
土壤紧实胁迫下根系–土壤的相互作用
2021-04-15 09:23:46张方博侯玉雪敖园园申建波金可默
植物营养与肥料学报 2021年3期
张方博,侯玉雪,敖园园,申建波,金可默,2*
(1 中国农业大学资源与环境学院/植物–土壤相互作用教育部重点实验室,北京 100193;2 中国农业大学国家农业绿色发展研究院,北京 100193)
土壤是植物赖以生存的载体,稳定的土壤物理、化学和生物环境是植物发挥生态功能的重要前提。据估计,全球33%的土壤由于侵蚀[1]、盐碱化、酸化、污染或压实作用而出现中度或严重退化,其中52%的农业用地受到了严重的土壤退化影响[2]。据预测,未来25年内土壤退化可能会使全球粮食生产率降低12%,粮食价格上涨30%[2]。其中,土壤紧实、土壤侵蚀和有机质下降并称为农业生产的三大威胁[3],是阻碍粮食增产的世界性难题。
土壤紧实胁迫无处不在,研究发现即使湿度较高的土壤也可能变得非常紧实,抑制根系的生长进而影响作物产量的提高[4]。近年来,随着农业现代化进程的加快,我国土壤紧实胁迫的问题日益严峻。据2016年《全国耕地质量监测报告》统计数据显示,我国耕地土壤结构性问题比较明显,其中耕层变浅、土壤板结问题较为突出。依据适宜作物生长的土壤容重为1.10~1.35 g/cm3的标准评价,全国有25.9%的监测点土壤容重大于1.35 g/cm3。土壤容重超过适宜作物生长范围后,即使是很微小的增加都会对植物根系生长产生较大的抑制作用。研究表明,当土壤容重达1.40 g/cm3时,豌豆根系的生长速率仅为生长在土壤容重为0.85 g/cm3上的65%[5]。土壤容重与土壤紧实度呈线性正相关,与产量呈线性负相关,土壤容重每增加0.1个单位,玉米减产10%~20%[6-7]。但是,土壤紧实胁迫这个重要的产量限制因素被长期忽视[8]。目前,缓解土壤紧实胁迫的措施主要包括深耕、施用有机肥、秸秆还田和施用土壤调理剂等[9],同时,合理的水肥管理和抗性品种的选育也很重要。然而,从长远角度来看,这些土壤改良措施一方面会加速土壤有机质的分解,破坏土壤团聚体结构,进而削弱土壤结构的稳定性;另一方面也会消耗大量的时间和人力物力,所以上述方式并非缓解土壤紧实压力的最佳选择[10-11]。
根系是植物重要的吸收、合成、固定和支持器官,同时又是土壤水分变化的直接感应者[12]。根系非常“聪明”,可以感知复杂的土壤环境因子,并做出适应性反应,具有较高的可塑性。根系在适应外部土壤环境和做出可塑性响应的同时,也会对土壤结构产生影响[11],根系对土壤物理结构的影响包括:1) 影响土壤养分和水分的分布[13];2) 影响根土界面的土壤微结构[14];3) 形成根系生长通道 (衰老、死亡的根系所形成的通道),影响土壤的通气性[15];4) 根分泌物的化学作用会影响土壤团聚体的分布、结构和稳定性,也会影响根土界面的土壤水分情况[16];5) 根分泌物还会对土壤有机碳库和微生物群落产生影响[17]。可见,挖掘和强化根系自身的生物学潜力,提高根系在紧实土壤中的适应性,对于最大限度地保证其在紧实胁迫下的正常生长非常关键,作为应对土壤紧实胁迫的有效策略,具有重要的现实意义[11,18]。
目前,农田管理不当和农业机械压实引起的土壤紧实问题,是农业绿色可持续发展和作物绿色高产高效的重要限制因素之一,也是导致土壤环境恶化以及土壤退化的主要因素。因此,为了全面思考和解决土壤紧实问题,必须将其放在与地下土壤环境保护和地上生态环境保护同等重要的位置[19-20]。然而植物根系分布于土壤之中,远不如地上部性状研究直观、方便[21],生长环境的复杂性和较大的取样误差等都极大影响了根系研究的准确性和稳定性,因而人们对根系的认识总是滞后于对地上部茎叶的研究和了解,使得土壤紧实胁迫下根系与土壤的互作研究亟待开展和突破。
