
西瓜CGMMV胁迫下MIR319家族成员及靶基因的鉴定分析
2021-04-19 03:27孙玉燕张慧青何艳军郭平安
核农学报 2021年5期
孙玉燕 张慧青 范 敏 何艳军 郭平安
(1浙江省农业科学院蔬菜研究所,浙江 杭州 310021;2山西师范大学生命科学学院,山西 临汾 041000)
西瓜(Citrullus lanatusL.)是葫芦科一年生蔓性草本植物,在全世界各地均有种植。目前西瓜生产受到黄瓜绿斑驳花叶病毒(Cucumber green mottle mosaic virus,CGMMV)的严重威胁。CGMMV 属于芜菁花叶病毒科烟草花叶病毒属,为正单链线状RNA 病毒,病毒粒体为杆状,大小为300 nm×18 nm[1]。CGMMV 主要侵染西瓜、甜瓜、黄瓜等葫芦科作物[2],可通过种子、花粉、土壤、介体、机械接触和水源等多种途径进行传播[3-4]。自1935年至今,已报道CGMMV 在全世界30 多个国家和地区均有分布[5-7]。近年,CGMMV 传播速度迅速加快,影响了葫芦科作物尤其是西瓜产业的发展,给商业育种、育苗培育、种子贸易、产品生产和零售等行业带来全球性危害[7]。因此,加强CGMMV的防治,提高西瓜对CGMMV 的抗性是我国乃至全球西瓜生产亟需解决的重要问题。
MiroRNAs(miRNAs)是长度为19~25 nt 调控基因表达的内源小RNA,2002年首次在植物中被报道[8]。MiRNAs 可与靶mRNA 的3′-UTR 特异性配对,引起靶mRNA 的降解或抑制其翻译,进而实现对靶基因的转录后调控。miRNAs 在植物生长发育及抗逆应答中发挥重要的作用[9]。逆境条件下,植物通过调控miRNAs 的表达进一步作用于相应的靶基因以提高植物对胁迫的抗性[10]。MIR319 是植物中一类保守的miRNA 家族,主要调节靶基因TCP和MYB转录因子的表达[11]。MIR319 及靶基因可调节植物叶片发育[12]、花器官发育[13]、次生细胞壁生物合成[14]、细胞增殖[15]及根结发育[16]等生物学过程。
此外,MIR319 及靶基因调控植物对生物胁迫的应答过程。在根结线虫(root knot nematode,RKN)的胁迫下,番茄miR319/TCP4 调节叶片茉莉酸(jasmonic acid,JA)合成基因的表达和内源激素JA 的水平,进而调控对RKN 的抗性[17]。棉花miR159-MYB、miR319-TCP4 和miR167-ARF8 在RKN 胁迫下的表达水平表现为负调控,说明miRNAs 介导的基因调控参与棉花对RKN 的胁迫应答[18]。水稻齿叶矮缩病毒(Rice ragged stunt virus,RRSV)侵染水稻可诱导miR319 的表达,抑制靶基因TCP2 的表达,使内源激素JA 水平降低,促进病毒侵染和症状发展[19]。此外,miR319a可靶向与马铃薯Y 病毒(Potato virus Y,PVY)相互作用的下游赤霉素(gibberellin,GA)信号转导,参与GA的信号转导过程[20]。但目前关于miR319 及靶基因在西瓜CGMMV 胁迫应答中的作用尚不明确。
