
不同碳氮源对枯草芽胞杆菌BAB-1产抗菌脂肽的影响
2021-04-21 08:35张晓云郭庆港王培培苏振贺鹿秀云赵卫松曲远航董丽红李社增
中国生物防治学报 2021年2期
张晓云,郭庆港,王培培,苏振贺,鹿秀云,赵卫松,曲远航,董丽红,丛 蓉,李社增,马 平
(河北省农林科学院植物保护研究所/河北省农业有害生物综合防治工程技术研究中心/农业农村部华北北部作物有害生物综合治理重点实验室,保定 071000)
芽胞杆菌Bacillus是目前研究较为广泛的一类生防菌资源,具有抑菌谱广、环境适应性强的特点,并且无毒、无污染,不易使病原菌产生抗药性,目前已有多种芽胞杆菌应用于微生物杀菌剂研发中(http://www.chinapesticide.org.cn/hysj/index.jhtml)。芽胞杆菌能够产生多种抗菌物质,包括脂肽类、肽类、细菌素等。其中脂肽类物质一般包括丰产素家族fengycin、伊枯草菌素家族iturin和表面活性素家族surfactin三大类。Fengycin具有较强的抗丝状真菌作用及低溶血活性[1],在农业及医药等领域有重要的应用前景[2,3]。Iturin对病原真菌的抑制作用较强,对细菌及病毒的抑制作用较弱[4]。Surfactin具有优异的表面活性及溶血功能,能够通过影响生物膜的形成从而影响细菌的定殖能力和生防效果[5];Surfactin与iturin A协同作用能够显著增强iturin A的抑真菌活性[6],与fengycin共同作用则能够诱导植物产生诱导系统抗性(ISR)[7]。
脂肽类物质是通过非核糖体途径合成的,除受调控因子调控外[8],还受多种环境条件如培养基组分、发酵条件(温度、时间、溶氧)等的影响,其中培养基组分是影响抗菌脂肽合成的主要因素[9]。为了提高芽胞杆菌抗菌脂肽的产量,国内外学者进行了大量的研究工作。Yi等[10]利用响应曲面法对枯草芽胞杆菌N7产抗菌脂肽的发酵培养基各成分用量进行了优化,抗菌脂肽的产量达到706.57 mg/L;Liu等[11]通过响应曲面法优化了解淀粉芽胞杆菌MB199产surfactin的发酵培养基组分比例,surfactin的产量较优化前提高了1.52倍。这些研究大都集中在发酵培养基及发酵条件的优化上,而针对影响抗菌脂肽合成的碳、氮源的筛选,尤其是关于糖、氨基酸影响抗菌脂肽合成的研究报道相对较少。
