
2016—2017 年河北正定县、湖南湘潭县及浙江玉环市轮状病毒VP7 基因的分子流行病学特征
2021-04-23 11:20陈红李庆亮段凯石晨张东董犇白萱乔建徐葛林杨晓明高招李放军吕华坤周海松鄢廷栋施海云
中国生物制品学杂志 2021年4期
陈红 ,李庆亮 ,段凯 ,石晨 ,张东 ,董犇 ,白萱 ,乔建 ,徐葛林 ,杨晓明,高招,李放军,吕华坤,周海松,鄢廷栋,施海云
1.武汉生物制品研究所有限责任公司,湖北武汉430207;2.中国生物技术股份有限公司,北京100029;3.河北省疾病预防控制中心,河北石家庄050000;4.湖南省疾病预防控制中心,湖南长沙410005;5.浙江省疾病预防控制中心,浙江杭州310051;6.正定县疾病预防控制中心,河北正定050800;7.湘潭县疾病预防控制中心,湖南湘潭411200;8.玉环市疾病预防控制中心,浙江玉环317600
轮状病毒(rotavirus,RV)感染是导致全球 5 岁以下儿童腹泻相关死亡病例的主要原因之一,2016年全球由于RV 感染引起的5 岁以下儿童死亡病例数达128 500 例,其中撒哈拉以南的非洲、东南亚和南亚的死亡率最高[1]。RV 属于呼肠病毒科轮状病毒属,1973 年,通过电镜首次观察到腹泻患儿粪便中的RV[2]。成熟的RV 颗粒为对称的二十面体球形,无包膜,病毒颗粒直径约75 nm。基因组为11 段dsRNA,编码 6 种结构蛋白和 6 种非结构蛋白[3],其中基因片段9 编码的表面蛋白VP7 是RV 主要的中和抗原之一。根据VP7 基因核苷酸序列的相似性和差异性,RV 分为不同的G 基因型,目前至少定义了 36 种 G 基因型[4]。
RV 易发生变异,目前,G1-G4、G9 和 G12 是全球主要的优势流行株[5]。监测结果显示,2011—2014 年中国26 个省(自治区、直辖市)5 岁以下儿童RV 感染腹泻病例中,G9 型占比不断增加,G3、G1 及 G2 型占比逐年降低[6]。王芳[7]调查发现,2004 年前,兰州市主要流行G 基因型为G3,2004 — 2005 年转变为 G2,2005 — 2007 年为 G1,2007 — 2012 年间 G3再次成为流行基因型,2013 年后流行基因型为G9。ZENG 等[8]对 2011 — 2015 年重庆市婴幼儿门诊腹泻病例样本分析显示,G 基因型优势流行株从G1 变为G9,2013 年以后G9 检出率逐渐增加。TINA等[9]对 2011 — 2016 年北京市 5 岁以下儿童门诊腹泻病例基因型的分析表明,2012 年前G3 为优势流行株,2012 年后为G9。基因的变异和进化导致其抗原发生漂移和转移,影响病毒的传播和流行[10]。
目前,RV 感染引起的儿童急性胃肠炎无特异性抗病毒治疗药物,疫苗是控制该疾病的有效手段[11]。国际上广泛应用的口服RV 疫苗主要是Rotarix 和RotaTeq,RotaTeq(WC3 株)为五价人-牛重配口服减毒活疫苗,包含G1 ~G4 的4 株VP7 基因重配株及1株 P[8]的 VP4 基因重配株;Rotarix 为单价口服减毒活疫苗,其VP7 基因为G1。2007 年,武汉生物制品研究所有限责任公司从国外引进六价UK-人重配疫苗株毒种用于RV 疫苗的生产,其VP7 基因为G1、G2、G3、G4、G8 和 G9。本研究收集了 2016 — 2017 年间河北正定县、湖南湘潭县及浙江玉环市2 岁以下儿童腹泻监测的粪便样本,对RV 阳性样本进行测序分型,通过系统发育进化树等分析方法,探讨RV VP7 基因的分子流行病学特点,比较UK-人重配疫苗毒株与3 个地区流行株VP7 基因序列同源性和进化关系,以期为UK-人重配疫苗在国内的应用提供相应的参考依据。