本研究前期通过对CGMMV 侵染前后的西瓜材料JJZ-M 进行miRNAs 高通量测序,鉴定获得了MIR319家族的3 个成员,miR319、miR319a 及miR319a-3p;其中miR319 在CGMMV 侵染后呈现抑制表达,而miR319a 在CGMMV 侵染后呈现诱导表达[21]。为进一步明确MIR319 在CGMMV 胁迫应答中的作用,本研究从西瓜材料JJZ-M 中分离并克隆MIR319 家族成员的前体基因并对其进行系统进化分析,分析其启动子区的顺式作用元件;对MIR319 的靶基因及剪切位点进行鉴定,对靶基因所编码蛋白进行生物信息学分析;利用转录组测序(RNA-Seq)及实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)对靶基因在CGMMV 不同侵染时间的表达进行分析,旨在为进一步了解MIR319 家族成员及靶基因对CGMMV 胁迫应答的作用机制奠定基础。
1 材料与方法
1.1 植物材料
试验材料为西瓜的高代自交系材料JJZ-M,来源于浙江省农业科学院蔬菜研究所。55℃温汤浸种30 min 后,28℃恒温箱催芽24 h,待种子露白后播种,在植株两片真叶期进行CGMMV 人工摩擦接种。取保存的发病叶片和磷酸盐缓冲液,用研钵研磨成糊状病毒汁液用于接种。分别于接种0 h(CK)、48 h、25 d 取样,各取3 株混成一个样品重复进行RNA-Seq,RNASeq 为1 个生物学重复。
1.2 MIR319 家族前体基因预测及分子克隆
利用Blastn(E-value =1e-2)将MIR319 家族3 个成员(miR319、miR319a 及miR319a-3p)的成熟序列与西瓜基因组数据库(http:/ /cucurbitgenomics.org/organism/1)进行比对,获得与MIR319 家族成员成熟序列完全匹配的基因组序列。截取MIR319 两端150 nt 的核苷酸序列,采用mfold 在线软件进行二级结构分析[22]。若其存在完整的茎环结构且成熟序列完全位于3′或5′端,则预测该茎环结构对应的核苷酸序列为MIR319 的前体基因Pre-MIR319。
根据Pre-MIR319 茎环结构的基因序列,设计特异性引物(F: 5′-AGAGCTTTCTTCAGTCCAC-3′;R: 5′-G GAGCTCCCTTCAGTCCAA-3′)进行PCR 扩增。PCR 反应体系为20 μL,包含2 μL DNA (50 ng·μL-1)、10 μL 2 × TSINGKE Master Mix、 上游和下游引物(10 μmol·L-1) 各0.5 μL、7 μL ddH2O。PCR 反应程序为:94℃预变性4 min;94℃变性30 s,55℃退火30 s,72℃延伸45 s,33 个循环;72℃延伸8 min。PCR 产物经1.0%琼脂糖凝胶电泳检测后,送杭州擎科梓熙生物技术有限公司测序。
1.3 MIR319 前体基因序列及成熟序列比对及系统进化分析
提取miRbase 22.0 数据库[23]中35 个物种的116条miR319 的前体序列,利用MEGA5.1 软件采用Neighbor-Joining 法构建西瓜Pre-MIR319 与其他物种miR319 前体基因的系统进化树。此外,选取来自不同物种的成熟miR319 序列,与西瓜MIR319 成熟序列进行序列比对和系统进化分析。