1 土壤紧实胁迫的成因与危害
1.1 土壤紧实胁迫的形成
目前,土壤紧实已经成为制约农业可持续发展的世界性难题[22]。据2016年联合国粮农组织 (FAO)统计数据显示,全球约有4%的土地正在遭受紧实胁迫 (http://www.fao.org/3/a-i6473e.pdf)。据估计,欧洲有3300万hm2可耕地、全世界有6800万hm2土地由于土壤紧实而发生土壤退化[23-24]。近年来,我国土壤紧实问题日益突出,其中东北黑土区、华北棕壤区以及南方红壤区土壤紧实现象较为普遍,且呈现出逐年加重趋势。多种因素会导致紧实胁迫的发生,包括土壤含水量、践踏频率、耕作措施、土壤有机质含量、降雨、轮作频率、灌溉措施等[25]。例如,水分会降低土壤膨胀和收缩的作用力,导致土壤紧实度增加,土壤动物减少[26]。Zhang等[27]的研究结果表明,2年的干旱胁迫显著降低了0—10 cm土层的饱和水电导率和总孔隙度,增加了土壤容重。华北地区是冬小麦–夏玉米连作一年两熟作物的主产区,高度集约化的农业生产,使华北平原地区面临严重的地下水资源枯竭问题[28-29]。而且该地区长期采用传统的冬小麦播前旋耕、夏玉米免耕播种的耕作模式,导致土壤耕层浅、犁底层变硬的问题日益突显[30]。此外,农业劳动力成本上升与农机购置成本下降,华北平原农业机械化快速发展,随之带来机械田间作业造成的土壤紧实问题[31]。适宜作物生长的土壤容重范围是1.10~1.35 g/cm3,而2016年《全国耕地质量监测报告》数据显示,华北地区土壤容重大于1.40 g/cm3,土壤紧实度过高的问题非常突出。目前,东北地区农业机械化率处于全国较高水平,据《黑龙江统计年鉴–2017 》显示,2016年黑龙江大中小型农业机械拥有量超过158.6万台,2015年吉林农业机械拥有量超过202.1万台,2016年辽宁大中小型农业机械拥有量超过137.6万台。因此土壤压实正逐渐成为限制东北地区粮食产量的主要因素[32]。农业机械的主要参数,包括农业机械重量、发动机振动、通行次数等,均会显著提高土壤的紧实胁迫水平,且农业机械行走时,地面压力向下传递及其传递效果取决于土壤水分状况及其剖面分布。有研究发现土壤紧实程度与农业机械重量呈正相关关系[33],与小型拖拉机相比,大豆田中中型拖拉机贯穿阻力平均增加率为9.1%~53.7%,大型拖拉机的贯穿阻力平均增加率为44.4%~58.1%[34]。
1.2 土壤紧实胁迫对土壤和作物生长的影响
农业机械的使用等所造成的土壤紧实胁迫,会严重影响土壤的物理、化学和生物环境,导致作物根系下扎困难、生长不良,严重减产,最终危害整个农业生产。土壤孔隙是土壤水分和空气的主要储存空间。土壤紧实会减少土壤孔隙,破坏孔隙网络,从而使土壤中空气和水分的输送受阻,导致土壤持水性降低,形成缺氧的微环境,严重阻碍根系的正常生长[25,35]。Haunz等[36]发现在降雨较多的年份,紧实的砂壤土使净矿化率降低33%,反硝化作用提高20%。Bhandral等[37]也发现紧实胁迫会加剧草原土壤的反硝化过程,紧实土壤中NO释放量是非紧实土壤的7倍,造成大量的氮素损失。另一方面,Li等[38]研究发现,耕层或耕层以下土壤容重的增加均会显著抑制土壤的酶 (转化酶、蛋白酶、脲酶、磷酸酶) 活性。Jordan等[39]研究表明,随着土壤紧实度的增加,土壤酸性磷酸酶活性显著下降,但碱性磷酸酶活性变化不明显。此外,Pupin等[40]的研究发现,紧实导致土壤硝化细菌丰度降低38%~41%,细菌总量显著降低22%~30%,但反硝化细菌增加49%~53%。也有研究表明,只有在土壤容重达到1.7 g/cm3以上时,土壤微生物量和碳矿化才会受到负面影响[41]。
