
高寒草地主要物种对放牧方式的响应
2021-04-20 07:32:14杨晓霞董全民张春平刘文亭张小芳孙彩彩杨增增张艳芬
草业科学 2021年3期
冯 斌,杨晓霞,董全民,张春平,刘文亭,俞 旸,张小芳,孙彩彩,时 光,杨增增,张艳芬
(青海大学畜牧兽医科学院 / 青海省畜牧兽医科学院 / 青海大学省部共建三江源生态与高原农牧业国家重点实验室 /青海大学青海省高寒草地适应性管理重点实验室,青海 西宁 810016)
青藏高原草地类型丰富多样,幅员辽阔。当前,放牧仍然是青藏高原高寒草地最基本的利用方式之一。草畜互作是高寒草地草畜平衡研究和放牧生态学的重要内容[1-2]。放牧会影响不同物种在群落中的竞争力,在草地生态系统当中功能群和物种的多样性对于维持草地生态系统的稳定具有重要的意义。相关研究表明,草地生态系统功能群多样性相较于物种多样性,其对草地生态系统的生产力和稳定性影响可能更为显著[3-5]。另有研究表明,适口性好的牧草受到家畜采食压力后在群落中的竞争优势减弱,特别是在重度放牧下表现尤为突出[6]。这种放牧干扰对群落的影响体现在群落结构和稳定性上,例如当草地受重度放牧干扰后,群落组成易发生改变,造成群落组成和结构的单一化,而当环境再度剧烈变化就会导致草地退化,严重表现为荒漠化和沙漠化[7]。当前关于放牧与群落关系的研究,主要集中在群落中某一优势物种在放牧干扰下个体功能性状的响应机制[8-9],据此区分植物功能性状的敏感性指标和惰性指标[10]。如羊草(Leymus chinensis)在放牧或刈割条件下株高会降低[11-12],在放牧条件下羊草的单株重也会降低[8];在不同的经营方式下短花针茅(Stipa breviflora)的自然叶高、叶长、叶鲜质量、叶干质量等叶性状受放牧的影响最大[13];关于放牧对群落生产力影响的研究表明,植物群落在放牧干扰下存在普遍的补偿性生长机制[14]。植物功能性状对环境干扰的响应,不仅仅包括放牧干扰,也包括气候变化、区域差异、土壤肥力差异等多方面。
在放牧系统中植物的个体特征、生理生化功能、生长和繁殖均受家畜采食、践踏和粪尿的影响,反之家畜的采食和营养供给也受植物类别、高度和适口性的影响[15-16],基于此,在漫长的生物进化史中动植物的相互影响促进了动物与植物的协同进化[17]。此外,依据采食优化理论,大型家畜食性选择很大程度上取决于采食过程中能量代谢的权衡[15,18]。以牦牛和藏羊为例,牦牛舌头具倒刺,在采食植物时采用“舌扫荡齿切”式,藏羊口窄且拱形门齿高度弯曲[19];而营养方面,藏羊喜食高营养低次生代谢物植物,而牦牛则可忍受营养相对较低但生物量较高的一类植物[20-21],这都是导致不同类型的家畜在采食过程存在偏食性的基础。当前关于放牧对草地生态系统影响的研究表明,不同类型家畜采食过程中对植物的选择性会影响草地的群落结构和植物的个体特征[22-23]。基于此,本研究以禁牧为对照,结合禾本科、莎草科、豆科和杂类草等各功能群主要物种的频度探究不同放牧方式对各功能群主要物种的个体特征影响,并以此为依据讨论不同放牧方式对青海湖流域高寒草地群落结构和植物个体特征的影响,揭示放牧与禁牧、不同畜种以及不同畜种混牧比例配置高寒草地各功能群主要物种的响应机制和各功能群物种个体特征的变化特点,从而为高寒草地的放牧利用和管理提供科学依据和理论支撑。
1 材料与方法
1.1 研究区概况