1.4 MIR319 前体基因启动子区顺式作用元件分析
提取西瓜Pre-MIR319 上游1 500 bp 的序列,利用PlantCARE 软件[24]对其所包含的顺式作用元件进行预测和分析。
1.5 MIR319 靶基因预测及相关生物学分析
根据前期降解组测序的结果(CGMMV 处理前后的叶片混样,1 个生物学重复),对MIR319 的靶基因及其靶切割位点进行分析。利用ScanProsite 对靶基因编码蛋白质的结构域进行分析;ProtParam 对靶基因编码蛋白的氨基酸数目、相对分子量、理论等电点进行分析;TMHMM 预测靶基因编码的蛋白质是否含有跨膜结构域;WolfPSORT 在线软件对靶基因的亚细胞定位进行预测。
1.6 转录组测序(RNA-Seq)及qRT-PCR 分析靶基因在CGMMV 不同侵染阶段的表达
根据本研究前期RNA-Seq 的结果,获得MIR319的靶基因在CGMMV 不同侵染阶段(0 h、48 h 和25 d)的表达,明确MIR319 与靶基因的靶调控关系。此外,进一步利用qRT-PCR 分析MIR319 靶基因在CGMMV不同侵染阶段(0 h、48 h 和25 d) 的表达。利用FastKing RT Kit [天根生化科技(北京)有限公司]将约2 μg 总RNA 反转录成cDNA,以TUA作为内参基因。qRT-PCR使用TransStart Top Green qPCR Supermix 试剂盒(北京全式金生物技术有限公司),在StepOne Plus Real-Time PCR System ( Applied Biosystems,美国)中完成。反应程序:95℃预变性30 s,95℃变性5 s,55℃退火15 s,72℃延伸10 s,40 次循环,3 次生物学重复。采用2-ΔΔCT法[25]计算基因相对表达量。所用引物及序列见表1。

表1 引物及序列Table 1 Primers and sequences
2 结果与分析
2.1 MIR319 家族成员鉴定及表达分析
根据前期对CGMMV 侵染前后的西瓜叶片的miRNAs 高通量测序结果[21]。在两个文库中共获得3个MIR319 家族成员,miR319、miR319a 及miR319a-3p。其中,在CGMMV 侵染25 d 后miR319 呈现抑制表达,而miR319a 呈现诱导表达,miR319a-3p 呈现下调表达(表2),说明MIR319 家族成员对CGMMV 的胁迫呈现不同的响应模式。
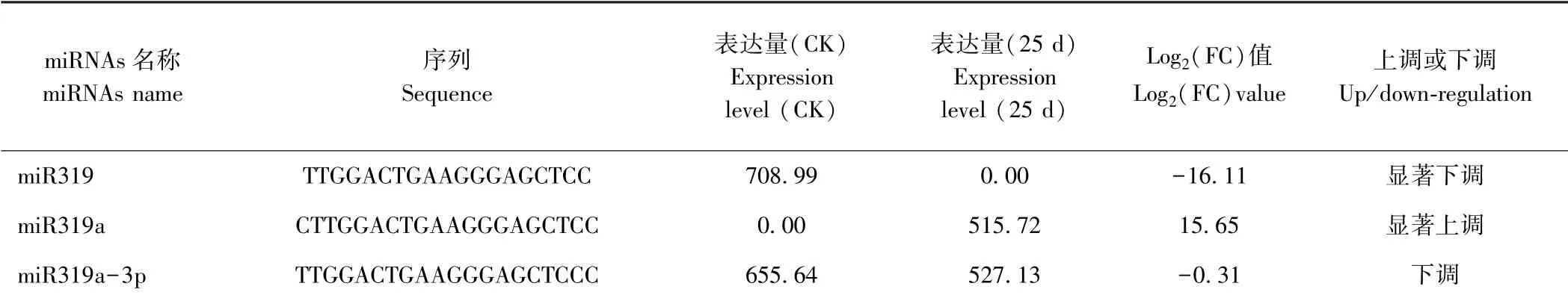
表2 CGMMV 胁迫下MIR319 家族成员鉴定及表达分析Table 2 Identification of MIR319 family member and their expression under the CGMMV stress
2.2 MIR319 家族前体基因预测及克隆