1 材料与方法
1.1 样本 收集 2016 年 11 月 — 2017 年 4 月间河北正定县、湖南湘潭县及浙江玉环市2 岁以下儿童腹泻病例RV 阳性粪便样本(经ELISA 试剂盒检测),共 228 份。
1.2 主要试剂 病毒RNA 提取试剂盒QIAamp Viral RNA Mini Kit 购自美国Qiagen 公司;无水乙醇购自国药集团化学试剂有限公司;RT-PCR 试剂盒Prime-ScriptTMOne Step RT-PCR Kit Ver.2 及DNA marker DL2000 购自日本TaKaRa 公司;琼脂糖购自上海贝晶生物技术有限公司;DEPC 水购自上海碧云天生物技术有限公司。
1.3 粪便样本处理 将0.1 mL 粪便样本置装有0.5 mL DEPC 水的1.5 mL 离心管中,获得10% ~20%的粪便悬浮液,旋涡振荡 1 min,2 823 × g 离心5 min;收集上清,于-30 ℃保存。
1.4 VP7 基因的扩增 根据GenBank 中登录的人RV VP7 基因(M21843、Q650124.1、EF672602.1、LC-482504)序列,应用Primer Premier 5 软件设计特异性引物,VP7 F:5′-GGCTTTAAAAGAGAGAATTTCCGTCTGG-3′,VP7 R:5′-GGTCACATCATACAATTCTAATCTAAG-3′,扩增产物大小为1 062 bp。引物由生工生物工程(上海)股份有限公司合成。采用QIAamp Viral RNA Mini Kit 提取粪便上清液中的病毒RNA,以其为模板进行RT-PCR 扩增。PCR 体系为:PrimeScript 1 Step Enzyme Mix 2 μL,2 × 1 Step Buffer 25 μL,VP7 F /VP7 R 各 1 μL,提取的样本 RNA 8 μL,RNase Free dH2O 13 μL。PCR 反应条件为:50 ℃30 min,94 ℃ 2 min;94 ℃ 30 s,50 ℃ 30 s,72 ℃ 1 min,共 35 个循环;72 ℃ 5 min,4 ℃终止反应。PCR 产物经2%琼脂糖凝胶电泳分析。同时以DEPC 水为阴性对照,试剂盒中的Positive RNA 为阳性对照。
1.5 VP7 基因测序分析 取20 μL PCR 产物送生工生物工程(上海)股份有限公司测序,测序引物为VP7F / VP7R。测序结果应用Sequencher 软件拼接处理,再经RV 在线基因分型工具RotaC V2.0 对测序结果进行分型[12],Mega X 或 Megalign 软件进行比对,分析序列相似性。
