
Origanum vulgare L. leaf extract alleviates finasteride-induced oxidative stress in mouse liver and kidney
2021-04-11 01:39QiFengSunShiXiangChenZhangFengTangXiaoYiSongFaJingHaoTianWuZhongYangDingAttallaElkottDiaaMassoudHebaKhalifa
Qi-Feng Sun, Shi-Xiang Chen, Zhang-Feng Tang, Xiao-Yi Song, Fa Jing, Hao-Tian Wu, Zhong-Yang Ding✉, Attalla El-kott, Diaa Massoud, Heba Khalifa
1Department of Hepatobiliary and Vascular Surgery, Jinan City People's Hospital, Jinan, Shandong Province, 271100, China
2Department of Hepatology, Zhejiang University Ming Zhou Hospital, Ningbo, Zhejiang Province, 315199, China
3Department of General Surgery, Wuxi Traditional Chinese Medicine Hospital Affiliated to Nanjing University of Traditional Chinese Medicine, Wuxi, Jiangsu Province, 214023, China
4Biology Department, College of Science, King Khalid University, Abha, Saudi Arabia
5Zoology Department, College of Science, Damanhour University, Damanhour, Egypt
6Biology Department, College of Science, Jouf University, P.O. Box 2014, Sakaka, Al-Jouf, Saudi Arabia
7Zoology Department, Faculty of Science, Fayoum University, Fayoum, Egypt
ABSTRACT
KEYWORDS: Kidney; Liver; Origanum vulgare L.; Inflammation; Antioxidant
1. Introduction
Finasteride (propecia) is a synthetic selective 5-α reductase type Ⅱ inhibitor that suppresses the conversion of testosterone to dihydrotestosterone[1]. Finasteride is used to treat a variety of disorders including androgenic alopecia, benign prostatic hyperplasia, and prostate cancer, but its application is limited due to its different side effects such as alternation in sexual behavior, ejaculation disorder, and liver and kidney toxicities[2]. Many studies have shown that 25-250 mg/kg/day of finasteride significantly increases the activity of liver enzymes and induces hepatocyte hyperplasia and hepatic parenchyma degeneration and fibrosis, especially in people with acute androgenic alopecia and benign prostatic hyperplasia who receive high doses of this drug[3]. Although the mechanism of hepatotoxicity of finasteride is unknown, it seems that the azasteroids metabolized by cytochrome P450 (CYP 3A4) are likely to interfere with the activity of hepatocytes and Ito cells and eventually alter the liver physiological activity[4]. Besides, due to the renal excretion of 36%-42% of finasteride metabolites, damage to the glomerular basement membrane and the podocytes and epithelial cells of proximal convoluted tubules is expectable. Also, it has been shown that low doses (e.g. 5 mg/kg) of finasteride can induce IL-6 production, renal tubular epithelial cell apoptosis, and thickening of the collagen surrounding the tubules, eventually causing renal lymphocytic infiltration, tubular fibrosis, and glomerulosclerosis[5]. Moreover, finasteride promotes the formation of tubular casts and calcium oxalate (CaOx) kidney stones through androgenic receptor-dependent signaling pathways[6]. Various studies have also shown that finasteride reduces renal physiological activity and ultimately leads to chronic kidney disease through triggering inflammatory pathways and interfering with androgenic receptorregulated renal hormonal homeostasis and hemodynamics[7].
Origanum vulgare L. (O. vulgare) belongs to the Lamiaceae family and grows in Asia and Europe. This plant has always been used in traditional medicine due to the presence of important flavonoid compounds such as apigenin, kaempferol, myricetin, quercetin, naringenin, and rosmarinic acid and its antioxidant, antiinflammatory, anti-cancer, anti-depressant, anti-viral, and antimicrobial properties[8]. Therefore, the present study was conducted to investigate the protective effects of O. vulgare hydroalcoholic extract against finasteride-induced hepatic and renal toxicities in mice.