分别将MIR319 家族的3 个成员,miR319(TTGGA CTGAAGGGAGCTCC)、miR319a(CTTGGACTGAAGGG AGCTCC)及miR319a-3p(TTGGACTGAAGGGAGCTC CC)的成熟序列与西瓜基因组数据库(http:/ /cucurbitgenomics.org/organism/1)进行Blast 序列比对。获得与miR319 成熟序列完全匹配的4 条基因组序列,包 括 Chr1: 1904345..1904363、 Chr1: 6347848..6347866、Chr5:29773551..29773569 和Chr7:6929678..6929696;获得与miR319a 成熟序列完全匹配的3 条基因组序列,包括 Chr1: 1904344..1904363、 Chr5:29773551..29773570 及Chr7:6929678..6929697;获得与miR319a-3p 成熟序列完全匹配的2 条基因组序列,包括Chr1:1904345..1904364 及Chr5:29773550..29773569。其 中 仅Chr1:1904345..1904363 (Chr1:1904344..1904363;Chr1:1904345..1904364)及其上、下游序列(各150 bp)能够形成完整的茎环结构,说明miR319、miR319a 及miR319a-3p 的成熟miRNA 序列来源于同一个前体基因Pre-MIR319。以西瓜基因组DNA 为模板对Pre-MIR319 进行PCR 扩增(图1)。通过测序分析,Pre-MIR319 的前体基因长度为170 bp,可形成稳定的茎环结构(图2)。
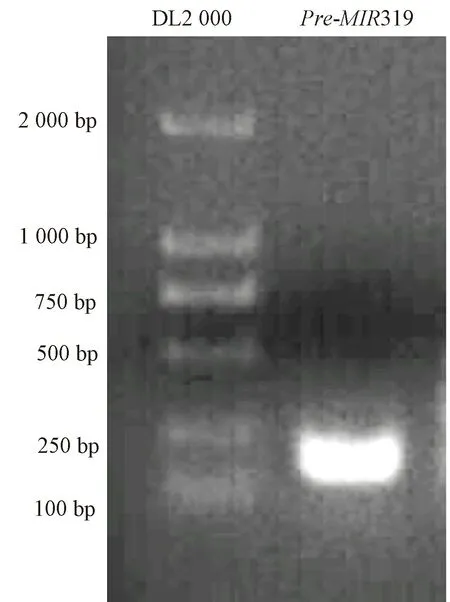
图1 Pre-MIR319 PCR 扩增Fig.1 PCR amplification of Pre-MIR319
2.3 MIR319 家族成熟序列的比对及前体基因系统进化分析
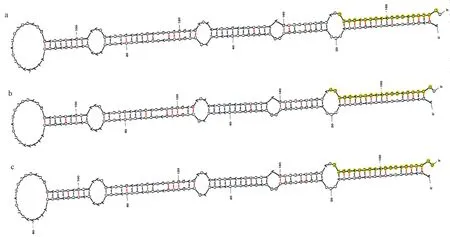
图2 Pre-MIR319 形成的茎环结构及MIR319 家族成员的成熟序列Fig.2 Stem-loop structures of Pre-MIR319 and mature sequences of MIR319 family member
选取miRbase 22.0 数据库中10 个物种的10 条成熟miR319 序列,与西瓜MIR319 成熟序列进行序列比对和系统进化分析。序列比对显示,MIR319 成熟体序列在5′端第2 ~第14 位以及16 ~20 位碱基具有较高的保守性,而在5′端第1、第15、第21、第22 位碱基存在差异(图3)。这些碱基差异可能使不同物种的MIR319 成员作用于不同的靶基因,进而产生不同的功能。选取miRbase 22.0 数据库中35 个物种的116 条MIR319 前体序列,与西瓜Pre-MIR319 一起通过MEGA5.1 进行系统进化分析,可将这些MIR319 前体序列分为四大分支。第一分支含有36 个成员,第二分支含有32 个成员,第三分支含有18 个成员,第四分支含有30 个成员,表明MIR319 在不同物种间的进化关系上存在差异性。其中西瓜Pre-MIR319 位于第一分支,亲缘关系与马铃薯 miR319a 的前体基因(MI0025952)最近(图4)。