1.6 VP7 基因系统进化树及氨基酸序列分析 从GenBank 中下载RV 的VP7 基因国内外近期流行株各型别原型株及不同谱系标准株序列作为参考序列,应用Mega X 软件(http://www.megasoftware.net /)[13]对测序结果与参考序列进行ClustalW 多重比对,构建Neighbor-Joining 进化树,通过Kimura-2-parameter模型测量系统发育距离,Bootstrap 对进化树的可靠性进行评价,重复1 000 次。应用Mega X 软件将核苷酸序列翻译为氨基酸序列,分析原型株(G1、G2、G3、G9 型原型株分别为 Wa、DS-1、P、WI61)及 UK-人重配疫苗毒株(G1、G2、G3 及 G9 的 GenBank 登录号为GQ225777、GQ225778、GQ225827、GQ225782)和 3 个地区样本VP7 基因氨基酸序列差异性。RV VP7 基因可变区氨基酸序列有VR1(9 ~20 位氨基酸)、VR2(25 ~ 32 位氨基酸)、VR3(37 ~ 53 位氨基酸)、VR4(65 ~ 76 位氨基酸)、VR5(87 ~ 100 位氨基酸)、VR6(119 ~ 132 位氨基酸)、VR7(141 ~ 150 位氨基酸)、VR8(208 ~ 224 位氨基酸)和 VR9(235 ~ 242 位氨基酸)[14],VP7 中和抗原表位主要位于 A(87 ~ 100 位氨基酸)、B(141 ~ 150 位氨基酸)和 C(208 ~ 224 位氨基酸)3 个区域内,即 VR5、VR7 和 VR8 的位置[15]。
2 结 果
2.1 VP7 基因扩增产物的鉴定 共有199 份样本扩增并成功测序。RV VP7 基因片段PCR 产物经2%琼脂糖凝胶电泳分析,可见1 062 bp 的目的基因条带,大小与预期一致,部分结果见图1。
2.2 VP7 基因测序分析 测序结果共有4 种G 基因型,即 G1、G2、G3、G9 型,所占比例分别为 10 / 199(5.1%)、43 /199(21.6%)、12 / 199(6.0%)、134 / 199(67.3%)。其中,正定县优势流行株为G2,占77.4%;湘潭县和玉环市优势流行株均为G9,分别占75.8%和75.5%。见表1。
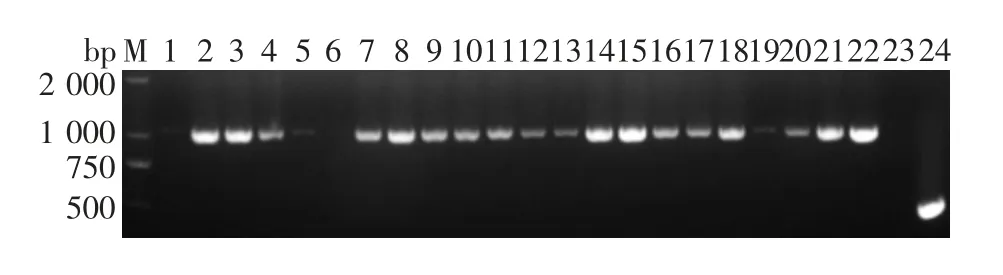
图1 部分粪便样本VP7 基因扩增产物电泳图Fig.1 Electrophoretic profile of PCR products of VP7 gene of partial stool samples
2.3 RV VP7 基因序列特征 3 个地区的测序结果中,G1 型 RV 共 10 份,VP7 核苷酸序列相似性为93.2% ~100.0%,氨基酸序列相似性为96.0% ~100.0%;G2 型 RV 共 42 份,VP7 核苷酸序列相似性为85.4% ~100.0%,氨基酸序列相似性为88.1% ~100.0%;G3 型 RV 共 12 份,VP7 核苷酸序列相似性为81.2% ~100.0%,氨基酸序列相似性为91.4% ~100.0%;G9 型 RV 共 133 份,样本 VP7 核苷酸序列相似性为78.1% ~100.0%,氨基酸序列相似性为72.8% ~100.0%。G1 型RV 样本间核苷酸序列差异较小,G9 型RV 样本间的核苷酸序列差异性相对较大,部分结果见表2 和表3。

