
生物炭施用下潮土团聚体微生物量碳氮和酶活性的分布特征
2021-04-15 09:23成宇阳王秀斌张水清
植物营养与肥料学报 2021年3期
张 帅,成宇阳,吴 行,郑 琴,王秀斌*,张水清
(1 中国农业科学院农业资源与农业区划研究所,北京 100081;2 河南省农业科学院植物营养与资源环境研究所,河南郑州 450002)
土壤团聚体是土壤微生物活动的主要场所,在土壤养分周转中起核心作用[1]。土壤微生物量碳(MBC) 和微生物量氮 (MBN) 含量是评价土壤肥力变化和扰动的一个重要指标,与土壤团聚体数量及质量之间存在密切联系[2]。已有大量研究报道了不同土壤类型、不同养分管理方式下 (单施化肥、有机无机肥配施) 土壤MBC、MBN在团聚体水平上的分异特征[3-4]。近年来,生物炭作为一种富含碳的有机物料被广泛应用于农田生态系统中,生物炭与土壤颗粒可形成团聚体和有机–无机复合体,有利于团聚体的形成与稳定[5]。基于Meta-analysis方法的研究结果显示,在培养、盆栽及田间试验下添加生物炭多数能提高土壤MBC含量,而盆栽和大田试验中生物炭对土壤MBN含量影响的报道却不尽相同[6],其主要原因归于植物对土壤氮的竞争[7]。但迄今为止,关于土壤MBC、MBN含量对生物炭添加的响应特征的研究多集中在整体土壤上,而在土壤团聚体水平上则报道较少。
土壤酶是土壤碳、氮转化的重要驱动因子,其活性可作为评价土壤碳、氮转化能力的重要指标[8]。由于不同粒径团聚体理化性状存在巨大差异,导致土壤微域中酶学反应条件也显著不同[9],一般来说,β-葡萄糖苷酶、β-纤维二糖苷酶、β-木糖苷酶、乙酰氨基葡萄糖苷酶等碳水化合物酶的活性随粒级由粗砂到粘粒的变化呈下降趋势[10],而参与氮、磷转化的酶活性则在粉粒和粘粒中占主导地位[10-11]。目前,关于土壤团聚体转化酶、脲酶、蛋白酶及磷酸酶等变化特征的研究多集中在其对不同耕作制度和不同养分管理方式 (秸秆还田、有机无机肥配施)的响应[10, 12]。而有关生物炭如何影响参与碳氮转化的土壤酶活性的研究主要集中在整体土壤上,有研究发现,生物炭施用增加了参与氮转化的土壤酶活性,降低了参与碳转化的土壤酶活性[13]或具有不确定性[14],这些差异性结论主要取决于生物炭用量、土壤类型以及酶特性[13, 15]。但目前为止,有关生物炭配施化肥对土壤团聚体中参与碳氮转化的酶活性的影响报道较少。
因此,以华北平原石灰性潮土为供试土壤,以花生壳生物炭为供试材料,采用两年田间微区试验,研究生物炭与化肥添加下土壤不同粒径团聚体中微生物量碳、氮含量以及胞外酶活性的变化特征,并阐明在团聚体中影响胞外酶活性变化的主控因素,旨在为改善小麦–玉米种植体系下土壤微环境,提升土壤质量提供科学依据。
1 材料与方法
1.1 试验设计