土壤紧实可以通过影响多种形态和生理过程降低作物产量[25]。由于根系生长在地表以下,对土壤有较强的依赖性,土壤紧实度增加会影响根系活力和植株地上部生长,进而对作物的产量产生负效应[42]。根系生长的土壤环境主要呈现为一种紧实与疏松土壤的混合结构[43-44]。“层”式土壤紧实分布情况 (土壤密度的垂直不连续性) 通常会限制根系生长及其对资源的利用,机械阻力的空间异质性也会影响根系的分布情况[45]。由于土壤耕作的影响,紧实度较大的土层分布较深,通常被定义为犁底层[46-47]。植物扎根于土壤中,通过根系获取水分和营养物质,而穿透阻力决定根系在土壤中的下扎能力[48]。土壤紧实程度越高,穿透阻力越大,根系生长就要耗费更多的能量。研究发现,土壤机械阻力 (硬度) 与土壤容重大体上呈线性关系[49-50]。土壤紧实度的增加会引起玉米产量的显著降低,使用轴重 (每根车轴允许分摊的最大整车重量)分别为8和19 t的农机进行压实时,平均产量分别降低了13%和20.4%[51]。Zhang等[52]发现作物产量与小型四轮拖拉机在试验区域的作业行走的次数成反比关系,随着行走次数的增加,玉米产量从 7.43 t/hm2降低至 6.25 t/hm2,当行走次数超过3 次时,春小麦的产量从 2.5 t/hm2降低至 2.0 t/hm2。
大量研究表明,土壤紧实胁迫下不同种类植物生长所受到的抑制作用和减产程度不同[18, 34, 53-55]。相较于饲用萝卜和油菜 (直根系),紧实胁迫对黑麦 (须根系) 根系穿透土壤的抑制作用更强。在较高的土壤紧实胁迫下,油菜和饲用萝卜的根系垂直穿透数量约是黑麦根数的两倍 (15—50 cm土层内),而在没有土壤紧实胁迫的情况下,3种作物的根系垂直穿透数量没有明显差异[54]。土壤紧实胁迫下土壤容重增大,大量研究表明,超过适宜植物生长的容重范围会导致作物不同程度的减产[56-60]。研究显示,大豆根系无法穿透容重为2.0 g/cm3的土层向更深层的土壤生长[61]。土壤紧实除了影响根系的生长,还有可能通过影响作物地上部的生长抑制产量的提高。严重的土壤紧实会降低作物叶片数量、叶面积,以及地上和地下的干物质产量[62]。与其他作物相比,玉米和块根作物(如甜菜) 对土壤紧实最敏感[63],而小麦对紧实土壤相对不那么敏感[64]。Jin等[18]研究表明,土壤紧实胁迫限制根系生长,进而降低了小麦分蘖和叶片的伸长。适度的土壤压实则能够增加小麦的产量[64],然而,Ahmad等[65]提出当土壤容重在 1.6~1.7 g/cm3范围时,会对小麦的产量产生负面影响。Muhammad[66]的研究表明,机械压实导致表层土壤 (< 30 cm土壤剖面) 紧实,0—30 cm土层的土壤容重和穿透阻力显著增加,严重限制了玉米根系生长和增生,导致产量降低 (2017年产量下降7.58%~15.50%)。Canarache等[67]提出土壤容重每增加100 kg/m3,玉米产量则下降 18%。在中度紧实 (1.76 g/cm3) 和严重紧实 (1.82 g/cm3) 的土壤条件下,玉米产量分别降低了25%和50%[68]。因此,土壤紧实会大幅降低农业生产力,造成严重的经济损失。