2. Materials and methods
2.1. Preparation of O. vulgare extract
O. vulgare leaves were gathered from the rivers of Shaanxi province, China, identified by a botanist (voucher NO: XBGH710061), and dried in dark at 40 ℃. After grinding, the dried leaves were poured into 70% ethanol, mixed gently, and then the extract solution was passed through a filter paper (Whatman no. 42, Millipore, USA. Cat no. 1442125), condensed by a rotary evaporator (Buchi RE111) at 50 ℃, and finally dried at room temperature for 72 h. The final dried O. vulgare extract (35 g) was stored at 4 ℃ until use[9].
2.2. Liquid chromatography-electrospray ionization-tandem mass spectrometry (LC-ESI/MS) analysis
The Agilent G6410 Triple Quadrapole Mass spectrometer (Agilent, Waldbronn, Germany) equipped with the Agilent zorbax SB-C18 (15 cm, 3.5 μm) column coupled with a HCTultra ion trap MS detector was used to analyze polyphenolic compounds in the plant. The LC condition was set as follows: mobile phase A: acetonitrile 0.1% (v/v) + formic acid 0.1% (v/v), and B: HO + formic acid 0.1% (v/v). Linear gradients included 0%, 5%, 95% (repeated twice), and 100% for solution A and 0%, 95%, 5% (repeated twice), and 100% for solution B with applying 0.3 mL/min washing time for 75 min. The injection volume was set as 20 μL with a flow rate of 0.2 mL/min in negative ion mode (ESI) using an ESI ion source electrospray (Bruker Daltonik GmbH, Bremen, Germany). Also, gain and probe voltage of LC detection were 1.8 kV and 3.5 kV, respectively. The nebulizer gas was N(grade 5) at the 1.2 mL/min flow rate, 285 ℃ block temperature, and 30 psi Nnebulizing pressure. To identify the extract compounds, quality control samples were prepared by diluting separate analytic stock solutions using the same volume of a known internal standard[9].
2.3. Experimental design, animals, and treatment groups
Thirty BALB/c male mice [7 to 8-week-old, weighing (30 ± 4) g] were obtained from the Animal House Section of People’s Hospital, Jinan City, Shandong Province, China. The animals were kept in polypropylene cages (6 animals per cage) and maintained under 12:12 h light: dark cycles in a well-ventilated room at (25±2) ℃ and (50±5)% relative humidity. The animals were fed with water and solid standard pellets for mice ad libitum in accordance with internationally accepted guidelines for use and care of laboratory animals (EEC Directive of 1986; 86/09/EEC; National Institutes of Health publication 85-23, revised 1985). After acclimation, the mice were randomly divided into five groups (n = 6 per group) as follows: Group 1: Normal (vehicle control) group, receiving 0.5 mL/day distilled water by gavage; Group 2: Model group, receiving 25 mg/kg/day of finasteride in 0.5 mL distilled water by gavage; Group 3-5: Treatment groups, receiving 100, 200 and 400 mg/kg/day of O. vulgare extract in 0.5 mL distilled water[10], respectively, and 25 mg/kg/day of finasteride in 0.5 mL distilled water by gavage[11].
There were no specific criteria for selection of mice in this study, and the animals that received finasteride and were eligible to continue the study were randomly assigned to the groups 2-5. Treatments continued daily for 35 d.
2.4. Acute toxicity test (LD50)
The Lork’s two-step method was used to determine the acute toxicity dose (lethal dose- LD) of O. vulgare extract in mice. In this method, three groups of mice (three animals per group) were first injected intraperitoneally with the extract doses of 100, 500, and 1 000 mg/kg. During a 24-hour period, the survival of mice and toxic effects of the extract were evaluated in each group. At the second step, three mice each received the extract doses of either 1 000, 3 000, or 4 500 mg/kg via intraperitoneal injection, and the potential toxic effects of these doses were investigated. Finally, LDof the extract was calculated using the following equation:
LD= (D× D)
Where Dis the highest dose in which no mortality occurred, and Dis the lowest dose that caused mortality[12].