研究样地位于青海省海北州海晏县西海镇(36°92′ N,100°93′ E,海拔3 000~3 100 m),隶属于青海湖流域,气候为高原大陆性气候,无明显的四季之分,只有冷暖季之分,冷季漫长而寒冷干燥,暖季短暂而凉爽湿润,年内无绝对无霜期,年日照时数为2 580~2 750 h,年均降水量为400 mm,年均温1.5 ℃,草地类型为高寒草原化草甸。样地植物主要物种有矮生嵩草(Kobresia humilis)、干生薹草(Carex aridula)、紫花针茅(Stipa purpurea)、早熟禾(Poa annua)、 洽 草(Koeleria macrantha)、 赖 草(Leymus secalinus)、垂穗披碱草(Elymus nutans)、斜茎黄耆(Astragalus laxmannii)、 星 毛 委 陵 菜 (Potentilla acaulis)和异叶青兰(Dracocephalum heterophyllum)等[24](表1),土壤类型为高山草甸土。
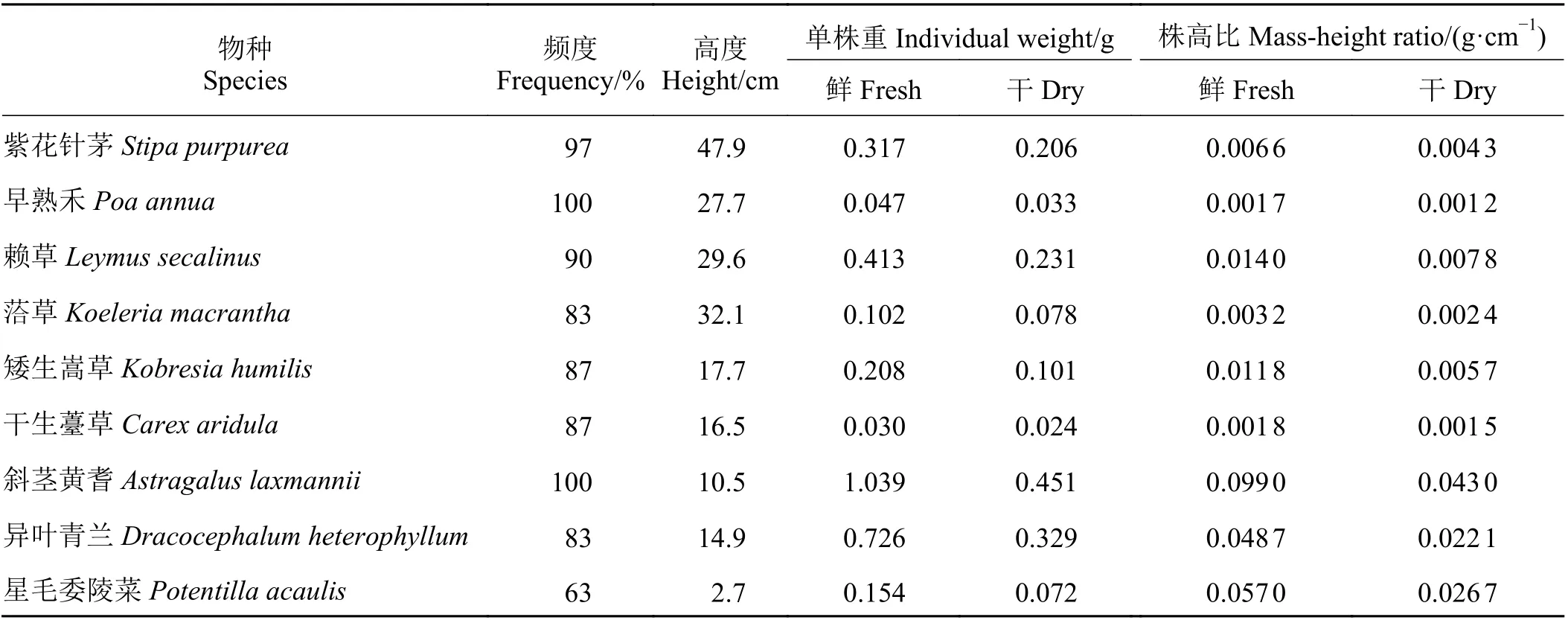
表1 各功能群主要物种在对照样地的基本概况Table 1 Basic characteristics of major species of each functional group in the control plot
1.2 研究方法
1.2.1 试验设计
试验样地为天然放牧地且基况较均匀,放牧试验开展于2014 年,采用随机区组设计,设禁牧对照(CK)、牦牛单牧(YG)、藏羊单牧(SG)、牦牛 : 藏羊 =1 : 6 混牧(MG 1 : 6)、牦牛 : 藏羊=1 : 4 混牧(MG 1 : 4)、牦牛 : 藏羊 = 1 : 2 混牧(MG 1 : 2) 6 个处理(表2),每个处理设有3 个重复,共有18 个试验小区。所有放牧处理的放牧强度均为中等放牧强度(11.6 AU·hm−2),即牧草利用率为50%~55%。放牧在植物生长季6 月 − 10 月进行,每月放牧10 d 左右,当牧草利用率达到50%时,将家畜转场至邻近草地放牧。放牧第6 年(即2019 年)测定放牧样地禾本科、莎草科、豆科和杂类草等各功能群主要物种的频度和个体特征。
为保证试验的一致性,选择同龄的公牦牛和公藏羊,牦牛年龄为1.5 岁,体重为(100 ± 5) kg,藏羊年龄为1 岁,体重为(30 ± 2) kg。每头牦牛为3 个羊单位,每只藏羊为1 个羊单位,每个处理所需的草地面积根据羊单位进行计算,以保证所有处理的放牧强度一致。放牧前对家畜进行了投药驱虫,以确保家畜在放牧期间的正常采食和代谢活动。

表2 放牧试验设计Table 2 Grazing experiment design
1.2.2 取样和测定
通过前期调查,本研究选择各样地共有且易于受放牧影响的9 个物种,即禾本科牧草紫花针茅、早熟禾、洽草和赖草,莎草科牧草矮生嵩草和干生薹草,豆科植物斜茎黄耆,以及杂类草异叶青兰和星毛委陵菜作为研究对象,将其频度、株高、单株重和株高比作为评估指标。频度测定以50 cm ×50 cm的样方框在各试验小区随机扔15 次并记录每次样方框中出现的物种。在每个试验小区选择各目标物种30 株测定其自然高度,同时将测量株高之后的植株以基部为基准,齐地面剪下后带回实验室后,在85 ℃下烘干至恒重后称重。
通过计算物种平均单株重和平均株高的比值,探讨物种单位高度的质量变化,本研究中将其定义为株高比,即:
株高比(g·cm−1) = 平均单株重/平均株高。