表1 3 个地区VP7 基因测序分型结果Tab.1 Sequencing and typing of VP7 gene in three regions
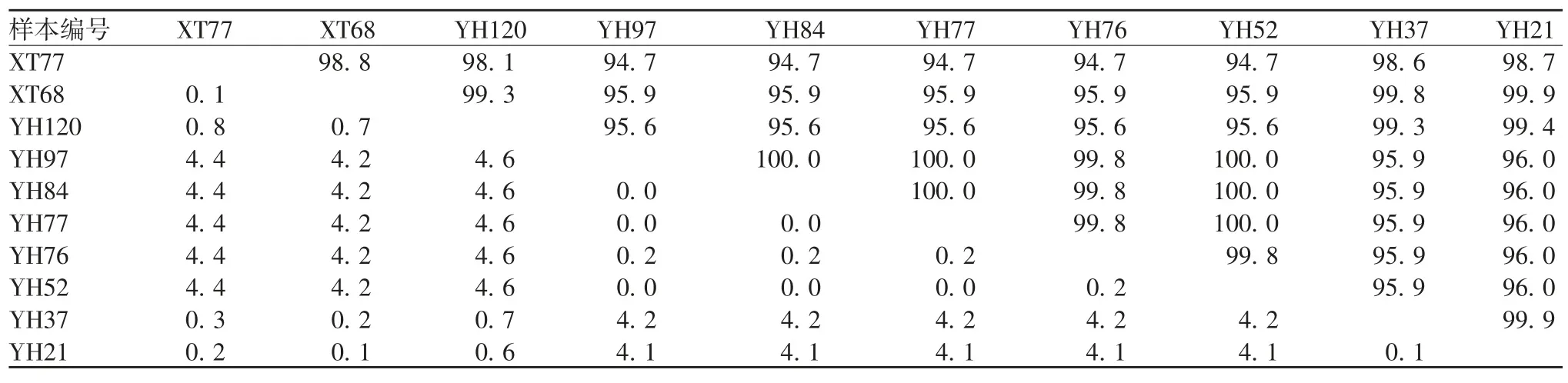
表2 G1 型样本核苷酸序列相似性(%)Tab.2 Homology of nucleotide sequences of samples of G1 genotype(%)
2.4 各型RV 与UK-人重配株VP7 基因进化分析
2.4.1 G1 型 10 株 G1 型 RV 均集中在 G1-Ⅰ亚型,分属G1-Ⅰ亚型的2 个不同分支,其中玉环市的YH76、YH97、YH84、YH77 和 YH52 5 株处于同一进化分支,其他 5 株:XT77、XT68、YH21 和 YH37 处于同一进化分支。UK-人重配疫苗毒株G1 与G1 型原型株 Wa(1974,USA)处于 G1-Ⅲ进化亚支,亲缘关系较近。10 株G1 型分离株均同G1 型RV 原型株Wa 的VP7 基因的进化距离较远,处于不同的进化树分支,见图2。G1 型分离株与Wa 株RV 核苷酸序列相似性为90.6% ~91.9%,与G1 型UK-人重配疫苗毒株核苷酸序列相似性为90.4% ~91.7%。
2.4.2 G2 型 43 株 G2 型 RV 分属于 G2 Ⅲ-2 和Ⅲ-3 两个亚支,其中 ZD24、YH34、XT23 和 XT14 均在Ⅲ-2 亚支,但在G2 Ⅲ-2 的不同分支;其他39 株属于Ⅲ-3 亚支。UK-人重配疫苗毒株G2 与G2 型原型株 DS-1(1976,USA)处于 G2-Ⅰ进化亚支,亲缘关系较近。43 株 G2 型 RV 均与原型株 DS-1 的 VP7 基因的进化距离较远,处于不同的进化树分支,见图3。G2 型RV 分离株与DS-1 株RV 核苷酸序列相似性为83.5% ~94.0%,与G2 型UK-人重配疫苗毒株核苷酸序列相似性为83.5% ~94.0%。

表3 G1 型样本氨基酸序列相似性(%)Tab.3 Homology of amino acid sequences of samples of G1 genotype(%)
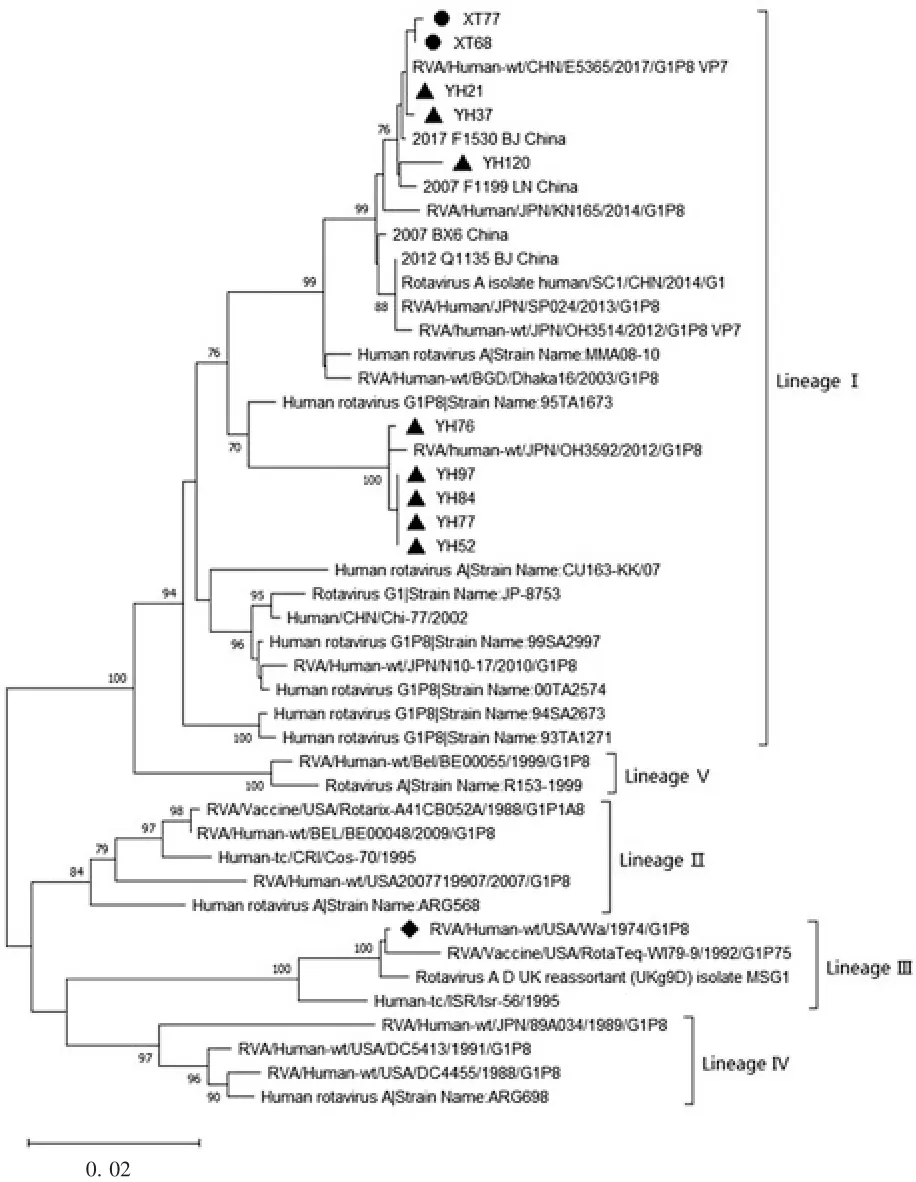
图2 G1 型RV 的VP7 基因核苷酸序列进化分析Fig.2 Phylogenetic analysis of nucleotide sequence of VP7 gene of genotype G1

图3 G2 型RV 的VP7 基因核苷酸序列进化分析Fig.3 Phylogenetic analysis of nucleotide sequence of VP7 gene of genotype G2
2.4.3 G3 型 12 株 G3 型 RV 分属于 G3-Ⅳ和 G3-Ⅴ两个不同亚支,其中玉环市的 7 株(YH53、YH4、YH83、YH116、YH117、YH31和YH44)属于G3-Ⅴ亚型,其他 5株属于G9-Ⅳ亚型。UK-人重配疫苗毒株G3 与G3型原型株 P(1974,USA)处于 G3-Ⅰ进化亚支,亲缘关系较近。12 株 G3 型 RV 均与原型株 P(1974,USA)的VP7 基因的进化距离较远,处于不同的进化树分支,见图4。G3 型分离株与P 株RV 核苷酸序列相似性为82.6% ~97.2%,与G3 型UK-人重配疫苗毒株核苷酸序列相似性为82.5% ~97.1%。