2.5. Total phenolic content (TPC)
A modified Folin-Ciocalteu assessment was used to determine the TPC of O. vulgare extract by using gallic acid as a standard. In this method, gallic acid (50 μL) at concentrations of 0.024, 0.075, 0.105, 0.3, and 0.4 mg/mL was gently mixed with sodium carbonate (NaCO) 7.5% (0.8 mL), Folin-Ciocalteu’s reagent (1.0 mL), and O. vulgare ethanolic extract (50 μL). After two hours of incubation at 30 ℃, the solution optical density (OD) was read at 765 nm with a spectrophotometer (Shimadzu UV-Vis spectrophotometer, Shimadzu, Japan). Finally, the TPC of O. vulgare extract was expressed as mg of gallic acid equivalent per gram of dried plant (mg GAE/g)[13].
2.6. Total flavonoid content (TFC)
A modified Miliauskas-Venskutonis assessment was used to determine the TFC of O. vulgare extract. In this method, an alcoholic mixture of ethanol/methanol (1 to 5 ratio) was prepared and added slowly to another solution containing potassium acetate 1 M (0.1 mL), aluminum trichloride (AlCl) 0.1 M (0.1 mL), and distilled water (2.8 mL). Then 10 mg of O. vulgare ethanolic extract was added to this solution and incubated at 25 ℃ for 30 min. Finally, the obtained solution absorbance was read at 415 nm by a spectrophotometer (Shimadzu UV-Vis spectrophotometer, Shimadzu, Japan), and the TFC of O. vulgare extract was expressed as mg of rutin equivalent (RE) per 1 g of O. vulgare extract (mg RE/g)[14].
2.7. 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity
A modified Brand-Williams assay was used to determine the DPPH radical scavenging activity of O. vulgare extract. For this, 3 mL of methanol DPPHradical solution (0.06 mM) was added to 25 to 170 μL aliquots of O. vulgare extract, and the solution was incubated at 25 ℃ for 3 h. Then the OD of the resulted solution was read at 516 nm by a spectrophotometer (Shimadzu UV-Vis spectrophotometer, Shimadzu, Japan) at dark, and according to the Trolox calibration curve, the DPPH radical scavenging activity of the extract was expressed as μM Trolox equivalent per 10 g of dried extract (μmol TE/10 g plant)[15].
2.8. Ferric reducing antioxidant power (FRAP) assay
A modified Benzie-Strain assay was used to determine the antioxidant activity of O. vulgare extract. In this pH-dependent method (pH = 3.6), the yellow Fe (Ⅲ)-tripyridyltriazine complex is converted to the Fe (Ⅱ)-tripyridyltriazine complex. O. vulgare extract was dissolved in methanol to reach 1 mg/mL concentration, and 20 μL of this solution was added to 180 μL distilled water containing 2 μL FRAP solution. Then the ODs of the obtained solution and methanol (as blank) were read at 593 nm by a spectrophotometer (Shimadzu UV-Vis spectrophotometer, Shimadzu, Japan), and the results were expressed as mM Fe (Ⅱ) sulfate heptahydrate per mg of O. vulgare extract [mmol Fe (Ⅱ)/mg plant][16].
2.9. Biochemical analysis
After 24 hours of fasting, the animals were weighed and sacrificed (on day 36) by cervical dislocation using standard laboratory animal protocols. Blood samples were collected through cardiac puncture and incubated at 37 ℃ for 15 min to allow clot formation. Then serum samples were separated by centrifugation (3 000 g for 15 min) to assess the liver and kidney biochemical parameters. The liver and kidney tissues were dissected, weighed with a digital scale (AND FA 2014, China) and fixed in 10% formalin solution for histopathological evaluation. Serum levels of the liver and kidney function parameters including alkaline phosphatase (ALP) (Cat No: 102395H917), aspartate aminotransferase (AST) (Cat No: 118408H912), alanine aminotransferase (ALT) (Cat No: 119395H917), gamma-glutamyl transferase (GGT) (Cat No: 120158H917), albumin (ALB) (Cat No: 101504H917), total bilirubin (TB) (Cat No: 105395H917), and total protein (TP) (Cat No: 128395H917), as well as serum creatinine (Cr) (Cat No: 109395H917) and blood urea nitrogen (BUN) (Cat No: 129395H917) levels were measured using ELISA quantification commercial kits following the manufacturer’s protocols (Pars Azmoon, Tehran, Iran) and applying a spectrophotometer (SINCO)[17]. Also, the serum levels of sodium and potassium ions were measured by a photometric method (Jenway Flame photometer PFP)[18].