1.3 数据分析与处理
采用R 4.0.1 (R Development Core Team,2019)进行数据分析处理,用平均值±标准误表示测定结果。各处理物种频度、高度、单株重、株高比均采用单因素方差分析(One-way ANOVA),运用Tukey-HSD对测定各数据进行处理间的多重比较,并采用Corrplot包对各功能群主要物种各指标进行相关性分析。使用Sigma Plot 12.5 进行绘图。
2 结果与分析
2.1 放牧方式对青海湖流域高寒草地各功能群主要物种频度的影响
禾本科牧草紫花针茅、早熟禾、赖草和洽草,莎草科牧草矮生嵩草和干生薹草,豆科牧草斜茎黄耆,杂类草异叶青兰和星毛委陵菜在群落中的频度,YG 和SG 处理之间均无显著差异(P > 0.05),也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2 MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)改变存在明显的不同。仅MG 1 : 2 与CK 相比,紫花针茅、早熟禾和洽草在群落中其频度分别显著降低了21.12%、11.00%和48.33%(P < 0.05);与CK 相比,斜茎黄耆频度在YG 和MG 1 : 6 中分别显著降低了28.89%和31.11% (P < 0.05);与CK 相比,MG 1 : 4 处理中异叶青兰频度降低了39.76% (P < 0.05);与CK 相比,星毛委陵菜的频度在各放牧处理下均显著增加(P < 0.05),其频度在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 中分别显著增加了32.14%、35.22%、30.48%、33.72%和28.97% (P < 0.05)。另对比各放牧处理发现,紫花针茅频度MG 1 : 2 与SG、MG1 : 6 相比显著下降 (P < 0.05),早熟禾和赖草的频度MG 1 : 2 与其余各放牧处理相比显著下降(P < 0.05),其余各物种的频度在不同的放牧处理之间无显著差异(图1)。
2.2 放牧方式对青海湖流域高寒草地各功能群主要物种株高的影响
禾本科牧草紫花针茅、早熟禾、赖草和洽草,莎草科牧草矮生嵩草和干生薹草,豆科牧草斜茎黄耆,杂类草星毛委陵菜在群落中的株高,YG 和SG 均无显著差异(P > 0.05),也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6与MG 1 : 2 相比)的改变存在明显的不同,仅禾本科牧草紫花针茅和杂类草星毛委陵菜株高在YG 与MG 1 : 6、MG 1 : 4 之 间 存 在 显 著 差 异(P <0.05),杂类草异叶青兰株高在YG 和SG 之间存在显著差异(P < 0.05)。与CK 相比,紫花针茅株高在SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 显著降低了22.09%、37.40%、38.69%和31.63% (P < 0.05);与CK 相 比,YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 中早熟禾株高分别降低了29.62%、32.12%、31.24%、34.66%和38.47%,赖草株高分别降低了47.36%、37.82%、38.63%、44.79%和41.25%,洽草株高分别降低了19.84%、21.87%、32.48%、26.42%和34.13%。