2.4.4 G9 型 134 株 G9 型 RV 分属于 G9-Ⅲ和G9-Ⅵ两个不同亚支,其中包括正定县所有7 株样本在内共73 株G9 型RV 属于G9-Ⅵ亚型,与中国原型株T203 处于同一亚型,其他61 株处于G9-Ⅲ亚型分支中。UK-人重配疫苗毒株与原型株WI61(1983,USA)处于相同的进化亚支G9-Ⅰ,UK-人重配株G9与AU32 株亲缘关系较近。134 株G9 型RV 均与WI61 株的VP7 基因的进化距离较远,处于不同的进化亚支。见图5。134 株G9 型样本与WI61 株RV 的VP7 基因核苷酸序列相似性为75.8% ~89.8%,与G9型UK-人重配疫苗毒株VP7 基因核苷酸序列相似性为74.8% ~89.2%。
2.5 各型RV 与原型株及UK-人重配疫苗毒株氨基酸序列差异的比较
2.5.1 G1 型 10 份 G1 型 RV 与原型株及 UK-人重配疫苗毒株比较,多个氨基酸位点存在差异,可变区共有15 个氨基酸位点存在差异。A 抗原区中存在差异的氨基酸位点有S94N、E97D 和E100D;B 抗原区中存在差异的氨基酸位点有N147S 和F148L;C抗原区中存在差异的氨基酸位点有T217M 和V218I。除抗原区外,可变区VR3、VR4 及VR9 也有部分氨基酸位点存在差异。见表4。
2.5.2 G2 型 43 份G2 型样本中,与原型株及UK-人重配疫苗毒株比较,可变区中共有12 个氨基酸位点存在差异。A 抗原区中存在差异的氨基酸位点有T87A 和 N96D,YH100 样本在第 94 位氨基酸对应的核苷酸测序峰图出现重叠峰,导致该位点碱基不能确定;C 抗原区中存在差异的氨基酸位点有D213N。其他差异出现在 VR1、VR3、VR4、VR6 和 VR9。见表5。
2.5.3 G3 型 12 份 G3 型样本与原型株及UK-人重配疫苗毒株比较,可变区共有17 个氨基酸位点存在差异。A 抗原区中存在差异的氨基酸位点有S87T和M93I;B 抗原区中存在差异的氨基酸位点有N145D;C 抗原区中存在差异的氨基酸位点有A212T,T213N。其他可变区的11 个氨基酸差异出现在VR1、VR3、VR4 和 VR6。见表6。
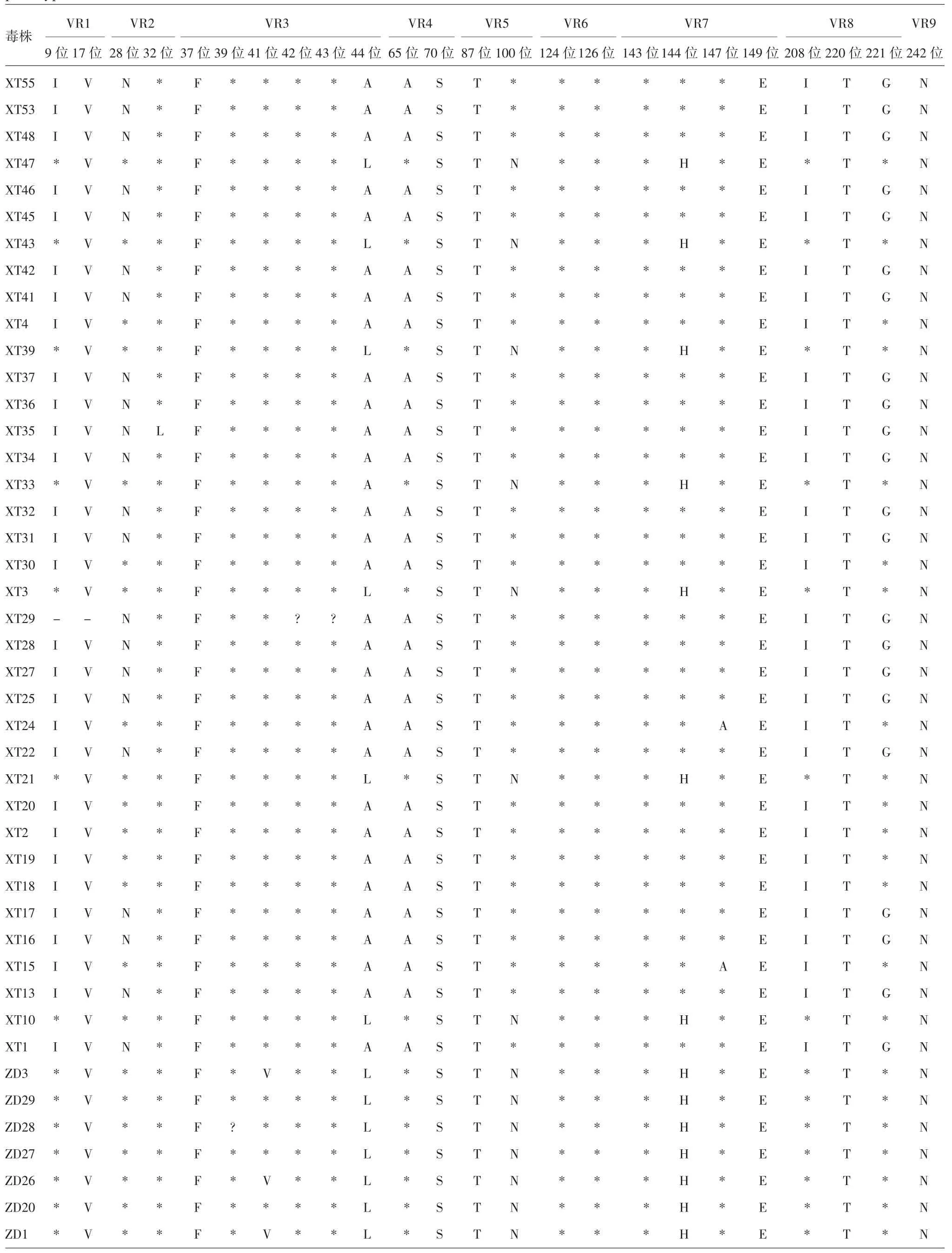
2.5.4 G9 型 134 份 G9 型 RV 与原型株及 UK-人重配疫苗毒株比较,共有32 处氨基酸位点存在差异。A 抗原区中存在差异的氨基酸位点有T87A、A90S 和N100D;B 抗原区中存在差异的氨基酸位点有H144Y、A147T 和E149K;C 抗原区中存在差异的氨基酸位点有 I208T、T220A 和 G221S,见表7 和表8。其中,aa87、aa149 和 aa220 3 个位点的分离株与疫苗株和原型株WI61 完全不同,而与国内原型株T203相同,表明国内流行的 G9 型 RV 在 aa87、aa149 和aa220 3 个位点发生了变异。其他可变区的氨基酸差异出现在 VR1、VR2、VR3、VR4、VR6 和 VR9。