2.10. Determination of pro- and anti-inflammatory cytokines
Commercial ELISA kits (IFN-γ Cat. No.: 88-7314-88; IL-12 Cat. No.: 88-7121-88; IL-10 Cat. No.: 88-7105-22; IL-6, Cat. No.: 88-7064-88; TNF-α, Cat. No.: 88-7324-88; and IL-1β, Cat. No.: 88-7013-88) were purchased from Invitrogen; Thermo Fisher Scientific Inc. (Heidelberg, Germany) to determine serum levels of the cytokines according to the manufacturer’s standard protocols. All the experiments were performed in duplicate. Briefly, a 96-well plate was first pre-coated and incubated with primary antibodies and 10× coating buffer at 37 ℃ for 24 h. Then the plates were washed three times with phosphate buffered saline, and 5× ELISA/ELISPOT diluent was added for blocking. After 1-hour incubation at 37 ℃, the standard dilution of serum samples was prepared and added to the tubes. Then a diluted (detecting) secondary biotin-conjugated antibody was added to the tubes which afterward were incubated for 30 min at 37 ℃. Finally, the color produced by the chromogenic substrate of 3,3′,5,5′-tetramethylbenzidine was read using an ELISA reader (Biotek Instruments, USA) at 450 nm. Serum levels of IFN-γ, IL-1β, IL-6 and TNF-α (pg/mL) as well as IL-10 and IL-12 (ng/mL) were reported[19].
2.11. Evaluation of oxidative stress
2.11.1. Nitric oxide (NO) assay
The Griess colorimetric method was used to measure NO level. Briefly, 6 mg of zinc sulfate was added to 400 μL of serum samples, and the mixture was centrifuged at 12 000 g for 12 min. Then the supernatant was added to a solution containing 50 μL sulfanilamide, 50 μL N-(1-naphthyl) ethylenediamine dihydrochloride, and 100 μL vanadium chloride and incubated at 37 ℃ for 100 min. The absorption of the resulting solution was read by an ELISA reader (Biotek Instruments, USA) at 540 and 630 nm wavelengths and reported as mmol/mL[20].
2.11.2. Measurement of malondialdehyde (MDA) level, total antioxidant status (TAS), and nitric oxide synthase (NOS) and catalase (CAT) activities
Colorimetric assay kits (Randox Cat No: NX2332; Randox Laboratories Ltd., Crumlin, Antrim, United Kingdom) were used for measuring serum TAS level according to the manufacturer’s recommendations using an ELISA reader (Biotek Instruments, USA) at the wavelength of 600 nm. The results were reported as mM/L. Briefly, this method is based on the bleaching of the color produced by a peroxidase (such as hydrogen peroxidase) and the conversion of unstable 2,2’-azino-di-(3-ethylbenzthiazoline sulphonate) radicals to stable ones upon the addition of antioxidants to serum samples. The ODs of samples were finally measured at 600 nm[21].
NOS catalyzes the production of NO from L-arginine and plays an important role in neurotransmission, vascular regulation, apoptosis, and immune and inflammation responses. NOS activity kit (colorimetric) (Abcam Cat No: ab211083; Abcam, Cambridge, MA, USA) was used for measuring the activity of NOS according to the manufacturer’s instructions. Based on this colorimetric assay, the NO produced by NOS participates in a series of reactions with Griess reagents and generates a colored product. Its intensity was measured at 540 nm using an ELISA reader (Biotek Instruments, USA) and reported as U/mL[22].