试验地点位于河南省平原新区河南现代农业研究基地 (35°0'N、113°43'E),该地属于北温带大陆性季风气候,四季分明,雨热同期,降水集中在夏季,全年平均降雨量542.15 mm,平均气温15.6℃,无霜期209 天。8月份最热,月平均气温25.9℃;1月份最冷,月平均气温2.15℃。全年日照时间约1869.7 h。供试土壤类型为石灰性潮土,有机质含量10.69 g/kg、全氮含量 0.44 g/kg、有效磷含量 4.41 mg/kg、速效钾含量 91.16 mg/kg、pH 8.14、土壤容重1.18 g/cm3。种植方式为冬小麦–夏玉米轮作。所施生物炭为河南三利新能源公司生产,采用花生壳在500℃高温厌氧条件下热解4 h制得,其物理化学性质为 pH 9.16、碳 717.60 g/kg、氮 18.51 g/kg、氢26.11 g/kg、氧 124.91 g/kg、磷 1.76 g/kg、钾 5.35 g/kg、比表面积 5.08 m2/g、平均孔径 8.18 nm。
田间试验开始于2017年小麦季,试验设置4个处理:不施肥 (CK)、单施化肥 (NPK)、单施生物炭(BC)、生物炭配施化肥 (NPK+BC)。每个处理3次重复,随机区组排列,小区面积为 4 m2(长宽均 2 m),用1 m深的水泥板将各试验小区隔开以防止相邻小区之间的相互影响。花生壳生物炭施用量为22.5 t/hm2,仅在2017年小麦播种前施用一次。小麦季氮磷钾肥料用量分别为纯 N 165 kg/hm2、P2O582.5 kg/hm2和 K2O 82.5 kg/hm2,施氮方式为基肥和拔节期追肥之比为1∶1,磷肥和钾肥全部基施;玉米季施氮量为纯氮225 kg/hm2,基肥和大喇叭口期追肥之比为7∶3,磷、钾肥施用同小麦季。施用的氮、磷、钾肥分别为尿素 (N 46%)、过磷酸钙 (P2O512%)、氯化钾 (K2O 60%)。
1.2 土壤样品采集与团聚体分级
于2019年9月玉米收获后采集各小区的耕层土壤样品。耕层土壤团聚体样品为每个小区随机取两个点的耕层原状土 (长 18 cm×宽 8 cm×高 12 cm)。土样运回实验室后,将各小区取的两点土样混合,并将大块土壤沿土块本身的裂隙分开,过10 mm筛后存放于4℃冰箱备用。
土壤团聚体分级采用鲜土湿筛法[16]。具体步骤为:每次称取相当于100 g干土的过筛鲜土,均匀覆盖于套筛最上层 (套筛孔隙从上到下分别为2 mm、0.25 mm、0.053 mm),将套筛置于湿筛装置 (型号DM200-Ⅲ团粒分析仪,上海德玛信息技术有限公司)中,蒸馏水浸泡5 min后筛分,振动频率为25次/min,幅度为3 cm;筛分后收集各级筛子上的团聚体,粒径 < 0.053 mm的团聚体溶液通过静置沉降后离心获得,最终得到粒径 > 2 mm 团聚体、2~0.25 mm 团聚体、0.25~0.053 mm 微团聚体以及 < 0.053 mm 粉粘粒,重复上述步骤多次,直至收集足够多的土壤团聚体鲜样,其中一部分鲜样于4℃下保存,用于微生物量碳、氮 (MBC、MBN) 和胞外酶活性的测定;另一部分收集到已称重的铝盒中,于60℃下烘干后称重,用于有机碳 (SOC) 和全氮含量的测定。
1.3 测定方法