枯草芽胞杆菌B.subtilisBAB-1是本实验室分离获得的一株生防细菌,能够有效地防治番茄灰霉病、瓜类白粉病等气传病害[12,13],现已成功开发成环保、高效的微生物杀菌剂“80亿CFU/mL枯草芽胞杆菌悬浮剂”(登记证号:PD20150190)。前期研究发现,枯草芽胞杆菌BAB-1能够产生脂肽类物质fengycin与surfactin,fengycin对番茄灰霉菌具有较强的抑菌作用[14];而surfactin能够加速细菌的表面运动,促进菌群扩展[15]。在此基础上,本研究采用快速蛋白液相色谱(FPLC)分析不同碳、氮源对菌株BAB-1抗菌脂肽产量的影响,并进一步采用溶血圈试验及排油圈法测定粗脂肽的性质,为指导抗菌脂肽的合成与调控提供理论依据,从而进一步为以获取脂肽类物质为目的的发酵生产奠定基础。
1 材料与方法
1.1 材料
枯草芽胞杆菌BAB-1(GenBank登录号:CP004405.1)由河北省农林科学院植物保护研究所植物病害生物防治实验室分离并保存。
LB固体培养基:胰蛋白胨10 g、酵母粉5 g、NaCl 5 g、琼脂15 g、H2O 1000 mL;LB液体培养基:胰蛋白胨10 g、酵母粉5 g、NaCl 5 g、H2O 1000 mL;Landy培养基:葡萄糖20 g、L-谷氨酸钠5 g、MgSO40.5 g、KCl 0.5 g、KH2PO41 g、FeSO40.15 mg、MnSO45 mg、CuSO40.16 mg、H2O 1000 mL;改良水琼脂培养基:琼脂1.5 g、H2O 1000 mL;血琼脂培养基:在水琼脂培养基中添加5%的无菌脱纤维绵羊全血。
1.2 脂肽类物质的提取及检测
将活化的菌株BAB-1单菌落接种于LB液体培养基中,30 ℃、180 r/min培养12 h得到菌株BAB-1种子液,将种子液以2%的接种量接入到发酵培养基中,30 ℃、180 r/min培养48 h。培养结束后,首先测定发酵液的OD600值,再将发酵液于4 ℃、8000 r/min离心20 min收集上清,在上清中缓慢加入6 mol/L的HCl调节pH值至2.0,4 ℃过夜沉淀。8000 r/min离心20 min收集沉淀,待沉淀干燥后加入原培养液体积的1/10甲醇抽提2次,每次2 h,合并抽提物并用直径0.22 μm的细菌过滤器过滤即得到脂肽粗提物。
采用快速蛋白液相色谱(FPLC)梯度洗脱分离脂肽化合物。FPLC分析系统为AKTA Purifier 10液相分析系统,色谱柱为SOURCE 5RPC ST 4.6/150柱。流动相A为含0.065%三氟乙酸和2%乙腈的水溶液,流动相B为含0.05%三氟乙酸的80%乙腈溶液。检测波长为215 nm,流速为1 mL/min。梯度洗脱过程为49.86 min内流动相A由100%到0[14]。
1.3 不同碳氮源对菌株BAB-1抗菌脂肽产量的影响