与CK 相比,YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2显著降低了矮生嵩草和干生薹草的株高(P < 0.05),矮生嵩草在各放牧处理下株高分别降低了18.70%、12.57%、18.91%、22.11%和21.58%;干生薹草在各放牧处理下株高分别降低了27.74%、27.14%、30.61%、31.73%和34.98%。与CK 相比,斜茎黄耆株高在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 分别降低了46.03%、40.16%、55.12%、61.20%和56.17%。 与CK 相比,异叶青兰株高在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2中分别降低了32.62%、49.34%、53.12%、 58.20%和54.93%;与CK 相比,SG、MG 1 : 6、MG 1 : 4 和 MG 1 : 2下星毛委陵菜的株高分别下降了19.69%、 30.44%、26.44%和22.92% (图2)。
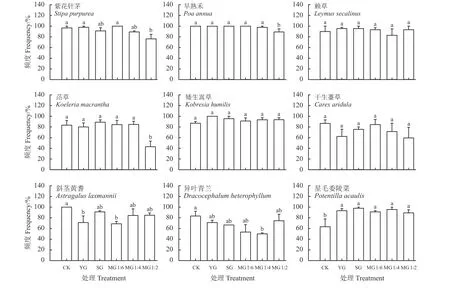
图1 放牧方式对环青海湖高寒草地主要物种频度的影响Figure 1 Effect of grazing mode on the frequency of major species in an alpine grassland of Qinghai lake basin
2.3 放牧方式对青海湖流域高寒草地各功能群主要物种单株重的影响

图2 放牧方式对环青海湖高寒草地主要物种株高的影响Figure 2 Effect of grazing mode on the height of major species in an alpine grassland in the Qinghai Lake basin
禾本科牧草赖草和洽草在群落中的单株重,YG 和SG 处理间均无显著差异(P > 0.05),也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在明显不同,仅紫花针茅和早熟禾的单株重YG 和SG 之间存在显著差异(P < 0.05);与CK 相比,YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理紫花针茅的单株重显著降低(P < 0.05),分别降低了19.89%、36.57%、45.68%、59.32%和51.17%;与CK 相比,YG、MG 1 : 6、MG 1 : 4和MG 1 : 2 处理早熟禾的单株重显著降低(P < 0.05),分别降低了10.61%、25.35%、32.12%和33.64%;与CK相比,YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理赖草单株重显著下降(P < 0.05),分别降低了45.13%、46.03%、44.02%、47.10%和52.86%;与CK 相比,MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理洽草单株重显著下降(P <0.05),分别降低了47.14%、39.44%和45.77%。