整体而言,土壤紧实度对根系生长影响的研究结果较为一致,即在紧实土壤中根系形态发生改变,生长速度减慢。紧实土壤抑制根系下扎深度,根层变浅,根系直径增加,根系夹角发生变化[18]。Bengough等[69]研究发现增加豌豆根系的生长阻力,根系会产生胁迫反应,30 min内根伸长速度减小50%,当压力解除后,生长速度会略微增加。当土壤紧实阻力达2.5 MPa时,根系的生长完全被抑制[70]。土壤容重较大时,根系的生长范围也会受到限制(P< 0.05),根系表面积 (P< 0.001)、根系总体积 (P<0.001) 和总根长均显著降低 (P< 0.05),而根系直径显著提高 (P< 0.05)[71]。Bengough 等[49]研究发现,机械阻力的存在使得豌豆根在距根尖2 mm处直径迅速增大,然后直径保持不变,直到距离根尖10 mm左右再继续增大。土壤紧实促进根系细胞膨胀,使根系变粗[72]。根系直径增大的生理意义可能是可以提高根系对轴向阻力的克服能力。De Freitas等[73]发现,生长在紧实土壤中的玉米根系,其侧根的发生形式也会发生变化,这是由于紧实土壤中缺少大孔隙且孔隙间不连续,致使根系不得不改变其形态或结构来穿透坚硬的土层。Grzesiak等[74]比较了1.33和1.50 g/cm3(花园土、泥炭和砂的比例为 1∶1∶3) 土壤容重下黑小麦根系的生长状况,发现轴根长度、侧根数量和长度均不同程度减少。同样有研究表明,在14个冬小麦品种中,土壤压实 (土柱容重1.6 g/cm3,紧实阻力1.06 MPa) 导致小麦轴根和侧根数量减少[75]。因此,土壤紧实阻碍根系生长及其对底层土壤养分资源的利用,降低作物生产力水平[23,76]。与根系较少的植物相比,保留较多根系的植物在紧实土壤中能够表现出更好的探索能力[77]。
2 紧实土壤中根系–土壤–微生物的协同互作
目前针对土壤紧实胁迫的缓解措施以农艺措施和品种选育为主。一些农艺措施,如行间深松、旋耕翻土等极易引起土壤二次压实,且耗费大量人力及自然资源等。而品种选育不确定性强,易受基因型和环境的影响,时间长成本高。而通过深入挖掘根系生物学潜力,强化关键根系/根际性状,可以提高植物对土壤紧实胁迫的适应性 (图1),更符合农业绿色可持续发展。
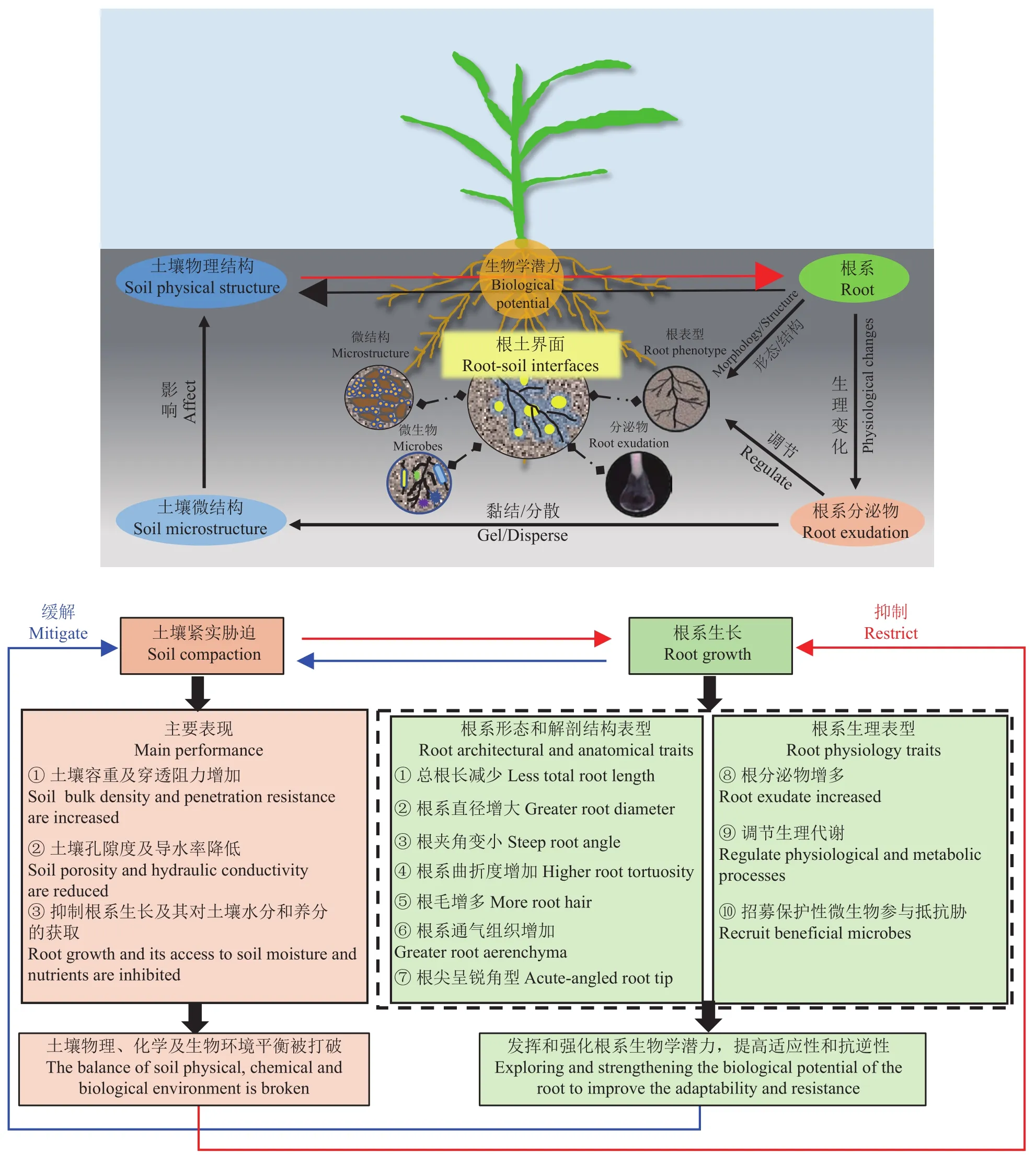
图 1 响应紧实胁迫的关键根系/根际性状Fig.1 The key root/rhizosphere traits in response to soil compaction stress
2.1 根系形态和解剖结构适应性变化
根系在漫长的进化过程中,发展出重要的适应性策略来应对外界环境胁迫[78-79]。根系生物学潜力的发挥对于保证其在胁迫环境中的正常生长至关重要。根系的伸长是分生区细胞分裂分化,并伴随着伸长区细胞体积的扩大的结果[80],当根系在紧实土壤中生长时,细胞直径及长度会发生变化。水分进入细胞后,细胞产生膨压,从而为细胞的伸长和随后根系的生长提供了原动力。当根系受到阻抗时,根系的伸长速率会下降,其中可能的原因是由于轴向的细胞伸长和形成速率受到了影响[81]。
根系在紧实土壤中穿插生长和锚定时,其自身的生物力学性质非常关键[14, 82]。根毛可以提高植物根系的抗张强度,并增加与周围土壤的接触面积,从而提高根系的固定作用[83],而这种固定作用可以促进根从疏松土层向较紧实的土层生长[50,82]。Haling等[84]发现,与无根毛大麦突变体相比,有根毛的大麦基因型其根系对紧实土层 (1.6~1.7 g/cm3) 的穿透能力更强。根系的拉伸强度 (tension strength) 和根部内聚力 (root cohesion) 是重要的根系生物力学特征[10,85]。其中,根系内聚力可以通过根面积比和拉伸强度计算得出[85]。土壤紧实度诱发根系力学性质的变化主要表现为直径变粗以降低根系在紧实土壤中生长时所受到的轴向作用力[18-19],避免根系受到阻抗时发生弯曲。