1.3.1 不同碳源对菌株BAB-1抗菌脂肽产量的影响 以Landy培养基为基础培养基,分别以等量的葡萄糖、D-木糖、D-核糖、L-阿拉伯糖与熊果苷作为碳源,L-谷氨酸钠作为氮源,配制不同碳源的培养基。按1.2的方法培养、提取脂肽,并进行FPLC分析。根据不同培养基脂肽的峰面积/发酵液OD600的比值比较不同碳源对菌株BAB-1脂肽产量的影响,每处理3次重复。
1.3.2 不同氮源对菌株BAB-1抗菌脂肽产量的影响 以Landy培养基为基础培养基,分别以等量的L-谷氨酸钠、L-半胱氨酸、L-谷氨酰胺、L-脯氨酸与L-天冬酰胺作为氮源,葡萄糖作为碳源,配制不同氮源的培养基,按1.2的方法培养、提取脂肽,并进行FPLC分析。根据不同培养基脂肽的峰面积/发酵液OD600的比值比较不同氮源对菌株BAB-1脂肽产量的影响,每处理3次重复。
1.4 不同碳氮源对菌株BAB-1粗脂肽溶血活性的影响
将配制好的血琼脂培养基倒入无菌培养皿中(φ=9 cm),待其凝固后在血平板上放置无菌牛津杯,每牛津杯注入200 μL从不同碳、氮源培养基中提取的菌株BAB-1的脂肽粗提物,30 ℃培养3 d后观察并测量溶血圈直径大小。以甲醇作为对照,每处理3次重复。
1.5 不同碳氮源对菌株BAB-1粗脂肽排油能力的影响
在无菌培养皿(φ=9 cm)中加入15 mL无菌水,滴加10 μL从不同碳氮源培养基中提取的菌株BAB-1脂肽粗提物,待整个培养皿表面形成一层薄膜后,在各培养皿中央滴加10 μL矿物油,观察并测量矿物油的扩散直径,矿物油的扩散直径越小说明脂肽粗提物的排油能力越强。以甲醇作为对照,每处理3次重复。
1.6 RT-PCR分析不同碳源条件下菌株BAB-1中srfAA基因差异表达
将菌株BAB-1种子液以2%的接种量转接到100 mL不同碳源的Landy培养基中,30 ℃、180 r/min振荡培养14 h,分别取2 mL不同碳源的培养液,利用细菌总RNA提取试剂盒(北京全式金生物技术有限公司)提取RNA后立即进行反转录反应。反转录结束后将合成的cDNA作为模板进行RT-qPCR分析,测定srfAA基因的表达情况。srfAA基因引物序列为:SrfAA-F(5′-GTCCGCAACAACAGATGACA-3′),SrfAA-R (5′-AAGAACAGCATTGAGGCGAC-3′)。扩增反应体系为 20 μL,其中 2×Trans Start Tip Green qPCR Super mix 10 μL,引物(10 μmol/L)各 0.4 μL、50×Passive Reference Dye 0.4 μL、cDNA 模板 2 μL、ddH2O 6.8 μL。PCR反应条件:94 ℃ 2 min;98 ℃ 10 s、60 ℃ 10 s、68 ℃ 30 s、40 个循环;设gyrB基因为内参基因,gyrB基因引物序列为 GyrB-F (5′-GAAGCACGGACAATCACC-3′)和 GyrB-R(5′-TCCAAAGCACTCTTACGG-3′)。利用公式2-ΔΔCT计算目标基因在不同处理之间的相对变化倍数。