2.4 Pre-MIR319 启动子区顺式作用元件分析
对Pre-MIR319 上游1 500 bp 启动子区所含的顺式作用元件进行分析,发现其含有多个顺式作用元件(图5)。包括基因启动子和增强子区的顺式作用元件(TATA-box 和CAAT-box)、光响应元件(TCCC-motif、chs-CMA1a、chs-CMA2a、Ⅰ-box、Box 4、GATA-motif 和GT1-motif)、赤霉素响应元件(P-box 和TATC-box)、乙烯响应元件(ERE)、茉莉酸甲酯响应元件(CGTCAmotif 和TGACG-motif)、类黄酮生物合成基因调控元件(MBSI)、分生组织表达响应元件(CAT-box)以及MYB和MYC 等顺式作用元件。

图3 MIR319 成熟序列比对Fig.3 Alignment of the MIR319 mature sequence
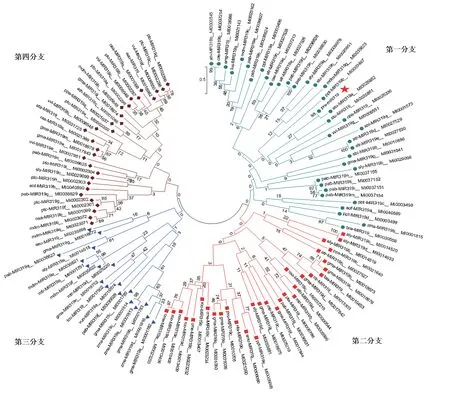
图4 不同物种MIR319 前体基因系统进化分析Fig.4 Phylogenetic relationship of MIR319 precursor gene sequences for diverse species
2.5 降解组测序鉴定MIR319 家族成员的靶基因
由表3 可知,降解组测序对MIR319 家族成员的靶基因进行鉴定,获得miR319 及miR319-3p 的4 个靶基因,分别为Cla002428、Cla013523、Cla019567 和Cla023342,均注释为TCP转录因子;获得miR319a 的2 个靶基因,Cla013523 及Cla013668,其中Cla013523注释为TCP转录因子,Cla013668 注释为MYB转录因子。进一步分析MIR319 对靶基因的剪切位点,发现miR319、miR319a 及miR319-3p 剪切靶基因的位点均位于其成熟序列5′端的第10 位碱基。此外,miR319及miR319a - 3p 剪切靶基因的位点分别位于Cla002428、Cla013523、Cla019567 和Cla023342 的第1 067、 731、1 196 和1 085 位碱基;miR319a 剪切靶基因的位点分别位于Cla013523 及Cla013668 的第732及952 位碱基(表3、图6)。
2.6 靶基因生物信息学分析
利用ScanProsite 对靶基因编码蛋白的结构域进行分析,其中 Cla002428、 Cl013523、 Cla019567 和Cla023342 均含有TCP 结构域,其位置分别位于39 ~97、52 ~110、95 ~153 和44 ~102 位氨基酸,而Cla013668 在33 ~85 位及86 ~140 位氨基酸均含有HTH_MYB 结构域(图7)。ProtParam 对靶基因编码蛋白的氨基酸数目、相对分子量、理论等电点进行分析,靶基因编码氨基酸数目为319 ~554 aa、相对分子量为34.94 ~61.21 kDa、理论等电点为5.29 ~7.80。利用WolfPSORT 在线软件对靶基因进行亚细胞定位预测,其中Cla013523、Cla019567 和Cla013668 定位于细胞核内,Cla002428 和Cla023342 定位于细胞核和细胞质中,TMHMM 预测显示靶基因编码的蛋白质均不含有跨膜结构域(表4)。