近年来大量研究发现,紧实土壤中根系解剖结构发生的变化,对根系生物力学性质的影响更加直接[10,86]。当根皮层细胞尺寸较小、排列较紧密时,根系组织密度提高,根系的硬度和张力增强,根系在紧实土壤中的抗弯曲能力提高[10]。研究发现与非紧实土壤相比,紧实土壤条件下黑小麦根系通气组织所占比例增加了20%,大豆根系通气组织所占比例增加了7%,通气组织比例的增加显著促进了根系在紧实土壤中的生长[87]。此外,Colombi等[88]发现土壤紧实度增加时,小麦胚根通气组织所占比例增加的同时,皮层细胞直径也显著增加。根冠处于根系的最前端,当根系在紧实土壤中穿插生长时,作为机械感受部位首先感知土壤的紧实压力[89-90],并传递胁迫信号引导根系趋利避害。Iijima等[91]发现,在紧实土壤条件下,去除玉米根冠会增加根系的生长阻力,降低根系伸长速率。最近,研究发现根冠的形状也会影响紧实土壤中根系的生物力学性质,与具有钝角形状根冠的小麦相比,具有锐角形状根冠的小麦在紧实土壤中根系的生物力学性质显著提高,根系的生长速度加快[89]。但是Roué等[90]在拟南芥的研究中发现,紧实土壤中具有锐角形状根冠的突变体fez-2根系穿透能力较弱,而具有钝角形状根冠的突变体smb-3则表现出较强的穿透能力,推测其主要原因是由于突变体fez-2根冠的细胞层数较少,导致根冠硬度发生变化,对紧实胁迫的机械感知敏感度大大降低。可见,根系生物力学性质直接决定了根系能否在紧实土壤中正常生长[10-11, 19, 86]。
根系不发生弯曲变形会使根系很难利用土壤中的缝隙和土壤强度相对较低的区域。但是,如果根系太容易发生变形,又不利于根系穿透坚实的土层进入到土壤强度较弱的深层土壤。根系贯穿坚硬土层时会受到不同强度土壤空间分布的影响,这种土壤强度的变化通常发生在耕作土层和底土的交界面上。根系发生弯曲时受到的土壤机械阻力与土壤水分状况有关[92]。具有深根系构型的植物可能更适于在水分胁迫的环境中生长[93-94]。深根系构型与植物的扎根深度有关,如前所述,根系在坚硬土层上的入射角度对根系在土壤中的穿透作用有重要的影响。具有近乎垂直角度的根系在利于穿透坚硬的土层同时也具有较深的扎根深度。小麦根系穿透紧实土壤时的基因型差异仅在接近垂直角度的根系研究中有所发现[95]。深入理解紧实土壤中根系的适应性生长和机制,对于挖掘根系自身生物学潜力保证作物生产力具有重要意义[19, 96]。
2.2 紧实胁迫下根系对土壤孔隙的利用
土壤紧实胁迫下,土壤孔隙网络遭到破坏,连通性降低,曲折度 (tortuosity) 增加,限制了养分向根表的扩散,进一步降低了养分在土壤孔隙中的移动性,使得土壤养分无法被根系吸收利用[97]。在紧实土壤中,由于土壤的干湿交替等造成的裂隙,蚯蚓的生长穿梭以及前茬作物生长留有的根系通道等,也会在土壤中形成很多机械阻力较小的区域[19, 98]。在此情形下,当根系遇到紧实度较高的土层或者土块时,通常会倾向于向土壤分布较疏松的位点进行生长。在自然土壤中,裂隙、生物孔隙、土壤孔洞以及团聚体间的孔隙等组成了复杂的土壤孔隙网络[15]。与周围土体相比,生物孔隙中生长阻力小,养分含量高,微生物活性高[99-100]。因此生物孔隙能够储存大量的水分和养分,有利于根系对深层土壤资源的吸收和利用[101]。根系非常聪明,可以“感知”或“寻找”土壤裂纹、孔洞或较大的土壤孔隙。