莎草科牧草干生薹草在群落中的单株重,YG 和SG 处理间无显著差异,也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2相比)的改变存在明显不同,但莎草科牧草矮生嵩草单株重在YG 和SG 之间差异显著 (P < 0.05),且随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在显著差异(P <0.05),其中MG 1 : 4 处理矮生嵩草的单株重显著高于MG 1 : 6 和MG 1 : 2 (P < 0.05)。此外与CK 相比,在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理矮生嵩草的单株重分别增加了43.20%、37.96%、31.52%、43.98%和29.48%;与CK 相比,SG 和MG 1 : 2 处理干生薹草单株重增加了12.41%和13.56%。豆科牧草斜茎黄耆在群落中的单株重,YG 和SG 处理间无显著差异(P > 0.05),也并未表现出随家畜数量的相对 增 加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在明显不同,仅与CK 相比,MG 1 : 6 处理斜茎黄耆单株重显著下降了31.44%(P < 0.05)。杂类草物种异叶青兰和星毛委陵菜单株重在YG 和SG 处理之间差异显著(P < 0.05),但并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在明显不同,且与CK 相比,异叶青兰单株重在SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理分别显著下降了40.21%、 33.98%、 46.63%和38.43% (P < 0.05); 与CK 相比,YG 处理星毛委陵菜的单株重显著增加了33.16% (图3)。
2.4 放牧方式对青海湖流域高寒草地各功能群主要物种株高比的影响

图3 放牧方式对环青海湖高寒草地主要物种单株重的影响Figure 3 Effect of grazing mode on the individual weight of major species in an alpine grassland in Qinghai Lake basin
尽管禾本科牧草早熟禾、赖草和洽草在群落中的株高比在YG 和SG 处理间均无显著差异(P >0.05),也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在明显不同,但禾本科牧草紫花针茅株高比在YG 和SG 处理之间差异显著,MG 1 : 4 和MG 1 : 2与CK 之间差异显著(P < 0.05);早熟禾株高比各放牧处理高于CK,且YG、SG 和MG 1 : 2 与CK 相比其株高比差异显著(P < 0.05);莎草科牧草干生薹草在群落中的株高比,YG 和SG 处理间无显著差异,也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在显著差异。