耕层土壤养分含量参照《土壤农化分析》[17]中的常规方法测定。其中,土壤有效磷含量采用NaHCO3(0.5 mol/L) 浸提—钼锑抗比色法测定;土壤速效钾含量采用 NH4OAc (1 mol/L) 浸提—原子吸收分光光度计测定;土壤有机碳含量采用重铬酸钾容量法—外加热法测定,全氮含量采用凯氏蒸馏法测定。土壤铵态氮 (NH4+-N) 和硝态氮 (NO3–-N) 含量采用0.1 mol/L氯化钙溶液浸提—流动分析仪 (SEALAA3)测定。耕层土壤及各粒级团聚体的MBC、MBN含量采用氯仿熏蒸—K2SO4提取法测定[18]。
各粒级团聚体胞外酶活性的测定:参与土壤C、N、P循环的β-葡糖苷酶、β-纤维二糖苷酶、α-葡糖苷酶、β-木糖苷酶、乙酰氨基葡糖苷酶、亮氨酸氨基肽酶以及磷酸酶等采用荧光微孔板酶检测法[19],根据不同酶的标准底物被水解后能产生荧光物质4-甲基伞形酮 (4-methylumbelliferyl) 或 7-氨基-4-甲基香豆素 (7-amino-4-methylcoumarin) 的原理,通过测定其荧光值的强度来表征土壤胞外酶活性。主要步骤如下:称取相当于1.0 g干土的新鲜土样,加入50 mmol/L醋酸盐缓冲液100 mL混匀,用磁力搅拌器搅拌以维持土壤悬浊液的均匀度;然后将缓冲液、土壤样品悬浊液、10 μmol/L标准品和200 μmol/L相应底物严格按照一定体积和顺序加入到黑色的96孔酶标板中;酶标板封口后避光培养4 h (25℃),上机前全部加入 10 μL 1.0 mol/L NaOH 溶液后立刻在激发波长为365 nm、发射波长为450 nm的条件下测定荧光 (多功能酶标仪 Scientific Fluoroskan Ascent FL,Thermo)。酶活性[nmol/(g·h)]计算公式如下:
淬灭系数 (flour./nmol)=(标准对照–样品对照)/标准荧光
激发系数 (flour./nmol)=标准荧光/0.5 nmol
净荧光值 (flour.)=(样品荧光–样品对照)/淬灭系数–底物对照
胞外酶活性=净荧光值×100 mL×100/[激发系数×0.2 mL×土样重 (g)×时间 (h)]
脲酶活性参照Guo等[20]的方法,采用土壤脲酶(S-UE) 试剂盒 (北京索莱宝科技有限公司) 检测,酶活性单位用 NH3-N μg/(g·d) 表示。
1.4 数据处理
采用Excel 2019对试验数据进行整理并作图,使用 IBM SPSS Statistics 22 进行方差分析 (Duncan test,P=0.05),单因素方差分析用于比较各处理之间以及各粒径团聚体之间的差异性,双因素方差分析用于分析生物炭、化肥及其交互作用对耕层土壤理化性质的影响。运用Canoco 5软件进行主成分分析 (PCA) 和冗余分析 (RDA)。
2 结果与分析
2.1 耕层土壤养分含量