根系生物力学性质除了保证根系有足够的“力量”在紧实土壤中穿插生长外,也会帮助根系去寻找土壤中阻力较小的土壤孔隙,通过提高对土壤孔隙的利用最大限度地拓展根系的生长空间[11,19,86]。研究发现,随着土壤紧实度增加,根系趋向阻力较小的孔隙而生长的现象非常明显[45,102]。Pfeifer等[103]研究紧实土壤中大麦根系对人造孔隙的响应,发现孔隙的存在促进了根系的生长和地上生物量形成,地上部干重增加了20%。最近,有研究表明不同作物根系利用土壤孔隙的方式也不同。Athmann等[104]采用内窥镜的方法发现小麦根系在土壤孔隙中有很多侧根与孔隙壁接触,蚕豆根系穿过土壤孔隙,油菜根系在土壤孔隙中垂直生长并有大量的根毛。近年来,随着X-ray CT技术在土壤科学中的应用,为研究根系和土壤之间的关系提供了有利的技术手段。Colombi等[88]在田间和室内试验条件下,采用X-ray CT方法研究紧实、非紧实土壤和存在孔隙的紧实土壤中大豆、小麦和玉米根系的生长方式,发现在紧实土壤中玉米根系主要沿着孔隙生长,大豆根系穿过孔隙和沿着孔隙生长的比例大致相同。小麦根系则在田间和室内试验中表现出不同的结果:田间条件下,小麦根系主要是沿着孔隙生长;室内试验条件下主要是穿过孔隙生长。这可能是由于在田间条件下,小麦的侧根水平生长使得CT扫描无法观察到根系。Atkinson等[105]在室内条件下研究非紧实和紧实土壤中不同基因型的小麦根系在孔隙中的生长方式,发现小麦根系在紧实土壤中主要沿着孔隙生长,而在非紧实土壤条件下,根系以穿过孔隙生长为主,这个结果与Colombi等[88]研究结果相反。Atkinson等[105]从穿透力的角度解释了结果的不同,Colombi等[88]研究中土壤的穿透阻力为1 MPa,穿透阻力太小不足以限制根系的生长。上述研究可知,利用X-ray CT这一重要的研究方法和手段,可为研究根系在孔隙的生长方式,明确根系与土壤孔隙之间的关系提供重要的技术支撑。此外,根系也会通过自身的生长,“重置”土壤颗粒的分布改变土壤孔隙结构,使得小孔隙扩大为较大的孔隙。Gao等[45]研究较深的根系分布时发现,在自然土壤中,只有当深层土壤存在孔隙时,根系才可以扎根其处。土壤耕作以及水分和养分的运移造成了土壤孔隙的异质性[106]。近年来,为了更好的揭示土壤结构的复杂性,利用微型拓扑图片对土壤进行结构分析成为该领域的研究热点[107]。Kravchenko等[108]发现,在土壤免耕和传统耕作下的土壤中可表征大孔隙的三维像素大多呈现异质性分布。Bodner等[109]研究土壤干湿交替循环对土壤孔隙的大小和分布的影响,发现干湿交替在提高土壤孔隙度的同时降低土壤孔隙的异质性。
2.3 根分泌物对根–土界面微结构的影响
在植物根系对胁迫环境的适应性生长过程中除了形态适应,还有生理适应。一个重要的体现就是根分泌物的变化。植物代谢产物总量可达20万到100万种[110]。土壤是孔隙空间和土壤团聚体的高度复杂系统[111]。土壤团聚体是与土壤微生物生存关系最密切的微环境。不同粒级的土壤团聚体由于其形成过程不同,其内部孔隙的数量、大小、分布和连通性不同,从而导致不同土壤团聚体中的水分、养分等物理化学性质存在较大差异。土壤物理结构,特别根际土壤微结构,不断地被植物根系分泌物和微生物代谢物的释放所改变[112]。
越来越多的研究表明,根系分泌物中存在的某些化合物能够通过分散和胶结土壤,参与根际过程,影响根–土界面的土壤结构 (土壤微结构)[113]。植物根系在根际释放不同的有机物质,对土壤结构可能产生不同的影响。有研究表明,根系分泌物对土壤微团聚体的大小、分布、稳定性、吸附性能及亲水性等物理性质有显著影响[114]。Traoré等[115]将葡萄糖、聚半乳糖醛酸、模拟分泌物和收集的根分泌物(总碳量相同)分别添加到等量土壤中,在25℃下孵育30天,发现添加的所有底物都增加了团聚体的稳定性,但是持续作用的时间取决于添加物的化学性质。