1.7 数据统计与分析
本试验所有数据经Excel软件进行整理,采用IBM SPSS Statistics19.0软件进行ANOVA方差分析。
2 结果与分析
2.1 不同碳氮源对菌株BAB-1脂肽产量的影响
2.1.1 不同碳源对菌株BAB-1脂肽产量的影响 以熊果苷作为碳源时,菌株BAB-1的surfactin产量显著高于葡萄糖、D-木糖、D-核糖及L-阿拉伯糖作为碳源产量surfactin,分别是这4种碳源的2.95、7.01、4.26和5.60倍;而fengycin产量与葡萄糖作为碳源时无显著差异,但显著高于D-木糖、D-核糖及L-阿拉伯糖作为碳源的产量(图1)。
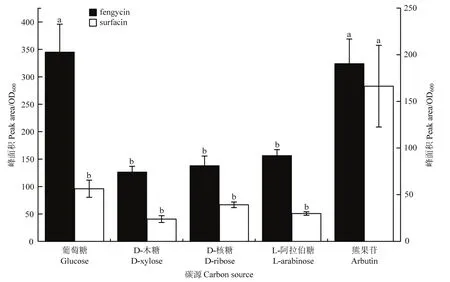
图1 不同碳源对枯草芽胞杆菌BAB-1 脂肽产量的影响Fig.1 Influence of different carbon sources on lipopeptide production from B.subtilis BAB-1
2.1.2 不同氮源对菌株BAB-1脂肽产量的影响 以L-天冬酰胺为氮源时菌株BAB-1 surfactin及fengycin的产量与 L-谷氨酸钠为氮源时的产量无显著差异,但显著高于 L-半胱氨酸、L-脯氨酸及 L-谷氨酰胺为氮源的产量,L-半胱氨酸及L-谷氨酰胺极显著降低菌株BAB-1 fengycin的产量(图2)。
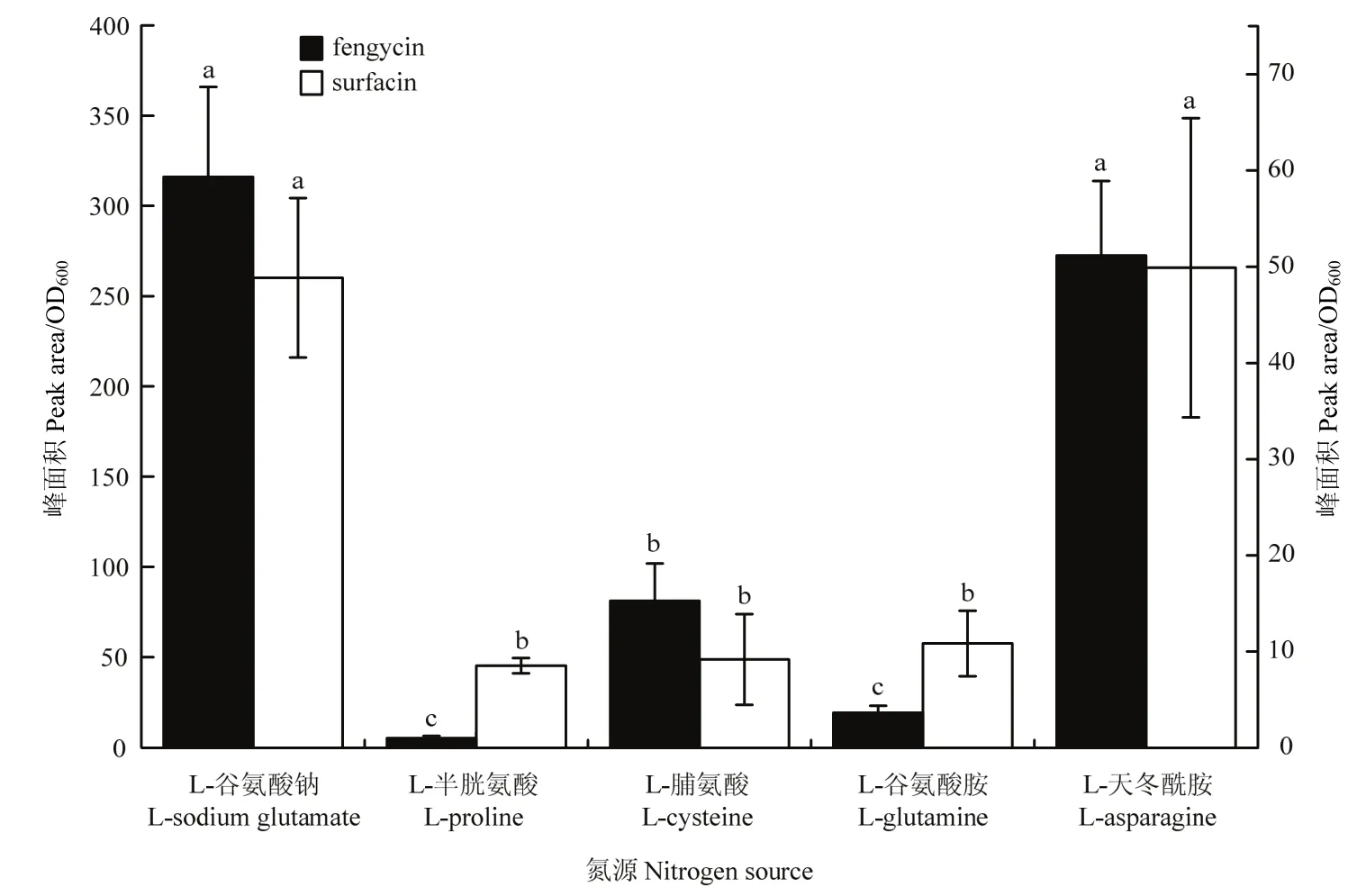
图2 不同氮源对枯草芽胞杆菌BAB-1脂肽产量的影响Fig.2 Influence of different nitrogen substrates on lipopeptide production from B.subtilis BAB-1
2.2 不同碳源对BAB-1菌株粗脂肽溶血活性及排油能力的影响
以熊果苷作为碳源时,菌株BAB-1粗脂肽的溶血活性最强,其溶血圈直径达到3.53 cm,显著高于葡萄糖、D-木糖、D-核糖及 L-阿拉伯糖作碳源时该菌株粗脂肽的溶血活性;以葡萄糖与 L-阿拉伯糖为碳源时,菌株BAB-1粗脂肽的溶血活性无显著差异,溶血圈直径分别为1.63和1.60 cm;以D-核糖为碳源时,该菌株粗脂肽的溶血活性最低,溶血圈直径仅为0.63 cm(图3)。