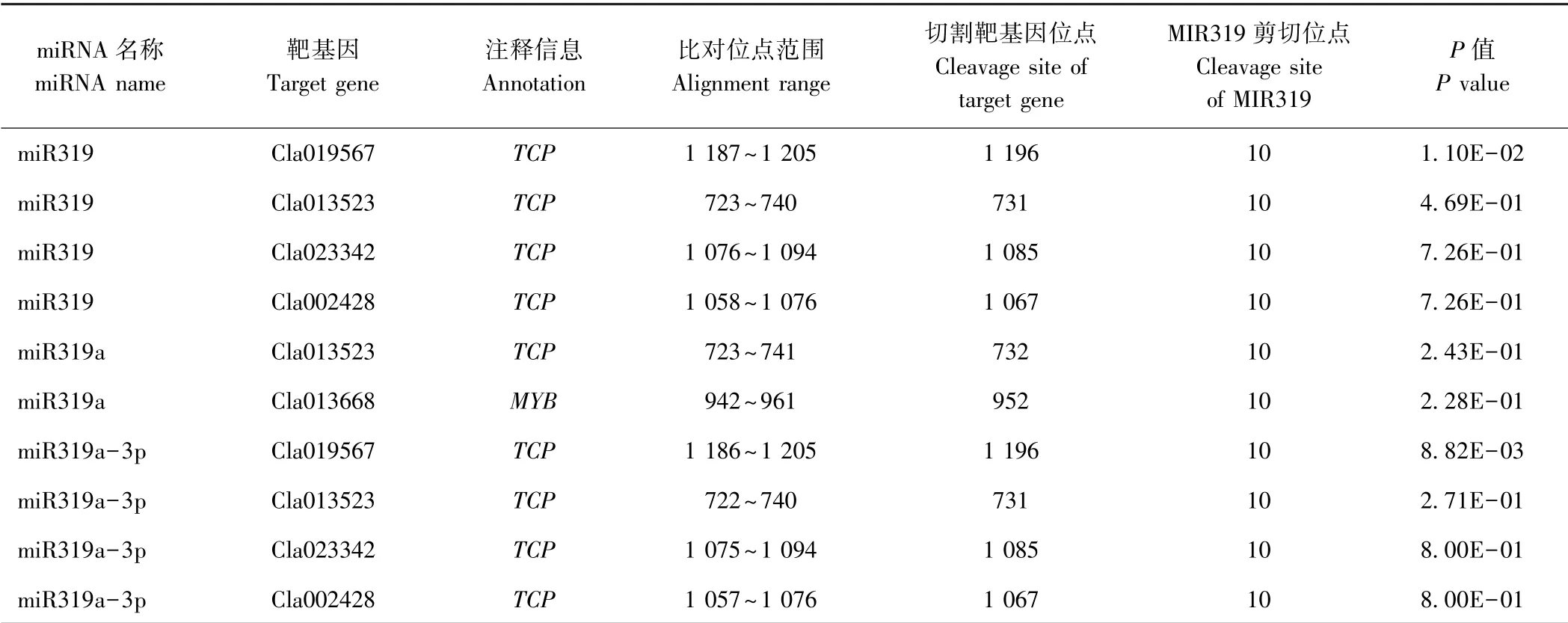
表3 降解组测序获得MIR319 家族成员靶基因Table 3 Target gene of MIR319 family member obtained by degradome sequencing

图5 Pre-MIR319 启动子区顺式作用元件分析Fig.5 Analysis of Pre-MIR319 promoter and its cis-acting regulatory elements
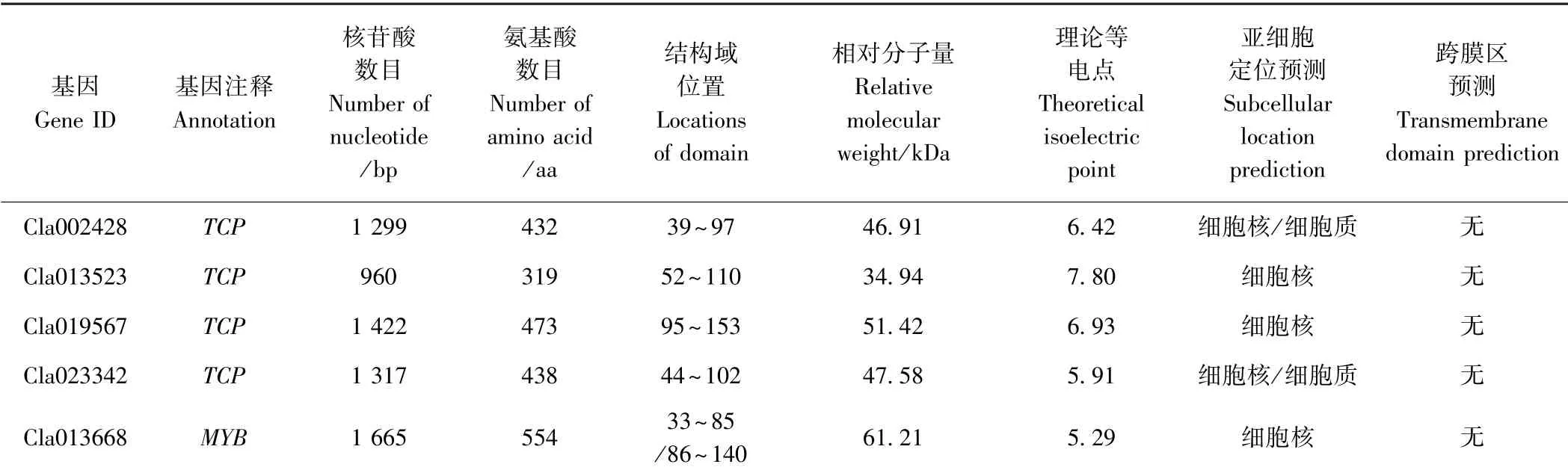
表4 MIR319 靶基因生物信息学分析Table 4 Bioinformatic analysis of MIR319 target genes
2.7 MIR319 靶基因的表达分析
根据CGMMV 侵染前后西瓜叶片的转录组测序结果,对MIR319 靶基因的表达进行分析。在CGMMV侵染48 h 后Cla002428(TCP)、Cla023342(TCP)及Cla013668(MYB)呈现上调表达;Cla013523(TCP)及Cla019567(TCP)呈现下调表达。此外,MIR319 所有靶基因在CGMMV 侵染25 d 后均呈现下调表达,其中Cla013523(TCP)及Cla019567(TCP)呈现显著下调(表5)。根据前期miRNA 高通量测序,MIR319 家族成员(miR319、miR319a 及miR319a-3p)在CGMMV 侵染前后的表达结果(表2),即miR319 及miR319a-3p在CGMMV 侵染25 d 后分别呈现显著下调及下调表达,而miR319 及miR319a-3p 的靶基因Cla002428(TCP)、 Cla013523 (TCP)、 Cla019567 (TCP) 及Cla023342(TCP)在CGMMV 侵染25 d 后呈现下调表达;miR319a 在CGMMV 侵染25 d 后呈现显著上调表达,其靶基因Cla013523(TCP)在CGMMV 侵染25 d表现为显著下调表达,另一个靶基因Cla013668(MYB)在CGMMV 侵染25 d 后表达量无差异。
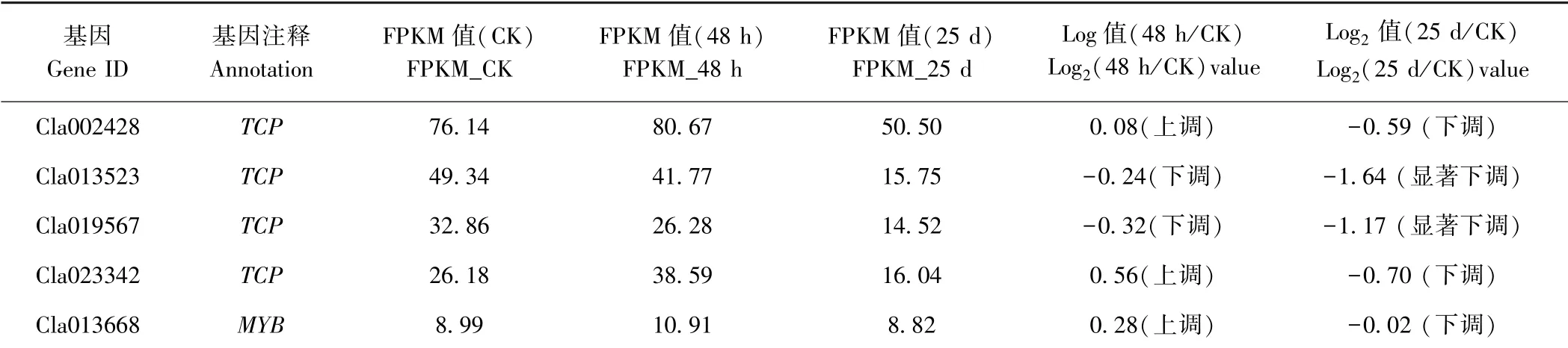
表5 RNA-Seq 分析MIR319 靶基因在CGMMV 不同侵染阶段的表达Table 5 Expression of MIR319 target genes during the process of CGMMV infection by RNA-Seq