与对照组相比,培养30天后水稳性团聚体比例分别增加了 3.8倍 (根分泌物)、4.2倍 (模拟分泌物)、2.5 倍 (聚半乳糖醛酸)和2 倍 (葡萄糖)。Akhtar等[116]使用一种基于硝化纤维的粘附力的方法,测定了土壤中可能存在的20多种多糖的粘附能力。通过对硝化纤维素片的扫描,定量测定了粘附在不同剂量的特定多糖斑点上的土壤重量。其中黄蓍胶、凝胶多糖、黄原胶和壳聚糖等表现出较强的粘附性,且黏附土壤重量随着多糖添加量的增加而增大。水培法收集奇亚籽、玉米和大麦根系分泌物,通过气相色谱-质谱联用 (GC-MS) 技术分析发现大麦中有机酸含量最高,而奇亚籽中糖类 (多糖或游离多糖) 含量最高,玉米居于二者之间。将收集到的分泌物添加到土壤中孵育一段时间,结果显示大麦根系分泌物趋向于分散土壤,玉米和奇亚籽根系分泌物趋向于胶结土壤,在根或种子周围形成更稳定的土壤结构[113]。
Galloway等[117]在谷类植物根系分泌物中发现结构高度复杂的多糖,具有与土壤结合的特性,对谷类植物根鞘的形成和稳定有很大的贡献,有利于土壤结构的稳定。有机酸阴离子易被土壤矿物颗粒吸附,增加粘土的净负电荷,增强了黏土的分散性[118]。Szatanik-Kloc等[119]发现,土壤紧实胁迫时,禾本科作物如小麦、大麦和玉米等根细胞中酸性基团的合成和流出减少,根表面负电荷和强酸性基团显著下降。相比于非紧实处理,在紧实基质中培养的玉米根系向根际释放的碳增加了2.5倍[120],这些碳主要以粘液的形式释放到根际中[121]。Somasundaram等[122]发现土壤压实导致土壤中根际沉积物增多,有助于减少根系在穿透土壤过程中的摩擦阻力。同样,Oleghe等[123]发现从奇亚籽涂层中提取的植物分泌物可以极大地改善根系生长的机械条件,随着分泌物浓度的增加,砂壤土和粘壤土的渗透阻力均有所降低。除了减小根部生长的机械阻力外,分泌物的释放还增强了土壤的回弹性。
随着X-ray CT技术的广泛应用,根系生长对根际微结构的影响成为根际研究领域新的突破口和创新点[105, 124]。Helliwell等[125]在小麦、番茄和豌豆的研究中发现,虽然根系对根际土壤产生挤压作用,但是 3 种作物在非紧实 (1.2 g/cm3) 和紧实胁迫 (1.5 g/cm3) 的环境下,均显著提高了根际土壤的孔隙度,增加比例的平均值分别为47.3%和26.8%。同时,小麦、番茄和豌豆所影响的根际土壤范围也不同,分别为7077、3940和3483 mm3,其原因可能是作物间根分泌物的差异所造成。有研究发现,根分泌物显著增加了根际团聚体的稳定性,而且这种稳定性的提高独立于土壤微生物的活性及作用,因为根分泌物释放到土壤后,立即对团聚体产生了作用[126-127]。Naveed等[113]通过模型计算,发现玉米根分泌物以糖类为主,对根际土壤有粘结的作用,油菜根分泌物以有机酸为主,对根际土壤有疏散作用。最近,Benard等[128]发现,根分泌的粘胶物质提高了土壤颗粒间的胶结程度,增强了介质的连续性。
3 展望
土壤环境如干旱、养分缺乏和土壤紧实等对作物根系的生长发育具有较大的抑制作用。植物根系的发育受土壤物理特性的高度调控,特别是受土壤机械阻力的调控[129]。根系构型决定着作物对水分和养分的吸收效率,被认为是保证粮食安全的“二次绿色革命”[130]。根系可以通过形态、生理等表型来适应土壤胁迫环境,但是,目前关于土壤紧实胁迫下根分泌物–土壤物理结构–微生物的交叉互作研究较少。在土壤紧实胁迫下,充分发挥根系的可塑性以实现更好的根系–土壤的相互作用,有助于进一步推动未来粮食产量的提高。