图4 G3 型RV 的VP7 基因核苷酸序列进化分析Fig.4 Phylogenetic analysis of nucleotide sequence of VP7 gene of genotype G3
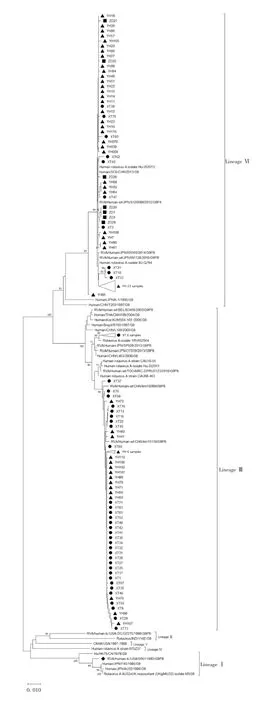
图5 G9 型RV 的VP7 基因核苷酸序列进化分析Fig.5 Phylogenetic analysis of nucleotide sequence of VP7 gene of genotype G9
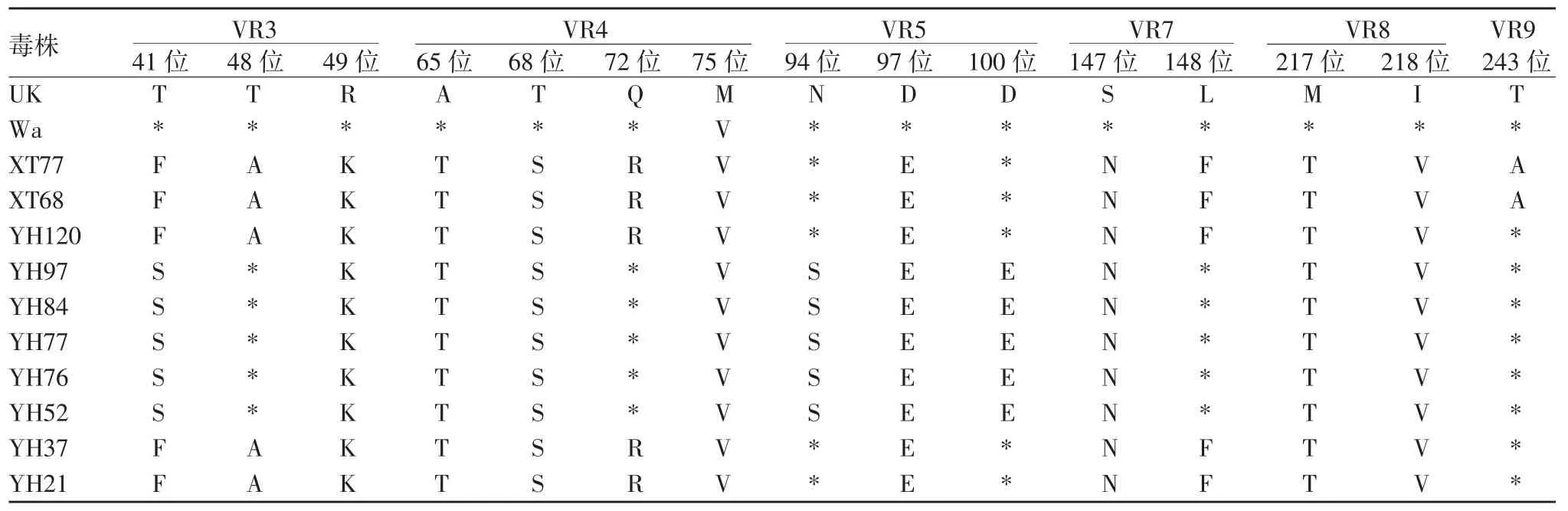
表4 G1 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸差异比较Tab.4 Difference in amino acid sequences of VP7 gene of RV of G1 genotype from prototype and UK-human reassortant vaccine virus strain
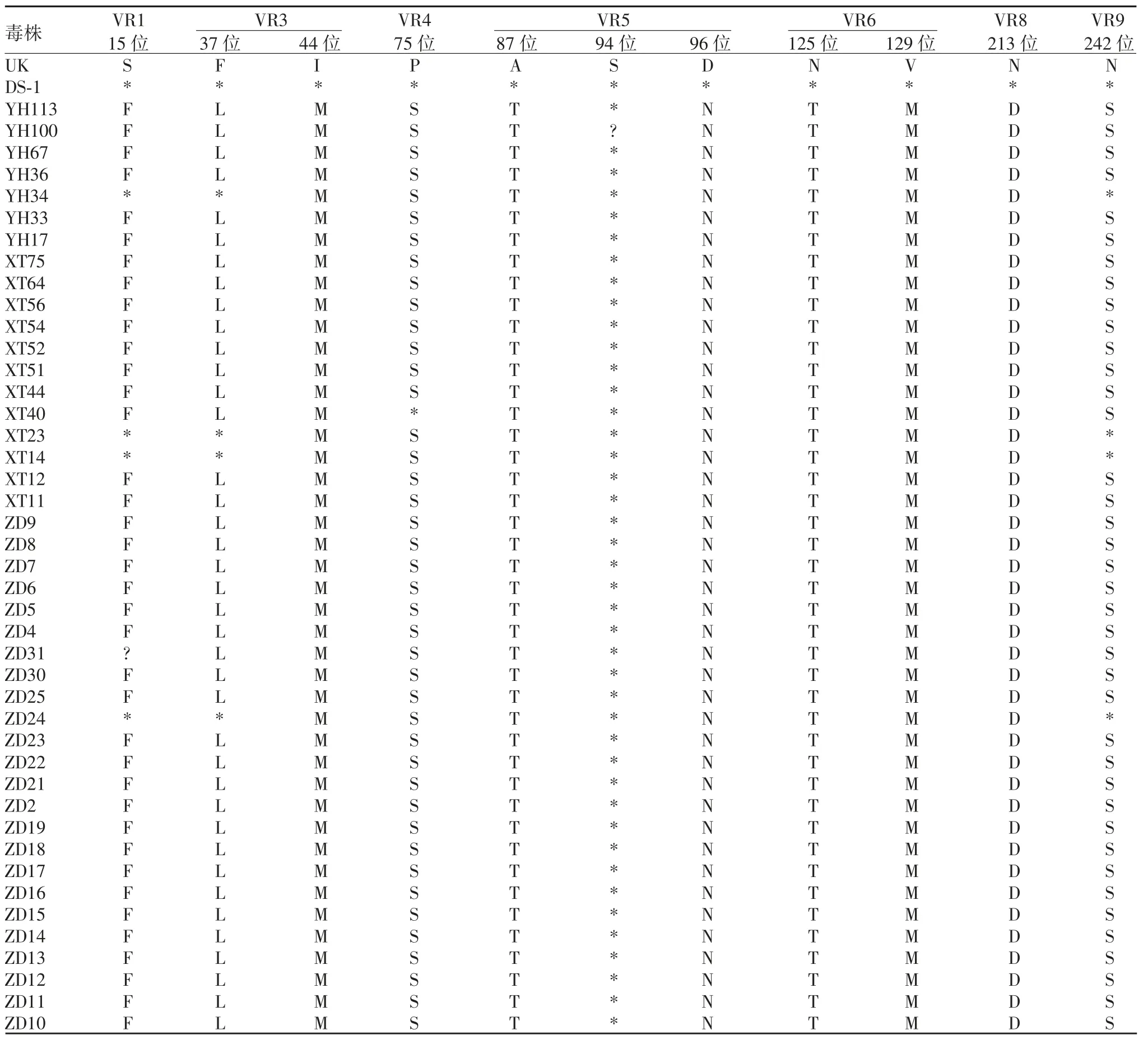
表5 G2 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸序列差异Tab.5 Difference in amino acid sequences of VP7 gene of RV of G2 genotype from prototype and UK-human reassortant vaccine virus strain
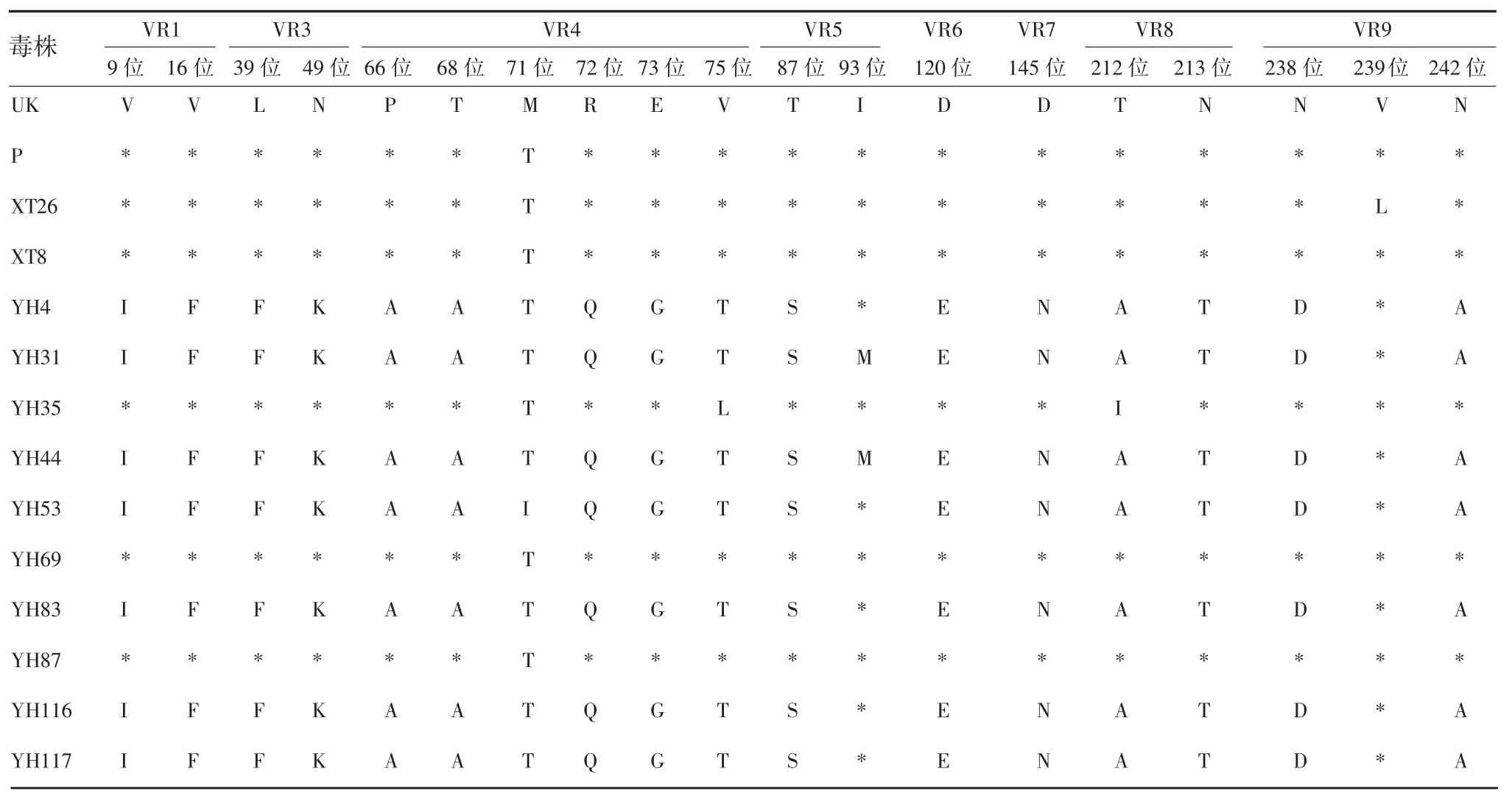
表6 G3 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸差异比较Tab.6 Difference in amino acid sequences of VP7 gene of RV of G3 genotype from prototype and UK-human reassortant vaccine virus strain

表7 正定和湘潭县G9 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸序列差异比较Tab.7 Difference in amino acid sequences of VP7 gene of RV of G9 genotype in Zhengding and Xiangtan Counties from prototype and UK-human reassortant vaccine virus strain
续表7 正定和湘潭县G9 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸序列差异比较Tab.7(Continued)Difference in amino acid sequences of VP7 gene of RV of G9 genotype in Zhengding and Xiangtan Counties from prototype and UK-human reassortant vaccine virus strain