以熊果苷作为碳源时,菌株BAB-1粗脂肽的排油能力最强,油滴扩散直径仅为0.27 cm,显著优于其他4种碳源的排油能力;以葡萄糖与D-木糖为碳源时,该菌株粗脂肽的排油能力无显著差异,扩散直径分别为0.57与0.53 cm;以D-核糖为碳源时,菌株BAB-1所产脂肽排油能力最低,滴入矿物油后立即散开,扩散直径达到5.30 cm(图3)。
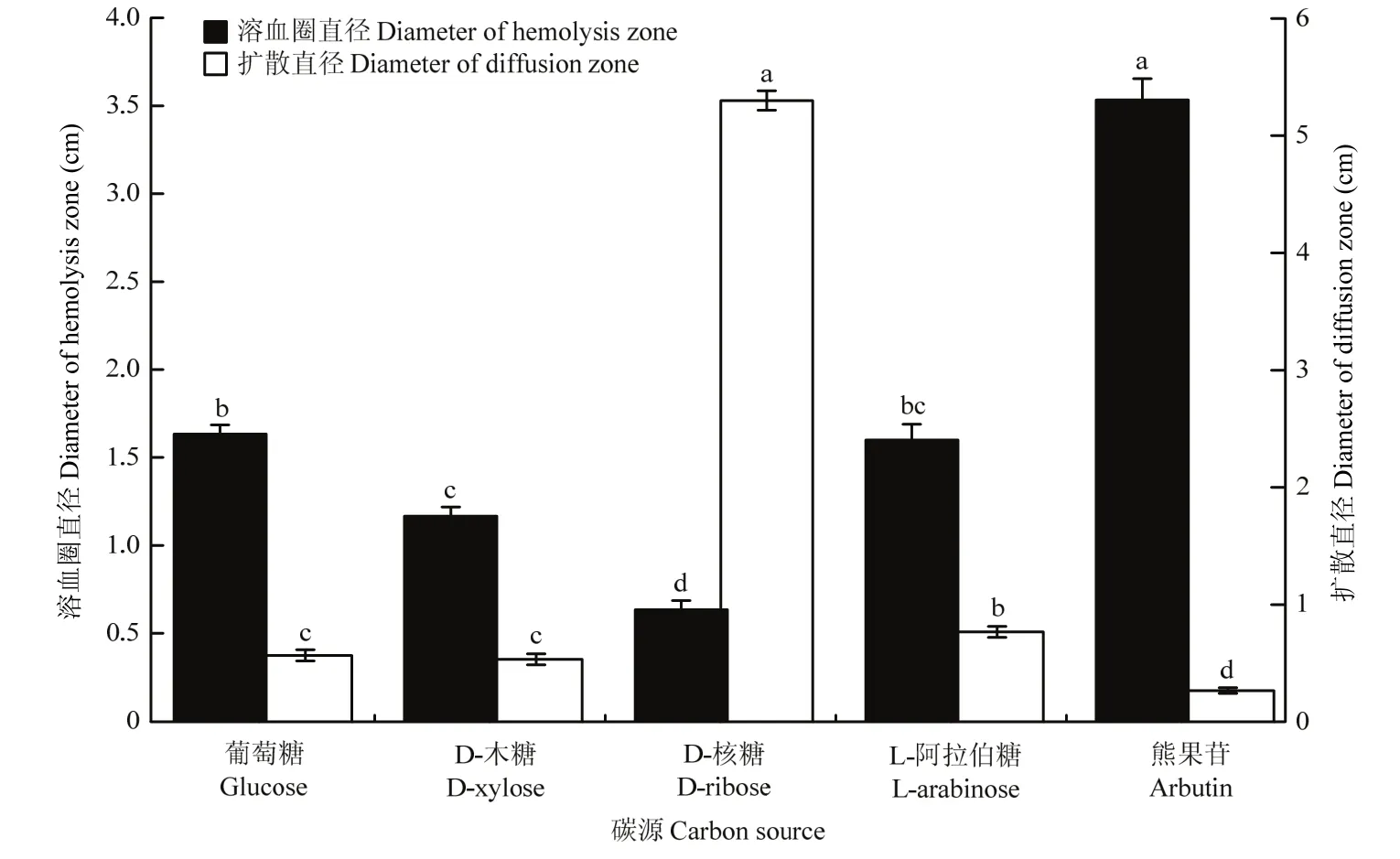
图3 不同碳源对枯草芽胞杆菌BAB-1粗脂肽溶血活性及排油能力的影响Fig.3 Influence of different carbon sources on hemolysis and oil discharge capacity of crude lipopeptide from B.subtilis BAB-1
2.3 不同氮源对菌株BAB-1粗脂肽溶血活性及排油能力的影响
以L-天冬酰胺为氮源时,菌株BAB-1粗脂肽的溶血活性与L-谷氨酸钠作为氮源时无显著差异,溶血圈直径分别为1.43与1.30 cm,且显著高于以L-脯氨酸、L-半胱氨酸及L-谷氨酰胺作氮源时溶血活性(溶血圈直径均为0.60 cm)(图4)。
以L-天冬酰胺为氮源时,菌株BAB-1粗脂肽具有较好的排油能力,油滴扩散直径为0.43 cm,与L-谷氨酸钠的0.47 cm无显著差异,但显著优于以L-脯氨酸、L-半胱氨酸及L-谷氨酰胺为氮源时该菌株粗脂肽的排油能力(油滴扩散直径分别为5.38、5.43和5.23 cm)(图4)。
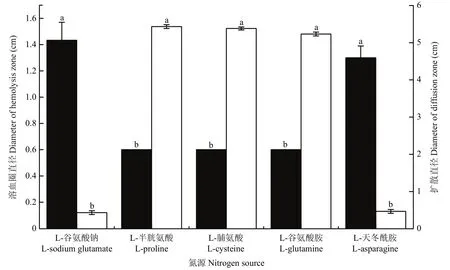
图4 不同氮源对枯草芽胞杆菌BAB-1粗脂肽溶血活性及排油能力的影响Fig.4 Influence of different nitrogen sources on hemolysis and oil discharge capacity of crude lipopeptide from B.subtilis BAB-1
2.4 不同碳源条件下srfAA基因差异表达分析
与葡萄糖为碳源相比,菌株BAB-1中srfAA基因的表达量在以熊果苷为碳源时提高了2.9倍,与FPLC分析的结果一致;但以D-木糖、D-核糖与L-阿拉伯糖为碳源时,srfAA基因的表达量分别降低了1.4、2.6和1.1倍(表1)。
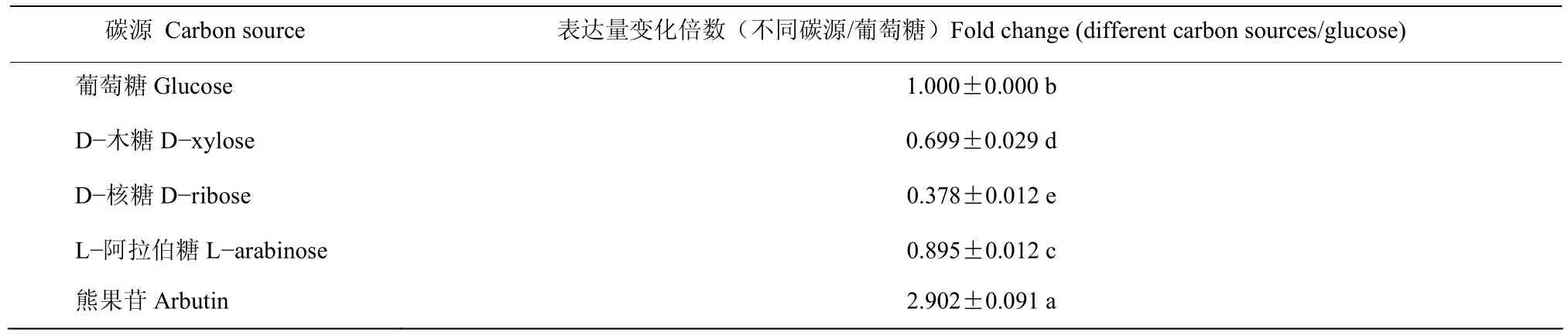
表1 不同碳源条件下srfAA基因的相对表达情况Table 1 Comparsion of expression of srfAA gene in B.subtilis BAB-1 in different carbon sources using RT-qPCR