ZellBio GmbH quantitative colorimetric kit (ZellBio GmbH Cat No: ZB-CAT-96A; ZellBio GmbH Laboratories Ltd, Germany) was used to determine serum CAT activity according to the manufacturer’s instructions. Hydrogen peroxide (HO) is generated in cells by various oxidative reactions, and its accumulation causes the oxidation of cellular components including cell membrane lipids and DNA, leading to apoptosis. The CAT is an enzyme that detoxifies this compound by catalyzing HOto water and oxygen. Briefly, in this two-step kit which contains HO, after the addition of a sample containing CAT, the enzyme was quenched with a quencher, and extra HOreacted with a chromogen. Finally, the OD was read at 405 nm by an ELISA reader (Biotek Instruments, USA), and serum CAT activity was expressed as U/mL[23].
Spectrophotometric method was used to measure the amount of MDA level based on thiobarbituric acid (TBA) reactivity. Briefly, 150 μL of TBA solution was added to 150 μL of 30% trichloroacetic acid (Merck) and after centrifugation of the resulting solution (3 000 rpm for 5 min), the supernatant was added to 200 μL of 1% TBA (Merck) in a boiling water bath for 30 min and incubated for 10 min on an ice-cold water bath. The absorbance of the final solution was read at 532 nm at room temperature. The extent of lipid peroxidation was expressed as MDA (nmol/mL)[24].
2.12. Histopathological investigation
The liver and kidney tissues were fixed in 10% formalin, and by routine tissue processing, 5 μm sections (with LEICA SM2010RV1.2 microtomes, Germany) were prepared from paraffin-embedded blocks and stained with either hematoxylin and eosin (H&E) for the liver or periodic acid-Schiff (PAS) for the kidney. Each section was examined at ×100 magnification using a light microscopy system (Olympus IX71 microscope, Japan). Hepatic central venules and the portal triad (portal bile duct, artery, and vein), as well as renal glomeruli, tubules (proximal, distal, Henle, and collecting), and arteries were examined[20].
2.13. Statistical analysis
Graph Pad Prism version 8.0 for Windows (Graph Pad Software, San Diego, CA, USA) was used for drawing graphs. Statistical analysis was conducted in SPSS v. 16. The normality and homogeneity of the data were determined by the Kolmogorov-Smirnov test (P > 0.05), and one-way analysis of variance (ANOVA) was utilized to compare the differences of means between groups, followed by the Tukey’s post hoc test. All results were expressed as mean ± SD, and P < 0.05 indicated statistical significance.
2.14. Ethical statement
All the protocols were performed under the supervision of the Ethics Committee of Nanjing University of Traditional Chinese Medicine (Approval number: 20190800358) and were in line with the guidelines of the Animal Ethics Committee (NIH Publication 80-23, 1996).
3. Results
3.1. LC-ESI/MS analysis
Based on m/z (molecular weights [M-H]), the retention time of standard compounds in the negative ion mode, the count (mass-tocharge, ×10) of expected polyphenolic compounds, and the peaks recorded in chromatogram (Supplementary Figure 1), a total of 25 major polyphenols (i.e. 25 peaks with an intensity of > 5×10) were identified (Table 1).
3.2. Acute toxicity test (LD50)
After monitoring the toxic effects of the O. vulgare extract on mice for 24 h, it was found that one mouse died after exposure to the dose of 4 500 mg/kg, and no death or toxic signs were observed in the mice exposed to other doses. According to the results of this study, the LDof O. vulgare extract was 3.67 g/kg, which means that the doses of less than 3 670 mg/kg would have no toxic effects on mice and could be used in experiments.
3.3. TPC, TFC, and antioxidant capacity of O. vulgare extract
The results showed that the TPC of the O. vulgare extract was (89.17 ± 2.19) mg GAE/g, and TFC was (75.16 ± 2.16) mg RE/g. The O. vulgare extract showed the antioxidant activity with (8 920.15 ± 19.13) mmol Fe(Ⅱ)/mg and (192.16 ± 4.61) μmol TE/10 g, respectively by FRAP and DPPH assays. These results indicated that the O. vulgare extract had a high antioxidant capacity, probably due to the presence of various polyphenolic compounds.