表8 玉环市G9 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸序列差异比较Tab.8 Difference in amino acid sequences of VP7 gene of RV of G9 genotype in Yuhuan City from prototype and UK-human reassortant vaccine virus strain
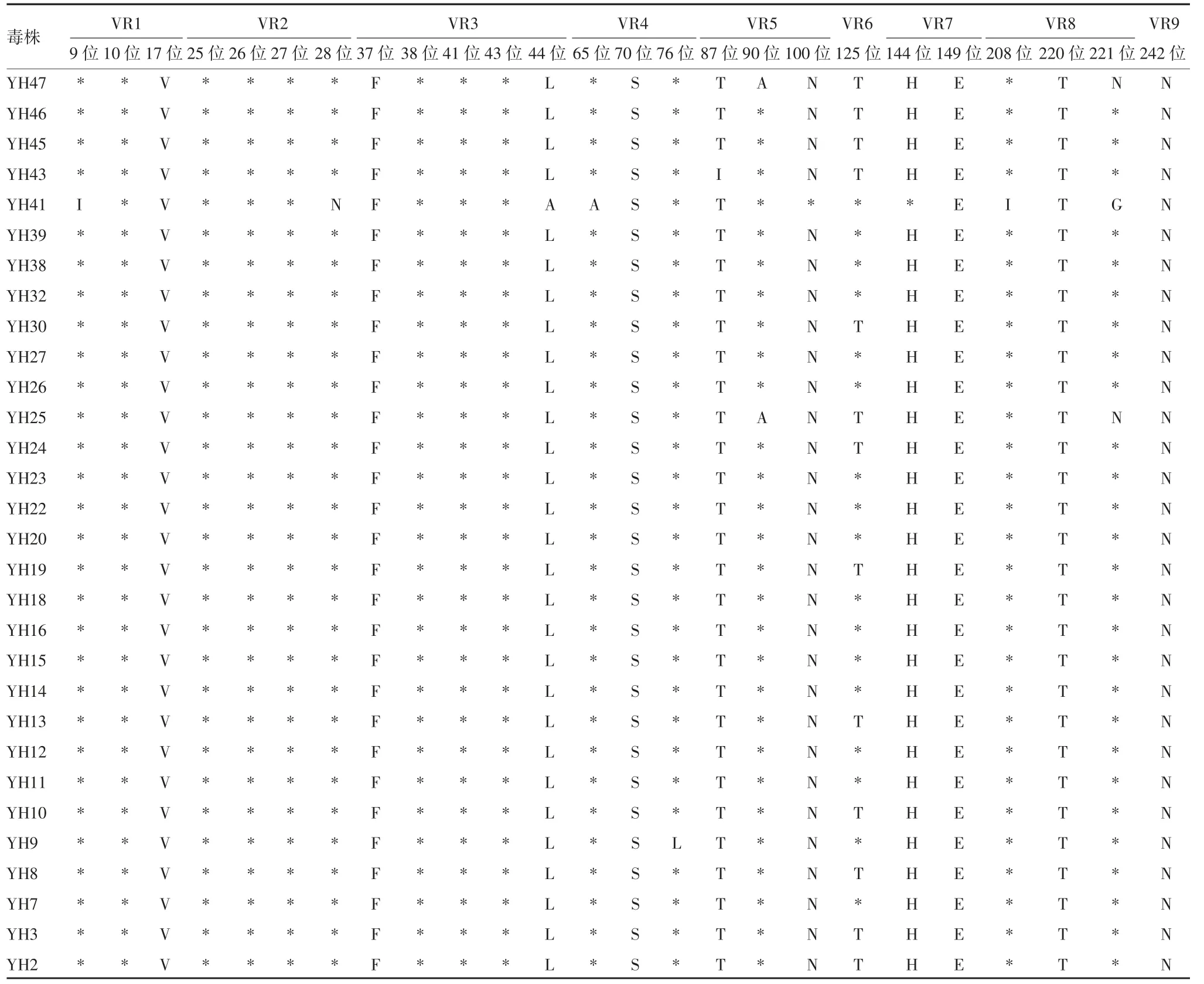
续表8 玉环市G9 型RV 的VP7 基因与原型株及UK-人重配疫苗毒株氨基酸序列差异比较Tab.8(Continued)Difference in amino acid sequences of VP7 gene of RV of G9 genotype in Yuhuan City from prototype and UK-human reas-sortant vaccine virus strain
3 讨 论
3.1 进化树分析 本研究对2016 年11 月— 2017年4 月间来自河北正定、湖南湘潭和浙江玉环3 个地区的RV 阳性粪便标本进行了VP7 基因测序及分型,确定了199 份样本的核酸序列及VP7 基因型,构建的系统进化树显示,199 份样本均与各自基因型原型株亲缘关系较远,与原型株核苷酸序列相似性在 75%以上。在 10 株 G1 型 RV 中,YH76、YH97、YH84、YH77 和 YH52 与 2012 年 的 日 本 分 离 株OH3592 亲缘关系较近,处于同一进化分支,XT77、XT68、YH21 和 YH37 与 2017 年中国分离株 E5365亲缘关系较近,YH120 与2007 年辽宁分离株F1199处于同一进化亚支;在 43 株 G2 型 RV 中,ZD24 与2005 年孟加拉国分离株MMC88 亲缘关系较近,YH34、XT23 和 XT14 与 2011 年日本B110056 株亲缘关系较近,这4 株均在G2Ⅲ-2 亚支,其他39 株属于 G2 Ⅲ-3 亚支,与 2015 年昆明株 km15028、2014 年北京株F440、2017 年日本株Tokyo17-15 及泰国2017年B3517 株和2018 年DBM2018-105 株亲缘关系较近;在 12 株 G3 型 RV 中,YH53、YH4、YH83、YH116、YH117、YH31 和 YH44 与 2016 年台湾株 08-102-689219-25-6 和 2018 年日本株 Tokyo18-25 亲缘关系较近,XT8 与2010 年中国分离株E2432 亲缘关系较近,YH69、XT26 和 YH87 与 2017 年日本分离株Tokyo17-08 株亲缘关系较近,YH35 与土耳其1RIZE-2016 株亲缘关系较近;在 134 株 G9 型 RV 中,处于G9-Ⅵ亚型的73 株与2012 年日本分离株S120088、中国分离株JS2013 及BJ-Q794 等亲缘关系较近,其他 61 株与中国分离株 km15099、km15118 及1RV-652564 等亲缘关系较近。玉环市的G3 型RV 分属于两个亚型,玉环市和湘潭县的G9 同样也分属两个不同的G9 亚型,G3 和G9 型RV 样本核苷酸与氨基酸差异性相对较大,表明在这两个地区流行株中G3和G9 流行的亚型具有多样性。