与CK 相比,干生薹草株高比在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2 处理分别显著增加了40.98%、41.62%、40.08%、34.49%和43.91% (P <0.05) ,而莎草科牧草矮生嵩草株高比在YG 与SG 处理之间差异显著(P < 0.05),与CK 相比,矮生嵩草株高比在YG、SG、MG 1 : 6、MG 1 : 4 和MG 1 : 2处理分别增加了52.85%、42.54%、40.95%、54.15%和41.38%。豆科牧草斜茎黄耆和杂类草异叶青兰和星毛委陵菜在群落中的株高比,YG 和SG 处理间无显著差异,也并未表现出随家畜数量的相对增加(SG 与MG 1 : 2、MG 1 : 4 和MG 1 : 6 相比)或牦牛和藏羊数量配比(MG 1 : 4、MG 1 : 6 与MG 1 : 2 相比)的改变存在明显不同,仅杂类草星毛委陵菜的株高比在MG 1 : 6 与MG 1 : 2 处理之间差异显著(P < 0.05)。与CK 相比,斜茎黄耆在MG 1 : 2 处理增加了60%。与CK 相比,异叶青兰在MG 1 : 6、MG 1 : 4和MG 1 : 2 处理分别增加了44.47%、46.83%和48.38%;与CK 相比,星毛委陵菜株高比在YG、SG、MG 1 : 6 和MG 1 : 4 处理分别增加了38.39%、31.09%、43.31%和29.22% (图4)。
2.5 各功能群主要物种各指标之间的相关性分析
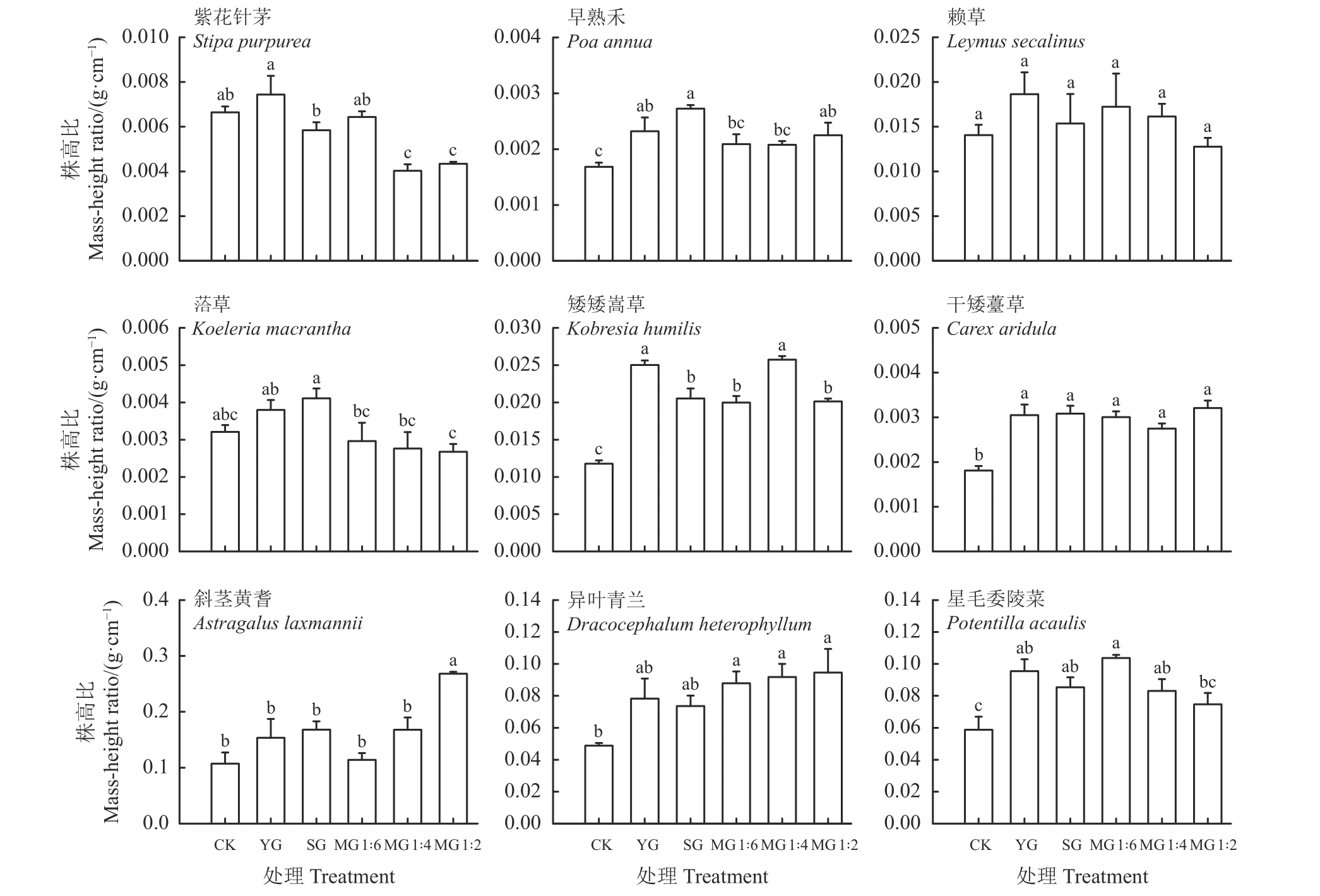
图4 放牧方式对环青海湖高寒草地主要物种株高比的影响Figure 4 Effect of grazing mode on the mass-height ratio of major species in an alpine grassland in Qinghai Lake basin
对高寒草地禾本科、莎草科、豆科和杂类草等各功能群主要物种个体特征有显著作用的4 个性状指标进行比较(表3),发现禾本科主要物种的单株重和株高、株高比之间呈极显著正相关关系 (P <0.01),株高和株高比之间呈显著负相关关系(P <0.05);莎草科主要物种频度、单株重、株高比之间呈极显著正相关关系 (P < 0.01),株高和频度、单株重之间呈极显著正相关关系 (P < 0.01),株高和株高比之间呈显著正相关关系 (P < 0.05);豆科主要物种株高比和株高呈显著负相关关系 (P < 0.05),株高比和单株重及频度和株高之间呈显著正相关关系 (P <0.05);杂类草主要物种的株高和单株重呈极显著正相关关系(P < 0.01),株高和株高比呈极显著负相关(P < 0.01),频度和株高、单株重之间呈显著负相关关系 (P < 0.05)。
3 讨论与结论
草地物种多样性的改变导致草地生态系统在不同利用方式下向不同演替方向变化[25],因而物种多样性的格局及形成机制是生态学研究的焦点[26-28]。研究表明,物种多样性格局的形成取决于两方面:一是以环境筛选为基础的确定性群落构建和以扩散限制为基础的随机性群落构建[26,29-30]的生态过程,二是以不同的功能群组成或不同系统发育阶段的物种组成以及不同的稀有种和常见种组成的相对贡献的物种组成[25,31-32]。其中有关稀有种和常见种对群落多样性格局相对贡献的研究较多,但也颇具争议[33]。一般认为,群落格局主要由大量分布范围较窄且频度较低的稀有种决定,而不是由少数分布范围广且物种频度大的常见种决定[34]。