3 讨论
枯草芽胞杆菌通过与病原菌进行营养和空间竞争、分泌多种次生代谢产物抑制或杀死病原菌及诱导植物产生抗病性等途径达到其防治植物病害的目的。脂肽类抑菌物质如surfactin、iturin和fengycin等是芽胞杆菌抑制病原真菌生长的主要抑菌物质[16],其中 fengycin与 iturin能够抑制多种病原真菌的生长[17,18],surfactin具有较强的表面活性,能够促进芽胞杆菌在植物体表的定殖能力进而影响其防治效果[5],同时fengycin与 surfactin还可以作为诱导因子引起植物的诱导抗病性[7]。本实验室前期研究表明,fengycin与surfactin是枯草芽胞杆菌BAB-1发挥防病作用的主要物质[14,15],因此,提高芽胞杆菌脂肽类抗菌物质的产量对提高其防效具有重要意义。
通过明确不同营养物质对生防芽胞杆菌产抗菌脂肽的影响,对于其定向发酵生产具有重要的指导意义。碳源与氮源是脂肽类抗菌物质产生所必需的营养成分,对脂肽产量的影响较大[19-21]。葡萄糖是芽胞杆菌的优选碳源[22,23],国内外学者围绕葡萄糖对芽胞杆菌脂肽类物质产生的影响开展了一定研究。Sandrin等[24]发现葡萄糖是产生surfactin和iturin的良好碳源;Nihorimbere等[25]报道葡萄糖有利于fengycin和iturin的积累;Roongawang等[26]研究表明葡萄糖是surfactin、bacillomycin L和plipastatin的有效碳源。除葡萄糖之外,本研究选择了 D-木糖、D-核糖、L-阿拉伯糖及熊果苷作为碳源,这 4种糖是通过表型芯片技术(phenotype microarray,PM)筛选出的能够明显促进菌株BAB-1生长的物质[27]。通过FPLC分析发现,熊果苷能够显著提高surfactin的产量,是葡萄糖作碳源时surfactin产量的2.95倍,这就说明不同菌株合成特定抗菌脂肽所需的碳源种类是不同的,同时也证明了PM技术能够应用于芽胞杆菌脂肽类物质发酵工艺营养物质的筛选中。Nakano等[28]报道,葡萄糖能够连续增加surfactin合成酶基因srfA的表达水平从而提高surfactin的产量,因此本研究检测了熊果苷对srfAA基因表达水平的影响发现,熊果苷可以显著提高该基因的表达水平,相比葡萄糖提高了2.9倍。