此外,利用qRT-PCR 对MIR319 靶基因在CGMMV 不同侵染阶段的表达进行分析,其中Cla013523(TCP)和Cla019567(TCP)在CGMMV 不同侵染阶段的表达模式一致,即与CK 相比,Cla013523(TCP)和Cla019567(TCP)在CGMMV 侵染48 h 和25 d 后呈现持续下调表达;Cla002428(TCP)和Cla023342(TCP)在CGMMV 不同侵染阶段的表达模式一致,即与CK 相比,Cla002428(TCP) 和Cla023342(TCP)CGMMV 侵染48 h 后呈现上调表达,在CGMMV 侵染25 d 后呈现下调表达;与CK 相比,Cla013668(MYB)在CGMMV 侵染48 h 和25 d 后呈现上调表达(图8)。qRT-PCR 结果表明,绝大部分靶基因在CGMMV 不同侵染阶段的表达与转录组测序一致。上述转录组测序结果及qRT-PCR 结果均表明,miR319a 对靶基因Cla013523(TCP)的表达表现为负调控。

图6 降解组测序鉴定MIR319 剪切靶基因的位点Fig.6 Cleavage sites of target gene predicted by degradome sequencing
3 讨论
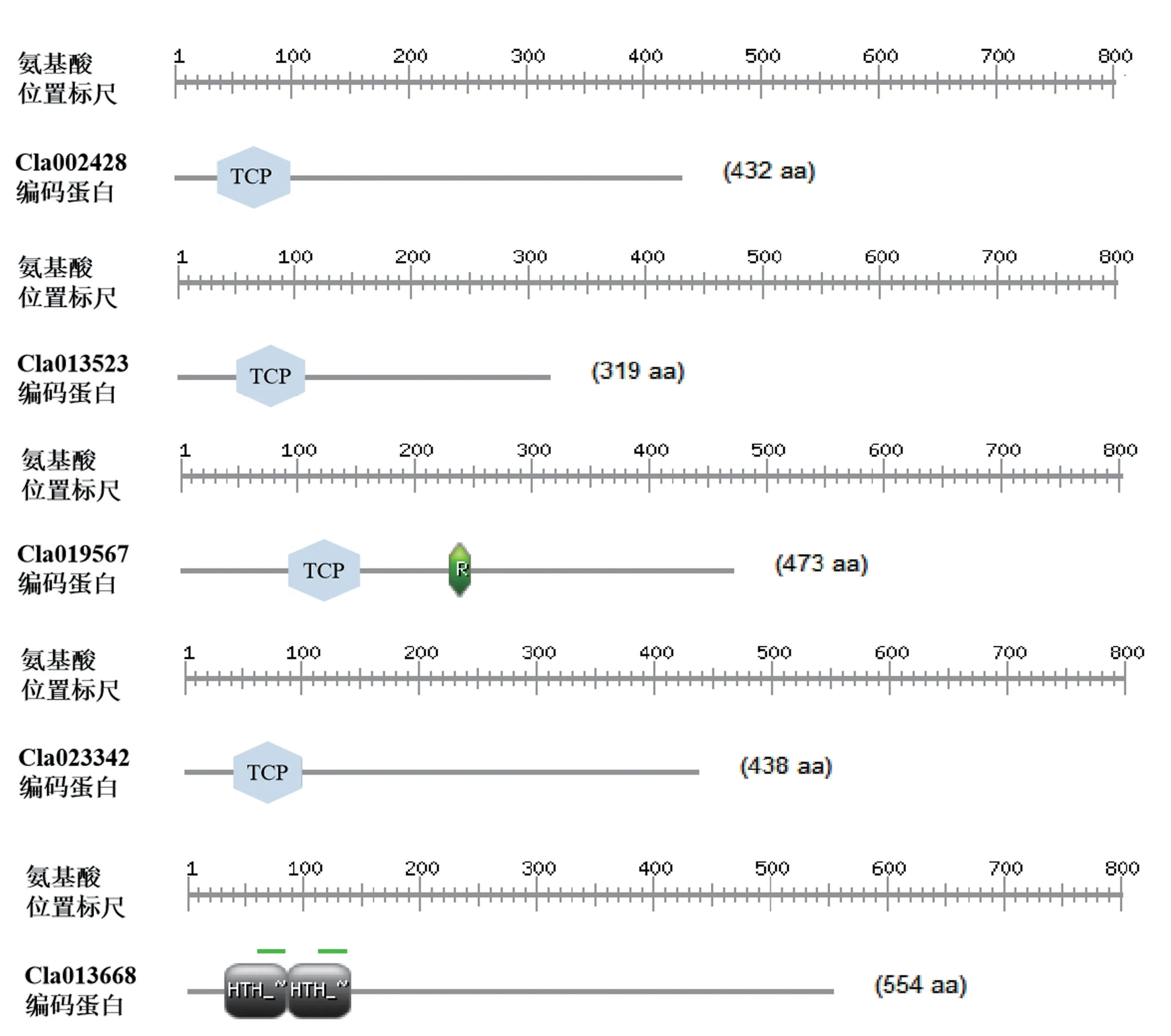
图7 ScanProsite 预测MIR319 靶基因编码蛋白结构域Fig.7 Domain prediction of MIR319 target gene using ScanProsite

图8 qRT-PCR 分析MiR319 的靶基因在CGMMV 不同侵染阶段的表达Fig.8 Expression analysis of MIR319 target genes during the process of CGMMV infection by qRT-PCR
目前,参与植物对CGMMV 胁迫应答的miRNAs越来越多地被挖掘出来。对接种CGMMV 前后的黄瓜进行miRNAs 高通量测序,获得多个参与CGMMV 胁迫应答的新的保守的miRNAs 成员[26]。本研究前期通过对CGMMV 侵染前后的西瓜叶片进行miRNAs 高通量测序,获得一些参与CGMMV 胁迫应答的miRNAs,如miR164b 及miR319[21]。miR164b 通过靶向NAC转录因子参与西瓜对CGMMV 的胁迫应答[27]。研究表明,miR319 在植物对生物胁迫应答中发挥重要的作用[17-20]。水稻齿叶矮缩病毒(RRSV)侵染可诱导miR319 的表达,抑制靶基因TCP21 的表达,JA 水平降低,表明RRSV 侵染可诱导miR319 介导的防御机制[19]。因此,进一步对miR319 及靶基因进行相关生物学分析可为揭示miR319 对CGMMV 胁迫应答的分子机制奠定基础。本研究对鉴定到的西瓜MIR319 前体基因Pre-MIR319 进行序列分析,显示Pre-MIR319长度为170 bp,可形成稳定的二级发夹结构。西瓜MIR319 成熟序列在5′端第2 ~第14 位碱基具有较高的保守型,可保证MIR319 对靶基因的精确切割。对MIR319 前体序列进行系统进化分析,显示Pre-MIR319 进化关系与马铃薯miR319a 的前体基因(MI0025952)最近。
