2成分是有关注意控制的策略认知控制过程中的一种ERP成分。这说明,相较于1.00水平协同运动,0.75水平的辨别过程需要更多的注意控制能力,这是由于0.75水平的难度要比1.00水平大所造成的。此外,本研究还发现,相较于控制条件,应激条件中0.75水平协同运动知觉诱发的N2波幅值较大,支持了假设。研究发现,心理应激会促使个体的唤醒水平与警觉性水平升高(Qi et al.,2020;Wang et al.,2005),而焦虑与紧张状态会让个体的注意控制能力得到进一步加强(Hum et al.,2013;Righi et al.,2009)。0.75水平属于难度适中的状态,同时也需要被试具有较好的注意控制能力,易受到焦虑状态的影响。因此,应激条件下0.75水平协同运动知觉任务过程中被试的注意控制能力较好。
关于LPP,本研究发现,相较于控制条件,应激条件下协同运动知觉的LPP平均波幅值较小。研究发现,LPP成分通常在人们大脑后部(枕区)出现(Hajcak et al.,2010),其中内源性LPP成分代表着有机体的高级信息加工阶段,它和个体的工作记忆更新密切相关。也有研究提出,LPP成分反映了大脑视觉皮层活动的全局抑制能力,与情绪刺激加工活动的选择密切相关,它是衡量情绪调节的重要指标之一(Thom et al.,2014)。前期研究发现,应激条件下有机体大脑警觉性升高,注意力范围缩小,被试需要付出更多的认知资源来应付应激环境,故在协同运动知觉判断任务中注意资源会受到应激状态的影响,进而造成大脑中用来参与协同运动知觉任务的认知资源减少,即大脑在协同运动知觉任务中抑制持续性较弱。而在控制条件下,LPP波幅较大,大脑用于协同运动知觉任务中认知资源较多,即大脑在该部分的抑制持续性较好。本研究还发现,右侧枕颞叶上0.50水平协同运动的LPP平均波幅要小于0.75水平与1.00水平。相较于0.75与1.00水平,0.50水平协同运动的任务难度较大,被试不仅要付出较多认知资源应对应激环境,同时还需要付出较多努力去判断协同方向,故其0.50水平协同运动中的抑制能力较低。
结合行为学数据与脑电数据的分析来看,相比较于控制条件,应激状态下协同运动知觉任务表现较好,可以通过较低的反应时来体现。同时,与控制条件相比,应激状态下协同运动知觉任务诱发的P2成分出现时间较早和N2波幅值较大,说明应激状态下早期注意资源投入较早以及注意控制增强。综合行为学和神经生理学结果来看,可以推断出应激条件下协同运动知觉任务反应时缩短与P2、N2成分的活动高度相关。这与前期的研究结果基本一致,即心理应激会促使有机体感觉信息输入和早期知觉加工更加迅速(Davis et al.,2001),同时紧张状态会使个体的注意控制能力增强(Hum et al.,2013;Righi et al.,2009),进而体现为有机体在协同运动知觉任务中的反应速度加快。随着协同性水平的增加,协同运动知觉任务中P2成分的潜伏期逐渐缩短,体现在行为学层面上为协同运动知觉任务反应速度加快。这一结果也表明,协同运动知觉任务反应速度大小与P2成分活动情况相关,即注意资源的调用速度与行为反应速度呈现一定程度的正相关。可见,本研究行为学结果与脑电数据结果具有一定的相关性。
通常认为急性心理应激是“有害”的,但本研究却发现,急性心理应激对协同运动知觉的影响并非如此。研究从行为学层面发现,急性心理应激促进了协同运动知觉任务的操作表现。这一结果符合了双竞争理论模型(Pessoa,2009),即两种任务之间产生了影响。同时该结果也符合倒U型假设模型(Yerkes et al.,1908),即适度的心理唤醒水平有利于个体的行为表现。此外,不论是ERP早成分还是晚期正成分,均从侧面说明急性心理应激增强了人们在协同运动知觉任务的注意控制能力,以及早期注意资源的投入。可见,急性心理应激提高了个体警觉性水平(Wang et al.,2005),促进了视觉信息内在辨别的加工过程(Davis et al.,2001)。也就是说,人们在急性心理应激状态下的知觉加工过程表现出了较好的自适应状态。据此,人们可依据知觉刺激的运动场景做出适时的情绪调整,达到最佳的知觉-情绪-行为状态。
4 结论
1)乘法心算任务成功地诱发了急性心理应激反应且削弱了后期注意资源分配。2)协同性水平对协同运动知觉加工具有积极效应,注意效率更高。3)急性心理应激状态下协同运动知觉加工速度加快、注意控制能力增强;在协同运动知觉加工早期阶段注意资源投入较早,在晚期阶段枕区表现出持续性抑制减弱现象。
猜你喜欢
新高考·高二数学(2022年3期)2022-04-29
兵工学报(2022年1期)2022-03-14
汽车工程师(2021年12期)2022-01-18
高考·中(2019年6期)2019-09-10
价值工程(2018年25期)2018-09-26
中国现代医生(2017年24期)2017-10-17
小学阅读指南·低年级版(2016年10期)2016-09-10
科技视界(2016年6期)2016-07-12
求学·理科版(2015年10期)2015-11-04
股市动态分析(2014年48期)2014-12-18