双因素方差分析结果 (表1)表明,施用NPK对土壤pH、有效磷、速效钾、硝态氮和全氮含量有显著影响,施用生物炭 (BC) 对土壤速效钾、有机碳和全氮含量有显著影响,BC和NPK的交互作用对全氮含量也有显著影响 (P< 0.05)。与 CK 相比,NPK、BC+NPK处理均显著降低了土壤pH,提高了有效磷、速效钾和硝态氮含量,对铵态氮没有显著影响;BC处理的土壤pH和有效磷及矿质氮含量与对照相比没有明显差异,但速效钾、有机碳和全氮含量显著提高(P< 0.05)。此外,与NPK处理相比,BC+NPK处理显著提高了有机碳和全氮含量,而土壤速效养分没有明显变化;与BC处理相比,BC+NPK处理显著提高了有效磷和硝态氮含量。
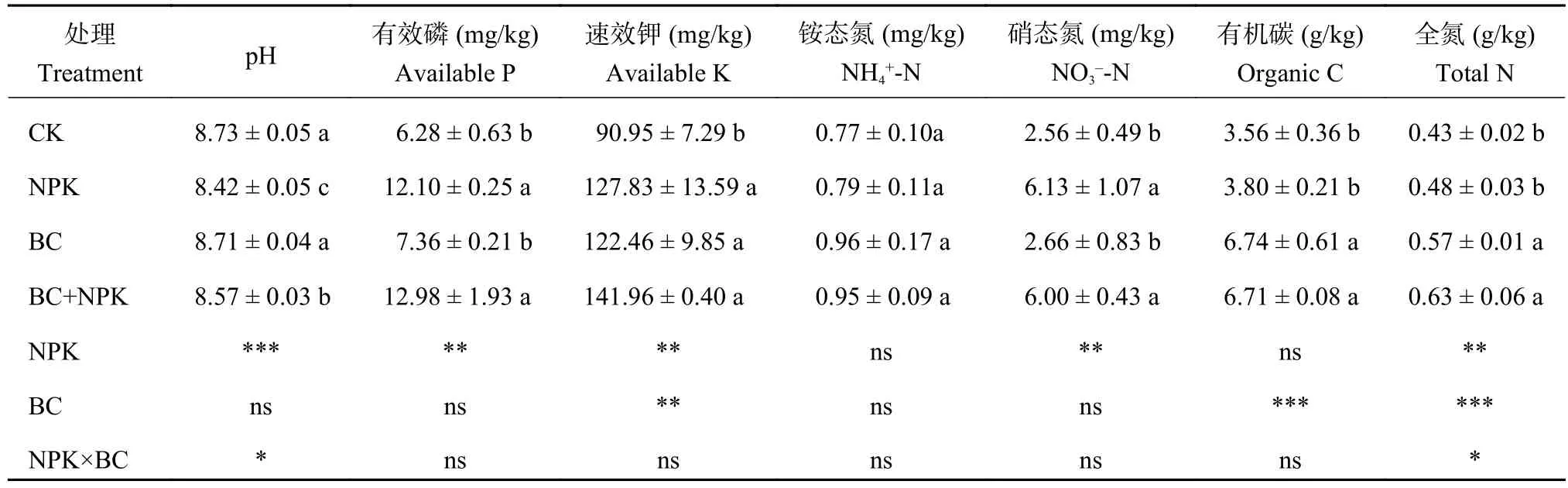
表 1 不同处理耕层土壤养分含量Table 1 Nutrient contents of plough layer soils under different fertilization
2.2 团聚体中微生物量碳、氮含量及微生物熵
土壤MBC和MBN含量在粒径2~0.25 mm团聚体中最高,> 2 mm团聚体次之,0.25~0.053 mm和< 0.053 mm 团聚体中的含量较低 (P< 0.05,图1)。BC或NPK处理的各粒级团聚体中MBC含量与CK相比没有明显变化;BC+NPK处理较CK显著提高了粒径 > 2 mm和0.25~0.053 mm 团聚体中的MBC含量,增幅分别为59.57%和34.68%,较NPK 处理显著提高了粒径 > 2 mm和< 0.053 mm 团聚体中的 MBC 含量 (P< 0.05);而耕层土壤的MBC含量在不同处理之间无显著变化 (图1)。同样,与CK相比,单施NPK处理显著提高了粒径2~0.25 mm团聚体的MBN含量,单施BC处理显著降低了粒径2~0.25 mm团聚体的MBN含量;与CK和BC处理相比,BC+NPK处理显著提高了耕层土壤以及粒径 > 2 mm、2~0.25 mm 团聚体的MBN含量(P< 0.05),较CK分别增加了17.33%、42.24%和19.28%;粒径 < 0.053 mm 团聚体的MBN含量在不同处理之间无显著变化 (图1)。
对于MBC/MBN值,单施BC处理相较于C K显著增加了粒径 2~0.2 5 m m团聚体的MBC/MBN值,单施NPK处理较CK显著降低了 粒径 < 0.53 mm 团聚体的 MBC/MBN 值 (图 1)。与CK相比,NPK、BC、BC+NPK处理相比均显著增加了粒径 > 2 mm团聚体的微生物熵,且BC+NPK处理相较于NPK和BC处理也显著提高了粒径> 2 mm团聚体的微生物熵,但施用生物炭处理(BC、BC+NPK) 与CK和NPK处理相比明显降低了耕层土壤及粒径2~0.25 mm和0.25~0.053 mm团聚体的微生物熵 (P< 0.05,图 1)。
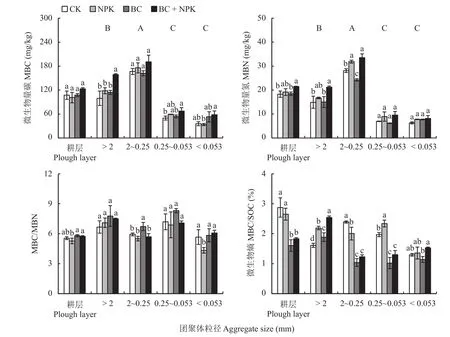
图 1 不同处理耕层土壤及各粒径团聚体中MBC和MBN含量和MBC/MBN、微生物熵 (MBC/SOC)Fig.1 The contents of MBC and MBN and the ratios of MBC/MBN and MBC/SOC in plough layer soil and different particle-size fractions under different fertilization