在非生物胁迫环境下,植物会改变根分泌物响应胁迫,会将光和固定碳的5%~21%以可溶性糖、氨基酸或次生代谢物等形式释放到根际[131-133]。这些根分泌物作为根系和根际微生物之间的信使[134],介导了土壤中植物与微生物的相互作用。研究发现细菌对植物分泌的不同化学分子具有积极的趋化性,包括糖、氨基酸、各种二羧酸如琥珀酸、苹果酸和富马酸,以及芳香化合物如莽草酸、奎宁酸、原儿茶酚酸、香草酸、丙酮环酮、没食子酸、儿茶酚和木犀草素[135]。此外,根分泌物也可以作为吸引菌株在根际定殖的信号分子,例如玉米[136]、西红柿[137]和谷子[138]根分泌物中的糖类、二羧酸盐和L-氨基酸分别吸引了BacillusvelezensisS3-1、Azospirillumbrasilense和PseudomonasfluorescensPf0-1在根际的定殖。Huang 等[139]发现拟南芥生长发育、环境适应过程中产生的一系列三萜类化合物,能够指导其特定微生物群落的组成。根分泌物驱动的这种根际对话对于提高植物养分吸收、抵抗生物和非生物的胁迫非常关键。因此,紧实胁迫下根分泌物与根际微生物的对话机制研究亟待深入开展。
此外,土壤物理条件,特别是根际土壤,也不断被植物根分泌物和微生物释放的代谢物所改变[140-141]。但是,根分泌物能否通过改变根际土壤物理性质,影响根际微生物组成的研究较少。随着现代显微技术的快速发展,为深入观察根–土界面微结构的变化提供了重要手段。土壤在结构和资源可利用性方面表现出很大的时空变化,土壤微生物群落存在并活跃在高度异质的孔隙空间和土壤颗粒表面,可以形成各种生态位或微生境。这些微生境的物理化学性质的差异形成了不同的微生物生态位[142-143]。土壤孔隙的空间几何特征,如孔隙体积、形状、连通性、大小和通道的曲折度,也会对土壤微生物的分布和活性产生影响[144]。Seaton等[145]发现土壤质地的异质性对细菌多样性具有积极影响,但对真菌多样性影响很小。而且土壤质地成分 (砂粒、粉粒、粘粒) 会影响细菌和真菌的组成,许多特定的微生物类群与粉粒和粘粒共存。Juyal等[146]比较了不同土壤孔隙结构对荧光假单胞菌和枯草芽孢杆菌生长扩散的影响,结果表明假单胞菌的细胞数不受团聚体大小的影响,但是相较于2~4 mm粒径团聚体,在1~2 mm粒径团聚体中枯草芽孢杆菌的细胞数比较高。Kravchenko 等[147]结合 X 射线微断层扫描 (X-ray CT)和微尺度酶映射的技术,发现土壤中孔径30~150 μm的孔隙中微生物酶活性更高,这种孔隙丰富的土壤碳储存能力更强。上述研究表明,土壤微结构会影响土壤微生物的群落分布、活性及功能表达。紧实胁迫下,植物根系分泌物如何重塑土壤微结构,进而影响微生物群落尚不清楚,这为深入理解植物根分泌物与微生物的互作提供了新思路。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
四川劳动保障(2021年3期)2021-06-09 07:08:56
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
现代园艺(2017年13期)2018-01-19 02:28:05
中国蔬菜(2016年8期)2017-01-15 14:23:38
西南农业学报(2016年5期)2016-05-17 05:42:36
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
华南农业大学学报(2015年5期)2015-12-04 03:04:38
家庭医药·快乐养生(2014年5期)2014-06-18 14:02:16
农村百事通·健康一点通(2011年11期)2011-06-04 09:38:14