本研究结果表明,分布范围广且物种频度大的禾本科牧草频度和莎草科牧草频度在多个放牧处理下并无明显改变,而适口性较好分布较窄的豆科牧草斜茎黄耆和杂类草异叶青兰的频度在各放牧处理中有所降低,另外,适口性差的星毛委陵菜的频度在各放牧处理中有所增加。主要原因可能是,对于分布范围广频度高的禾本科牧草和莎草科牧草而言其受到的放牧压力轻,生长状况较好,盖度较高,家畜食源充足,而未存在明显的选择性采食。斜茎黄耆和异叶青兰虽频度较高,但其在整个群落中的分布范围窄,生物量占比较低,在适度放牧下其实遭受着高强度的放牧压力,同时牛羊采食对其繁殖结构的破坏极大,影响其种子的形成;而对于适口性较差,依靠根茎繁殖的星毛委陵菜在放牧条件下恰恰为其竞争提供了优势。
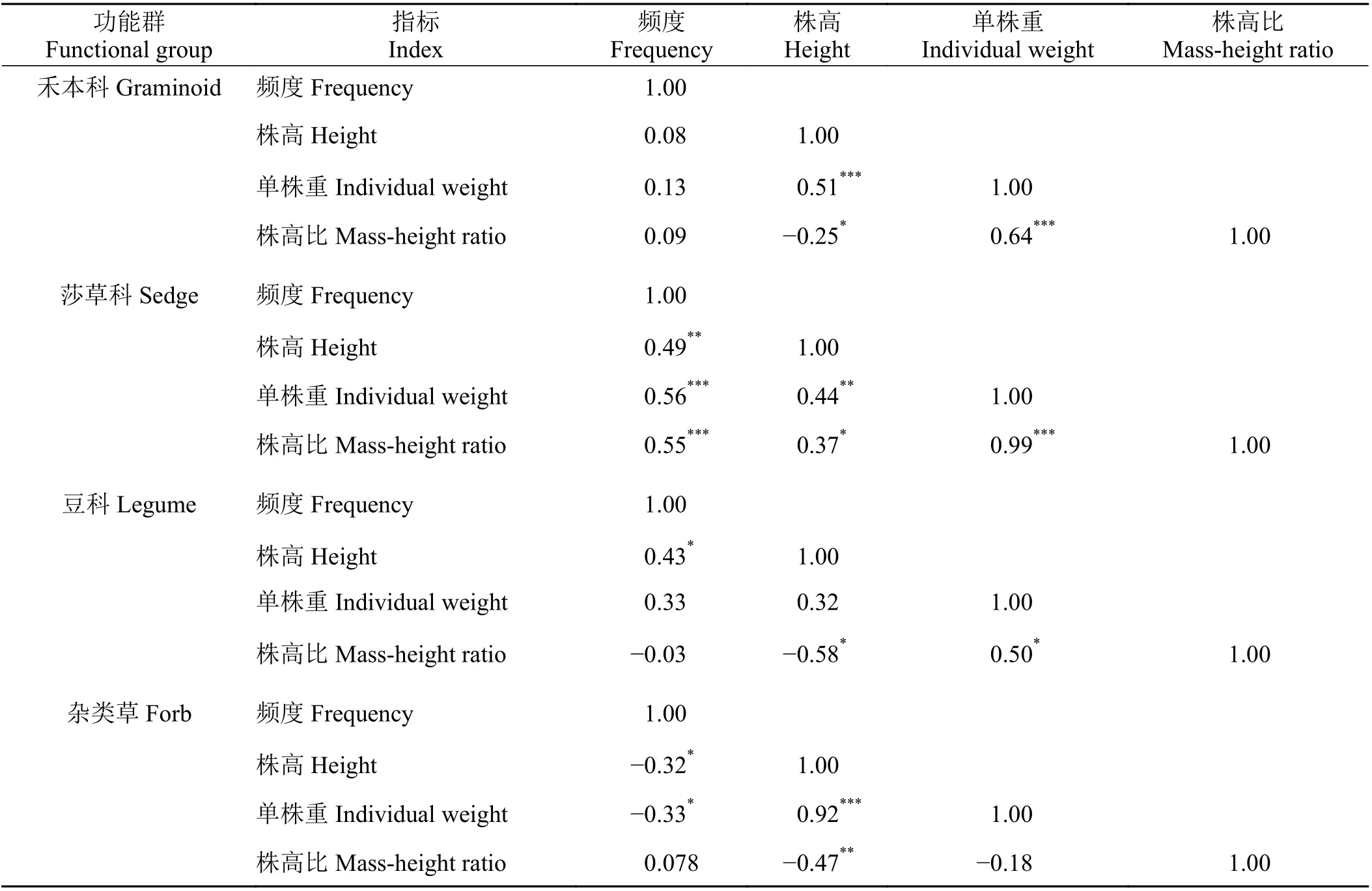
表3 各功能群主要物种各指标之间的相关性分析Table 3 Correlation analysis of major species indexes of each functional group
植株的生长受多方面因素的影响,如气候、降水、放牧等[35-36]。有关放牧和降水相结合的研究表明,在降水丰沛的年份,生长在内蒙古草地上的羊草株高仅在重度放牧下有所下降,而在降水减少的年份,只要存在放牧,羊草株高就会表现为降低,同时降低的程度与放牧强度正相关[37-38]。此外,国内其他区域内的放牧试验研究结果也表明放牧降低了植株高度[8,11-12]。本研究发现,与对照相比禾本科、莎草科、豆科和杂类草等各功能群物种在受放牧干扰后株高均有显著降低,同时不同放牧处理间个别物种的株高也存在差异,如禾本科牧草紫花针茅株高在牦牛和藏羊混牧下其下降较牦牛单牧更为显著,通常藏羊更易取食禾本科牧草[39]紫花针茅,其余3 种禾本科物种在不同放牧处理之间株高并无显著不同,这可能与这几种物种特性相关,这几种物种的株高与紫花针茅相比更低,叶量更少,受到牛羊采食带来的影响相对更少。莎草科优势种矮生嵩草、次优势种干生薹草和豆科类牧草斜茎黄耆的株高在不同的放牧处理之间差异不显著,相比禾本科类株高较高的牧草,放牧对这种低矮型牧草株高的影响有限。杂类草异叶青兰和星毛委陵菜在不同放牧方式下株高呈降低趋势,其中在牦牛单牧中异叶青兰株高显著高于其他放牧处理,主要在于牦牛在食性方面更偏向于选择气味小、低营养高生物量的物种[20]。不同于异叶青兰,放牧对星毛委陵菜的干扰更多的源于牛羊的践踏和粪尿的施入。通常个体在群落当中为获得与其他个体或物种竞争光资源的优势,会在生理方面采取一些竞争性的策略,如调整其在株高方面的资源投入等。植物群落呈垂直分布,各物种处在不同的生态位,在不受放牧干扰的情况下,上层植被对中下层植被产生较大的遮阴作用,上层植被在生长过程中具有较强的光资源获取优势[40-41],中下层植被在光资源的竞争中为了增加竞争优势,会增加在株高方面的资源投入。当群落受到放牧干扰时,上层植被为避免受到家畜的采食,会减少在株高方面的资源投入,同时也有可能是植物受到家畜采食造成损伤[18],使其在株高方面资源投入不足。上层植被的生长受到抑制时,为中下层植被的生长创造了条件,特别有利于丛生型的矮生嵩草和根茎型的星毛委陵菜这类中下层物种的生长和繁殖,在这一竞争过程中中下层植被会将更多资源用于新生株丛的形成。