2.3 团聚体中胞外酶活性
不同粒径团聚体中参与碳循环的酶活性差异显著 (P< 0.05),β-葡糖苷酶、β-纤维二糖苷酶、α-葡糖苷酶和β-木糖苷酶活性均呈现出在粒径2~0.25 mm 团聚体中最高,> 2 mm 团聚体次之,0.25~0.053 mm 团聚体较低,而在粒径 < 0.053 mm 团聚体中最低 (表2)。与CK相比,NPK处理显著提高了粒径2~0.25 mm和0.25~0.053 mm团聚体中这四种酶的活性,BC处理显著提高了粒径0.25~0.053 mm团聚体中的β-葡糖苷酶活性,BC+NPK处理显著提高了粒径 > 2 mm、2~0.25 mm和0.25~0.053 mm 团聚体的这四种酶活性;此外,BC+NPK处理较BC处理显著提高了粒径 > 2 mm和2~0.25 mm 团聚体的这四种酶活性 (P< 0.05)。
与氮、磷转化相关的乙酰氨基葡糖苷酶、亮氨酸氨基肽酶、脲酶以及磷酸酶活性亦呈现出粒径2~0.25 mm 团聚体中最高,> 2 mm 团聚体中次之,但其在粒径 0.25~0.053 mm和< 0.053 mm 团聚体中的活性相差不大 (表2)。NPK、BC+NPK处理与CK相比均显著提高了粒径2~0.25 mm和0.25~0.053 mm团聚体中的磷酸酶活性,并显著提高了粒径 > 2 mm团聚体中的脲酶活性和粒径0.25~0.053 mm团聚体中的乙酰氨基葡糖苷酶活性 (P< 0.05)。对于亮氨酸氨基肽酶,BC+NPK处理较CK显著提高了粒径 > 2 mm和0.25~0.053 mm 团聚体中该酶活性,较BC处理显著提高 > 2 mm团聚体中该酶活性(P< 0.05,表 2)。粒径 < 0.053 mm 团聚体中各酶活性在不同处理间无显著变化。
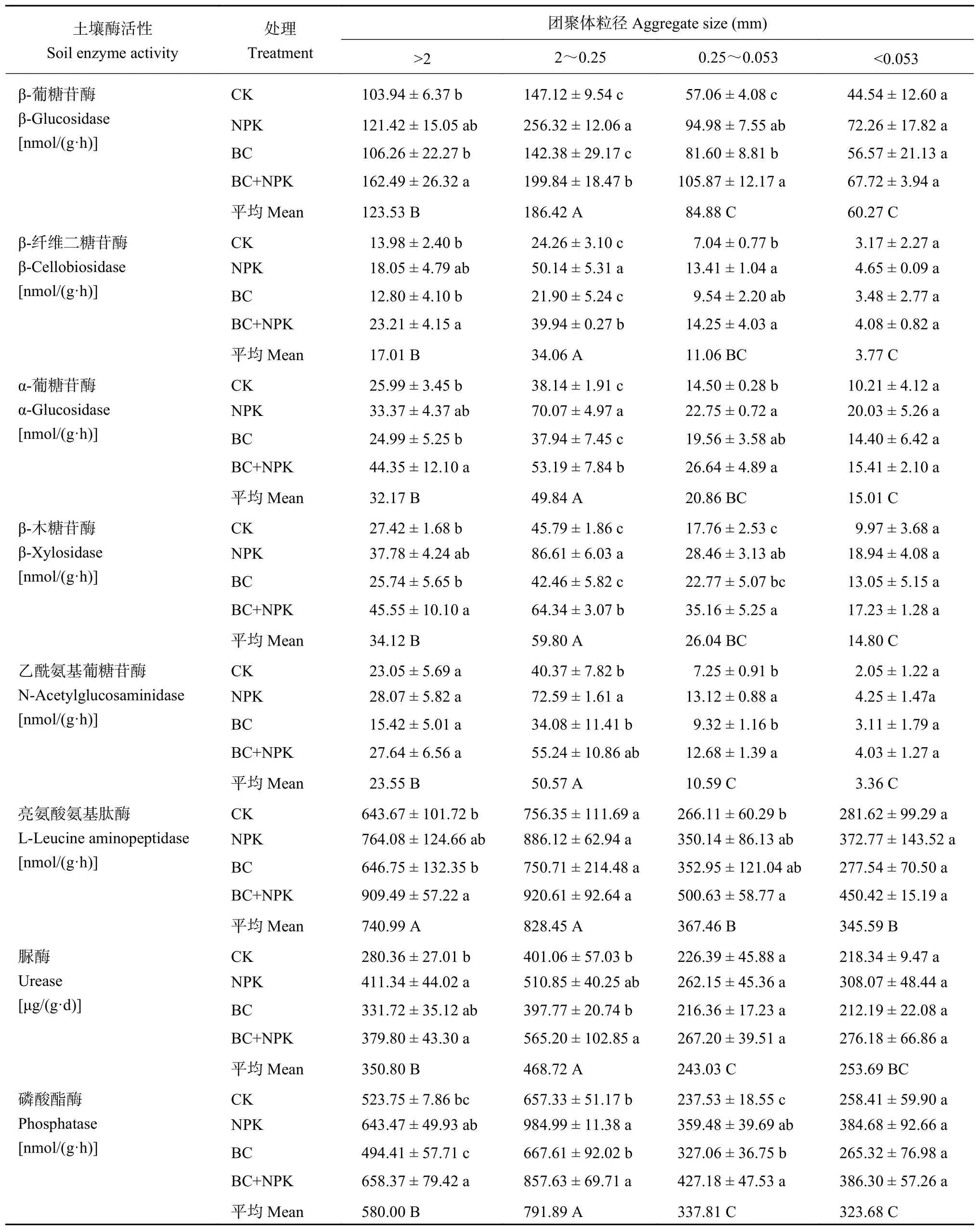
表 2 不同处理各粒径团聚体中胞外酶活性Table 2 Extracellular enzyme activities in particle-size fractions under different fertilization
2.4 团聚体酶活性的主成分分析及冗余分析
不同处理间土壤团聚体酶活性的主成分分析(PCA) 显示,PC1和PC2分别占总变异的88.63%和5.27%,粒径 > 2 mm、2~0.25 mm 团聚体与0.25~0.053 mm、< 0.053 mm 团聚体在第一主成分上被明显分开,团聚体粒径对酶活性影响较大 (图2)。通过冗余分析 (RDA) 表征团聚体酶活性与土壤环境因子的相关性,发现RDA1和RDA2分别解释了酶活性总变异的77.12%和2.54%,土壤团聚体中的MBC、MBN以及MBC/MBN值与团聚体酶活性变化显著相关,其中MBC对团聚体酶活性变化的贡献最大 (图 3)。