研究表明miR319 与其他miRNAs 存在相互作用。植物miR159 主要靶向MYB转录因子,miR319 主要作用于TCP转录因子。研究发现miR319 家族与miR159 家族关系密切,它们的靶基因在21 个核苷酸中有17 个具有共享序列,miR159 和miR319 的功能特化是通过表达差异和序列差异实现的[28]。本研究中西瓜miR319 的2 个靶基因Cla013523 和Cla019567,同时也作为miR159 的靶基因,受到miR159 和miR319的共同调控。除此之外,miR319 与miR172 也存在相互作用,miR319 前体异常的长折叠结构与miR172 的加工有关[29]。
MIR319 和TCP的mRNA 几乎互补的核苷酸序列构成了基因调控的分子机制[30]。MIR319 单核苷酸突变可降低其靶向5 个TCP基因的能力[13]。本研究中,西瓜MIR319 家族的3 个成员靶向4 个TCP转录因子基因,1 个MYB转录因子基因,剪切位点在MIR319 家族成员成熟序列5′端第10 位碱基。根据转录组测序的结果,仅miR319a 与靶基因Cla013523(TCP)的表达表现为负相关。分析其原因主要是同一个靶基因受不同miRNAs 的调控,如Cla019567(TCP)除受miR319和miR319a-3p 的调控,还受miR159 的调控,调控机制较杂,需进一步研究。
研究发现miR319 的靶基因TCP调控JA 的生物合成及信号转导过程。RRSV 侵染一方面诱导miR319 的表达,抑制TCP21 的表达,另一方面水稻内源JA 水平降低,表明RRSV 可诱导JA 介导的防御机制,调控水稻对RRSV 的抗性[19]。本研究通过对Pre-MIR319 启动子区的顺式作用元件进行分析,发现其启动子区含有茉莉酸甲酯响应元件CGTCA-motif 和TGACG-motif,说明西瓜MIR319 也可能通过参与JA介导的防御反应调控西瓜对CGMMV 的胁迫应答。
4 结论
本研究获得西瓜MIR319 家族3 个成员的前体基因序列,系统进化分析将多个物种的miR319 前体基因分为四个分支。Pre-MIR319 启动子区含有多个参与植物生长发育及胁迫应答的顺式作用元件,降解组测序获得MIR319 家族3 个成员的5 个靶基因,其中4个为TCP转录因子,1 个为MYB转录因子。转录组测序及qRT-PCR 对MIR319 的靶基因在CGMMV 胁迫下的表达进行分析,miR319a 对靶基因Cla013523(TCP)的表达表现为负调控。本研究明确了MIR319 家族成员及其靶基因对CGMMV 的应答模式,为西瓜CGMMV 抗性基因(因子)获得及分子机理研究奠定了基础。
猜你喜欢
传感器世界(2022年6期)2022-11-25
当代水产(2022年1期)2022-04-26
空气动力学学报(2022年1期)2022-03-16
天津医科大学学报(2021年1期)2021-12-05
空气动力学学报(2021年2期)2021-05-04
科学大观园(2020年22期)2020-11-30
中国瓜菜(2019年8期)2019-09-19
科技与创新(2019年16期)2019-08-26
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