脂肽类化合物一般是由1个β-羟基脂肪酸与7~10个氨基酸以酰胺键形式连接而形成的环肽[29]。关于抗菌脂肽的组成成分氨基酸是否影响其合成的研究报道较少。Peypoux等[30]报道,L-缬氨酸与 L-异亮氨酸能够促进surfactin合成;Doekel等[31]报道,L-谷氨酸和L-天冬氨酸能够显著提高抗菌脂肽lichenysin-A的产量;孙力军等[32]研究表明,L-谷氨酸和L-天冬酰胺是抗菌脂肽fengycin与surfactin产生的良好氮源,能够促进抗菌脂肽的产生。本研究选择了能够明显促进菌株BAB-1生长的4种物质L-脯氨酸、L-半胱氨酸、L-谷氨酰胺及 L-天冬酰胺[27]以及 L-谷氨酸钠作为氮源结果发现,L-天冬酰胺及 L-谷氨酸钠利于fengycin与surfactin的产生,这与孙力军等[32]的研究结果一致。但在fengycin与surfactin的肽链结构中,并没有天冬酰胺残基,天冬酰胺对这2种物质的促产作用,可能是由于天冬酰胺在菌体内的代谢途径影响了2种物质的合成,具体原因还需进一步研究。
据报道,surfactin是枯草芽胞杆菌在植物根际产生的主要脂肽类物质[25,33],能够促进生防菌在根部的定殖进而提高防病效果。Surfactin可以通过溶血圈法结合排油圈法进行定性检测[34],并利用FPLC进行定量检测[35]。本研究通过FPLC分析发现,熊果苷能够显著提高菌株BAB-1的surfactin产量,进一步采用溶血圈法与排油圈法验证了熊果苷作为碳源时该菌株粗脂肽的溶血活性与排油能力最强,与FPLC结果一致。Li等[36]报道,枯草芽胞杆菌突变株Bs-H74 surfactin产生能力比野生型菌株提高3倍,同时对水稻纹枯病的防效也提高了14.6%。在本研究中熊果苷促进surfactin产量增加,是否能够提高菌株BAB-1在植物体表的定殖能力与防病效果还需要进一步试验验证。
本研究发现,虽然熊果苷作为碳源、L-谷氨酸钠作为氮源能够显著提高菌株BAB-1的surfactin产量,但fengycin产量与葡萄糖作为碳源时没有显著差异;综合成本因素考虑,葡萄糖与L-谷氨酸钠是BAB-1菌株产脂肽最适宜的碳、氮源。但可以寻找熊果苷廉价的替代品,有效降低发酵成本,为进一步中试放大及产业化生产提供科学依据。
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09
防爆电机(2022年2期)2022-04-26
微生物学杂志(2021年2期)2021-07-01
湖南电力(2021年1期)2021-04-13
微生物学杂志(2020年2期)2020-12-31
宁夏电力(2020年1期)2020-05-29
山东化工(2020年4期)2020-03-30
分析科学学报(2019年2期)2019-04-25
测控技术(2018年3期)2018-11-25
物理化学学报(2015年7期)2015-12-30