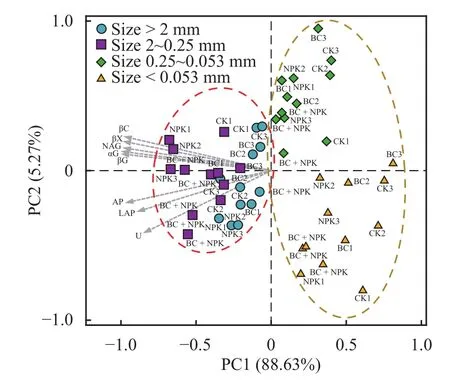
图 2 土壤团聚体酶活性的主成分分析Fig.2 Principal component analyses of enzyme activities in particle-size fractions
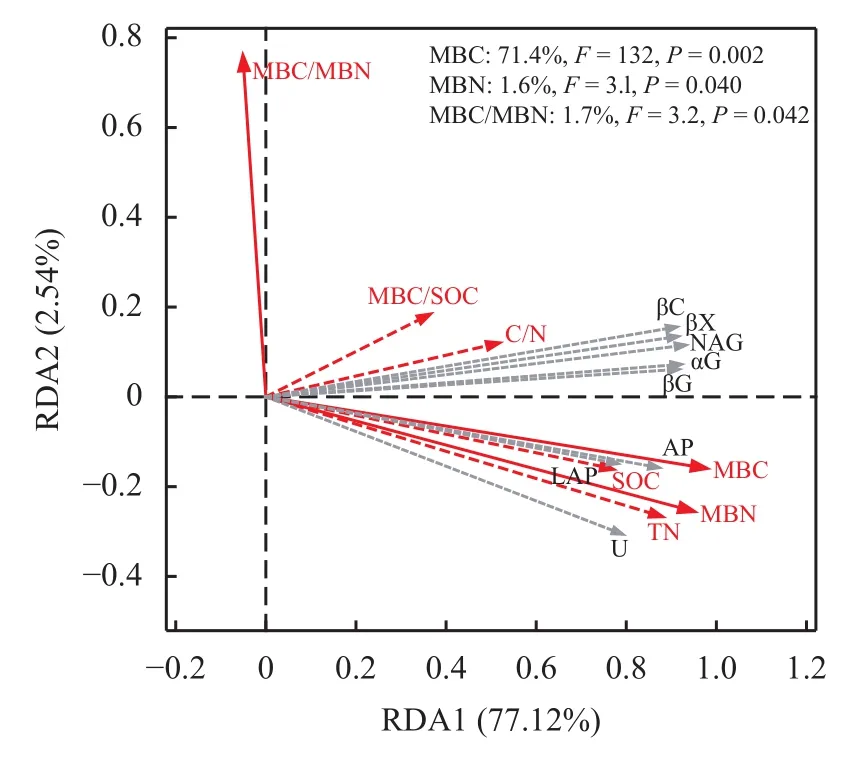
图 3 土壤团聚体酶活性与环境因子的相关性Fig.3 Correlations between enzyme activities and environmental factors at the aggregate scale
此外,通过RDA分析了每个粒径团聚体中的酶活性与环境因子的相关性,发现不同粒径团聚体中影响酶活性的环境因子各不相同 (图4)。粒径> 2 mm团聚体酶活性变化与该粒径团聚体微生物熵、全氮和MBC显著相关,其中微生物熵对该粒径团聚体酶活性变化的贡献最大;而粒径2~0.25 mm团聚体酶活性的变化与该粒径团聚体MBC/MBN显著相关;粒径0.25~0.053 mm团聚体酶活性变化与该粒径团聚体MBC含量显著相关;粒径 < 0.053 mm团聚体中的酶活性与该粒径团聚体碳氮以及微生物量碳氮含量均不相关。
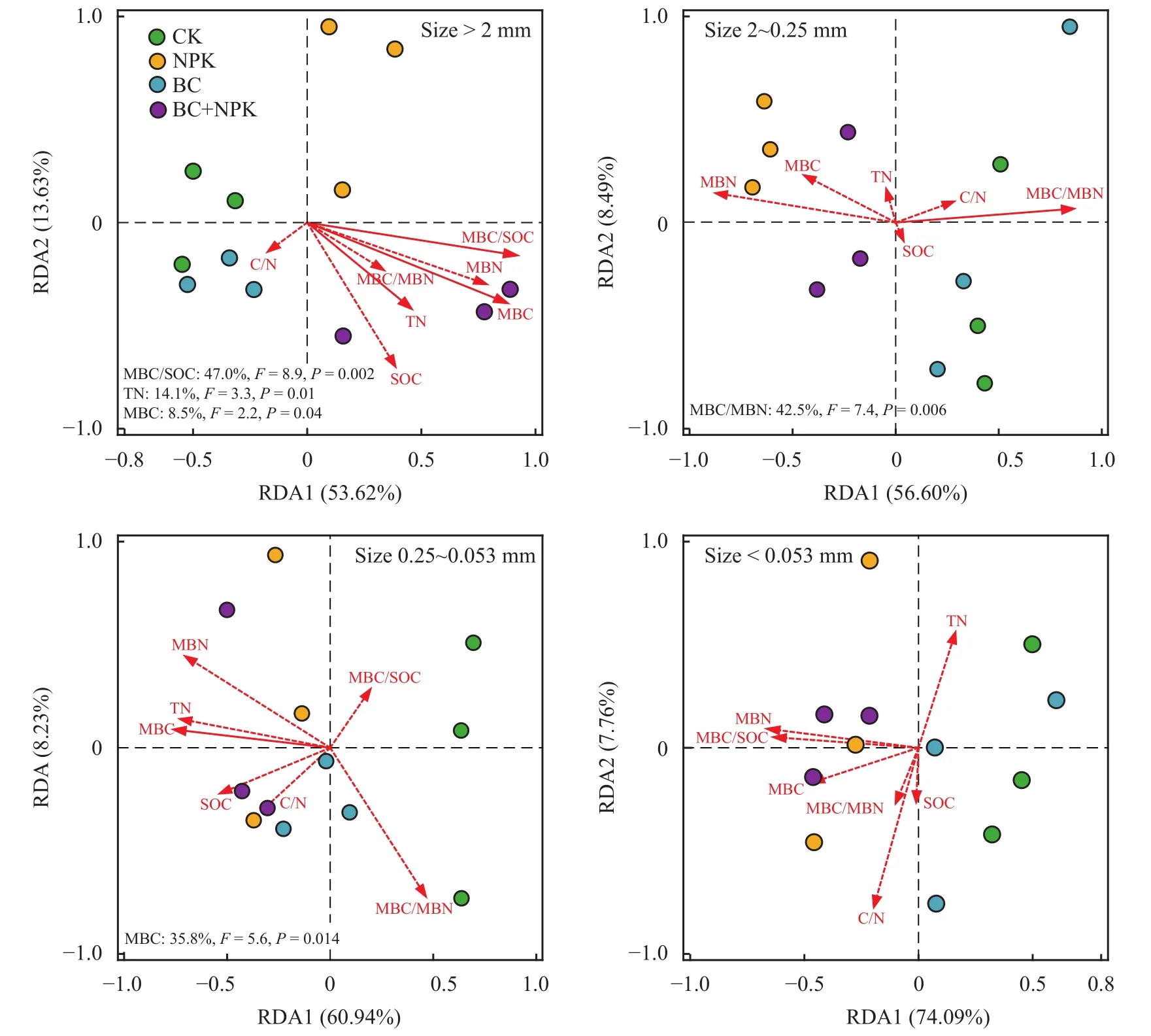
图 4 各粒径团聚体中酶活性与环境因子的相关性Fig.4 Correlation between enzyme activities and environmental factors of each particle-size fraction
3 讨论
3.1 生物炭配施化肥对潮土耕层土壤养分含量的影响
施肥是土壤养分的重要来源。诸多研究表明,生物炭作为一种富含碳的有机物料在培肥地力方面作用显著[21-24]。本研究结果显示,生物炭对土壤有机碳和总氮含量影响显著,而对铵态氮和硝态氮含量没有显著影响,其中单施生物炭或生物炭与化肥配施均显著增加了土壤有机碳和总氮含量,这与前人报道的结果[21, 25-26]较吻合。这主要归功于生物炭自身携带一定的碳、氮源,且施入土壤后不易分解、相对稳定[22, 24]。本试验中,连续2年施用化肥后潮土有机碳和总氮含量变化不大,这与刘长安[27]在有机质极低的土壤上的试验结果相一致;然而,Zhu等[28]在果园土壤上的研究发现,单施化肥显著增加了土壤有机碳和总氮含量,这些差异性结果可能与土壤类型、土壤肥力、肥料用量等有关。本研究发现,化肥和生物炭对土壤速效钾含量均有显著影响,与前人的研究结果[29]相一致,并发现生物炭施入土壤后可显著增加土壤速效钾含量,这可能是由于生物炭中的灰分携带了大量的钾素,且易于释放到土壤中。本研究还发现,土壤有效磷含量的变化主要是由施用化肥引起的,而添加生物炭的影响较小,其可能原因:一方面是本试验所选用的生物炭裂解温度较高,易生成磷钙矿,导致其有效磷含量较低[30],另一方面是生物炭中检测到的全磷含量也较低,导致其施入土壤后释放到土壤中的磷素有限。