3.2 G9 型RV 流行特征 G9 型毒株最早于1983年在美国发现。在许多国家,G9 型被认为是最常见的新血清型之一,我国于1994 年首次检测到G9型毒株(T203)[16],随后几年内仅在少数地区检出,至2004 年逐渐增多,检出率也呈波动性上升趋势[17]。本研究表明,湘潭县和玉环市优势流行株均为G9,占比67.3%,为主要优势流行株。根据VP7 基因片段的核苷酸序列分析,G9 型RV 毒株分为Ⅰ ~Ⅵ 6 个分支[18],134 份 G9 样本主要分布在 G9-Ⅲ和 G9-Ⅵ两个亚型,其中属于G9-Ⅵ亚型的73 株(54.5%)RV与中国原型株T203 核苷酸序列相似性为84.2% ~95.5%,61 份(45.5%)G9-Ⅲ型样本与昆明株km15099、km15118 遗传距离较近。有研究认为,G9-Ⅵ在90年代中后期至2000 年前仅在中国和日本流行,随后国内流行的G9 型RV 主要是G9-Ⅲ,本研究表明,G9-Ⅵ占比超过G9-Ⅲ,呈现优势流行,与董慧瑾等[19]对2011 — 2012 年北京部分腹泻患儿检测优势流行株一致。结果表明,RV 的VP7 基因流行株亚型也在不断变化。
3.3 氨基酸序列差异分析 本研究结果表明,G1型RV 氨基酸序列差异较小,可变区仅有15 个氨基酸位点不同;G9 型RV 氨基酸序列差异较大,可变区有32 个氨基酸位点不同,其原因可能是由于检测出的G9 型RV 病毒较多,且第一代测序的方法会有部分样本测序结果不佳造成的。通过分析3 个抗原区氨基酸序列的变异情况发现,G2 型RV 抗原区氨基酸序列差异较小,G2 型UK-人重配株疫苗的保护效果较好;与原型株 WI61[20]比较,G9 型 RV 抗原区aa87、149 和220 3 个位点的氨基酸发生了变异。
3.4 RV 优势流行株不同地区存在差异 疫苗是保护儿童预防RV 感染的重要手段,自RV 疫苗问世以来,在保护儿童预防重症腹泻上发挥了重要作用。VP7 蛋白作为RV 关键的中和抗原之一,可诱发保护性中和抗体,目前上市的疫苗多以VP7 蛋白为主要抗原设计而来。本研究表明,正定县的优势流行株为G2,同时期的湘潭县和玉环市优势流行株则为G9,一种有效的疫苗必须包括与流行地区抗原性相同的病毒株。
综上所述,不同地区在同一时间优势流行株不同,且流行株的亚型也在发生变化,各G 血清型的RV 样本间的核苷酸序列差异性也有不同。研究可变区发生变异的氨基酸位点对发现野毒株逃逸机制具有一定意义。因此,对不同地区的RV 进行持续的监测十分必要。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
小资CHIC!ELEGANCE(2021年45期)2021-01-11
英美文学研究论丛(2018年2期)2018-08-27
剑南文学(2016年14期)2016-08-22
人间(2015年20期)2016-01-04
动物医学进展(2015年10期)2015-12-07
特产研究(2014年4期)2014-04-10