单株重是植物在干扰条件下物质变化的一项量化性指标,受到干扰后,其变化与株高的变化通常是一致的。根蘖型的禾本科物种,其株高与单株重之间呈极显著正相关关系,在不同放牧方式下有所降低。不同于根蘖型禾本科,丛生型植物和根茎型植物的单株重并不与植物高度变化相一致,这主要是株丛的增大和新植株的产生使植物整株获得更多可分配的资源,这也是当群落高度整体矮化后更有利于矮生物种对草地更快的侵占,以获得更好的竞争优势。牛羊放牧对草地的影响不仅仅限于采食,践踏和粪尿也是草地群落变化的重要因素[42],特别是牛粪对草地的影响,覆盖草地后分解周期长,容易造成草地物种的黄化和死亡[43]。在这种情况下,丛生型矮生嵩草和根蘖型物种星毛委陵菜在该胁迫下的竞争优势和存活优势更高。
株高比是为量化单位高度上生物量的变化,参照了在叶片功能性状属性研究中的比叶面积或比叶重等性状[12]。其目的是通过该性状去理解植物在资源分配过程中的策略、物种对环境变化的响应及物种和生态功能之间的关系。植物在放牧干扰条件下的补偿性生长通常可以用生长状况或生物量的变化去衡量,据此又将植物的生长分为超补偿性生长、等补偿性生长和欠补偿性生长3 类[44-45]。在本研究中,受放牧干扰各功能群大部分物种的生物量(株高比)与对照相比有所增加是植物超补偿性生长的体现。研究发现,与对照相比,除个别禾本科物种的株高比低于对照,各功能群大部分物种生物量(株高比)在不同放牧方式下均有所增加,并且个别物种在不同的放牧处理中差异显著。植物受到外界干扰的情况下通常会表现出避牧性的特征,自身合成一些防御性物质[46-47],减少被采食的几率,或者改变组织内的细胞排列方式,增加单位空间的细胞数量,平衡由于放牧损伤带来的代谢失衡[48],从而增加植物单位体积的质量并增强了植物在生长过程中的优势。放牧方式的不同对各功能群物种生物量(株高比)造成不同的影响,这一结果可能源自牛羊采食过程中的选择性,所以天然草地物种组成和比例的不同也都会影响家畜取食行为和家畜的营养需求,反之高寒草地的物种多样性和物种个体特征也会受到不同类型家畜的影响。当前对于放牧方式对草地生产力和稳定性的作用机制,以及其他放牧方式对草地群落的影响,还需要进一步结合影响草地生产力和结构特征的其他因素、量化草地生产力和结构的其他指标以及放牧试验的研究时限的延长去加以论证。
综上所述,青海湖流域高寒草地植物在物种水平对放牧响应明显,而且不同物种之间差异显著,但在功能群水平对放牧响应较小。就不同的放牧方式而言,牦牛和藏羊1 : 2 混牧处理对各功能群及物种的频度和个体特征影响最大。此外,大部分物种的个体株高比在各放牧处理下均增加,表明中度放牧强度有利于群落主要物种的个体特征向着相对生长速率增加的方向变化,因此在一定程度上解释了高寒草地在中度放牧干扰下的补偿性生长。
猜你喜欢
家庭医药(2021年12期)2021-12-09 08:02:09
家庭医药(2021年23期)2021-10-21 09:39:46
妇女之友(2017年3期)2017-04-20 09:20:00
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
中国药物应用与监测(2015年5期)2015-12-11 03:15:55
农产品市场周刊(2015年39期)2015-10-31 19:12:27
草业学报(2014年3期)2014-11-12 02:07:36
食品科学(2013年14期)2013-03-11 18:25:12
草食家畜(2012年2期)2012-03-20 13:22:34
现代农业科技(2009年9期)2009-02-21 08:49:22