3.2 生物炭配施化肥对潮土团聚体微生物量碳、氮含量及微生物熵的影响
土壤团聚体中微生物量碳、氮的分布特征可以在一定程度上表征土壤质量的变化。本研究发现,MBC、MBN 含量在大粒径团聚体 (> 0.25 mm) 中的含量高于在微团聚体 (0.25~0.053 mm) 和粉粘粒(< 0.053 mm) 中的含量,与前人研究结果[31-32]一致。这是因为大团聚体相较于微团聚体和粉粘粒能够保持更高的营养水平,有利于微生物的定殖[33]。本研究还发现,粒径2~0.25 mm团聚体中的微生物量碳、氮要高于粒径 > 2 mm团聚体,这可能是由于本试验中用的潮土为砂壤土,有机质含量偏低,且粒径 > 2 mm团聚体孔隙较大,碳循环周转速率过快,难以保证足够的养分来维持微生物的生长繁殖,这表明在潮土中粒径2~0.25 mm团聚体是微生物分解利用有机碳最有利的微环境。
施肥对不同粒径团聚体中的MBC、MBN含量影响各异,白文娟等[32]的研究表明,不同有机肥施用均可显著提高各粒级团聚体的MBC、MBN含量,其中对粒径 > 2 mm团聚体中微生物量增幅较大。本研究结果显示,生物炭和化肥配施相较于不施肥显著增加了粒径 > 2 mm团聚体MBC、MBN含量,可能原因是生物炭配施化肥有利于该粒径团聚体中有机碳及氮的储存,为微生物生长繁殖提供了良好的微环境[34]。本研究还发现,在粒径2~0.25 mm团聚体中,所有施用化肥的处理 (NPK、BC+NPK) 较不施肥均显著增加MBN含量,而单施生物炭则降低MBN含量。这可能是因为作物和微生物对土壤氮素有竞争作用[35],施用化肥促进了该粒径团聚体中含氮量较高的微生物生长,而单施生物炭显著增加该粒径团聚体中的C/N值,不利于微生物利用氮源。
此外,微生物熵 (MBC/SOC) 是指微生物量碳占土壤总有机碳的比率,能有效指示有机物质施入土壤后微生物量碳的转化效率和土壤碳损失等[36]。土壤中微生物熵一般为1%~4%[37],其值越大,土壤微生物对总有机碳的利用率越高。已有研究表明,生物炭中能够被微生物利用的成分很少,因此其施用会降低土壤微生物熵[38]。本研究发现,与不施肥相比,施用生物炭处理 (BC、BC+NPK) 均显著降低了2~0.25 mm和0.25~0.053 mm 团聚体中的微生物熵,但显著增加了粒径 > 2 mm团聚体的微生物熵。这可能是因为生物炭施入土壤后其少量的易分解有机碳成分首先在粒径 > 2 mm团聚体中被微生物利用。随着不同粒径团聚体间的相互转化[39],其惰性碳成分主要进入到粒径2~0.25 mm和0.25~0.053 mm团聚体中。这表明生物炭的施用有利于2~0.25 mm和0.25~0.053 mm两个粒径团聚体对有机碳的固存。
3.3 生物炭和化肥对潮土团聚体中胞外酶活性的影响
土壤团聚体通过控制不同粒级中有机质的可接触性、微生物群落组成、水分等因素直接影响土壤酶的活性[40-41]。本研究结果显示,在大团聚体 (粒径> 0.25 mm) 中的各酶活性均显著高于微团聚体和粉粘粒中的,最高酶活性在粒径2~0.25 mm团聚体中。Bach等[42]研究结果也表明,湿筛分离的大团聚体 (粒径 > 1 mm) 具有更高的潜在酶活性,尤其是与碳循环相关的纤维二糖水解酶和β-葡萄糖苷酶,这是因为土壤酶活性主要来源于土壤中微生物的代谢活动,而分布在大团聚体中的微生物比微团聚体中的更丰富[43],因此多种酶活性均在大团聚体中较高。本研究中,粒径2~0.25 mm团聚体作为土壤酶活的主要载体,其形成与稳定对于促进土壤有机碳及养分循环具有重要意义。
本研究结果表明,粒径 > 0.25 mm团聚体中的β-葡糖苷酶、β-纤维二糖苷酶、α-葡糖苷酶和β-木糖苷酶活性在单施生物炭和不施肥处理之间差异不显著。这与Tian等[35]在整体土壤上的研究结果相近,其结果显示单施生物炭对土壤β-葡糖苷酶和β-木糖苷酶活性无明显影响,并对β-纤维二糖苷酶活性有抑制作用。这主要归因于生物炭的高度芳香化结构,不易被微生物分解利用[44];另外,生物炭对酶分子的吸附作用可能掩盖了酶活性位点,从而降低酶活性[45]。然而本研究发现,生物炭配施化肥相较于不施肥显著提高了粒径 > 2 mm团聚体中的上述四种酶活性,这主要归因于化肥和生物炭具有协同增效作用[35],有利于该粒级团聚体中的微生物利用碳源,并减弱生物炭对酶反应结合位点的吸附阻隔,但其深层机理还需要进一步研究。而施用化肥 (NPK、BC+NPK) 可显著提高粒径 2~0.25 mm和0.25~0.053 mm团聚体中的上述四种酶活性,并提高了乙酰氨基葡糖苷酶活性,可能是由于这两个粒径团聚体对土壤有机质有较好的保护作用,化肥施入后激发了碳循环过程[46]。
4 结论
在田间试验条件下,生物炭配施化肥可显著提高耕层土壤养分含量,粒径 > 2 mm和0.25~0.053 mm团聚体中的MBC含量,以及耕层土壤、粒径 >2 mm、2~0.25 mm团聚体中的MBN含量;显著提高粒径 > 2 mm、2~0.25 mm和0.25~0.053 mm 团聚体中的碳循环相关酶活性,并显著降低耕层土壤以及粒径2~0.25 mm和0.25~0.053 mm团聚体中的微生物熵。冗余分析显示,团聚体中的微生物量碳氮(MBC、MBN、MBC/MBN) 是酶活性变化的主导因子,其中的MBC含量对团聚体酶活性变化的贡献最大。总体来看,生物炭与化肥配施有利于土壤碳的固存,改善土壤微环境,提升土壤质量,且生物炭添加到土壤中有较长的后效作用。
猜你喜欢
腐植酸(2021年2期)2021-12-04
吉林蔬菜(2021年2期)2021-07-19
云南农业(2021年5期)2021-06-19
作物学报(2020年5期)2020-04-16
福建茶叶(2019年12期)2020-01-19
农业机械学报(2019年12期)2019-12-31
中成药(2018年5期)2018-06-06
中成药(2017年8期)2017-11-22
天然产物研究与开发(2014年3期)2014-04-27
新疆农垦科技(2014年2期)2014-02-28