3.4. Effects of O. vulgare extract on the body, liver, and kidney weights in finasteride-treated mice
At the end of the study, it was found that finasteride significantly reduced the body weight (P=0.023) and increased the weights of the liver (P=0.006) and kidneys (P=0.026) in mice compared to the normal group. However, treatment with the O. vulgare extract restored the weights of the body and the organs in a dose-dependent manner. The group receiving 400 mg/kg of O. vulgare extract showed significant improvement in the weights of the body and the organs (Figure 1).
3.5. Effects of O. vulgare extract on biochemical parameters in finasteride-treated mice
Finasteride significantly increased the levels of all examined liver enzymes [ALT (P=0.001), AST (P=0.004), ALP (P=0.016), and GGT (P=0.008)] compared to the normal group (Figure 2). However, a dose-dependent reduction was noticed in the liver enzymes after administration of O. vulgare extract (Figure 2A).
Moreover, finasteride significantly reduced serum levels of ALB (P=0.016) and TP (P=0.006) and also increased TB (P=0.004) compared to the normal group (Figure 2B). The O. vulgare extract treatment restored serum levels of ALB, TP, and TB towards normal (Figure 2B).

Figure 1. Body weight, and liver and kidney weight in different groups. The data are expressed as mean±SD. $P < 0.05, $$P < 0.01 compared with the normal group; *P < 0.05 and **P < 0.01 compared with the finasteride group. OV: Origanum vulgare.

Table 1. Qualitative evaluation of Origanum vulgare flavonoids based on LC-ESI/MS analysis.

Figure 2. Serum activity of ALT, AST, ALP and GGT (A), and serum levels of ALB, TB and TP (B) after treatment with Origanum vulgare extract. The data are expressed as mean±SD. $P < 0.05, $$P < 0.01 compared with the normal group; *P < 0.05, **P < 0.01 compared with the finasteride group. ALP: alkaline phosphatase; AST: aspartate aminotransferase; ALT: alanine aminotransferase; GGT: gamma-glutamyl transferase; ALB: albumin; TB: total bilirubin; TP: total protein.
3.6. Effects of O. vulgare extract on serum Cr, BUN, Na+, and K+ levels in finasteride-treated mice
Finasteride significantly increased serum BUN (P=0.002) and Cr (P=0.004) while reducing serum Na(P=0.014) and K(P=0.008) compared to the normal group (Figure 3). The O. vulgare extract improved serum levels of all these parameters in a dose-dependent manner, and 400 mg/kg of the extract markedly reduced finasterideinduced changes (Figure 3).
3.7. Effects of O. vulgare extract on serum levels of pro- and anti-inflammatory cytokines in finasteride-treated mice
Finasteride significantly increased the serum levels of proinflammatory cytokines [IFN-γ (P=0.007), IL-12 (P=0.034), IL-6 (P=0.011), TNF-α (P=0.015), and IL-1β (P=0.024)] and reduced anti-inflammatory cytokine IL-10 (P=0.014) compared to the normal group (Figure 4A). On the other hand, O. vulgare extract at the dose of 400 mg/kg significantly reduced the serum levels of proinflammatory cytokines [IFN-γ (P=0.007), IL-6 (P=0.002), TNF-α (P=0.006), and IL-1β (P=0.009)] and increased IL-10 level (P=0.04) compared to the finasteride-treated group (Figure 4A).
3.8. Effects of O. vulgare extract on serum CAT and NOS activities and NO, MDA, and TAS levels in finasteridetreated mice
Serum activities of CAT (P=0.019) and NOS (P=0.021) and the serum level of TAS (P=0.011) were significantly decreased in the finasteride group. Moreover, significant elevations were observed in the serum levels of NO (P=0.002) and MDA (P=0.005) in finasteride-exposed mice compared to the normal group (Figure 4B). However, O. vulgare extract improved the serum levels of these oxidative stress-related parameters dose-dependently (Figure 4B).
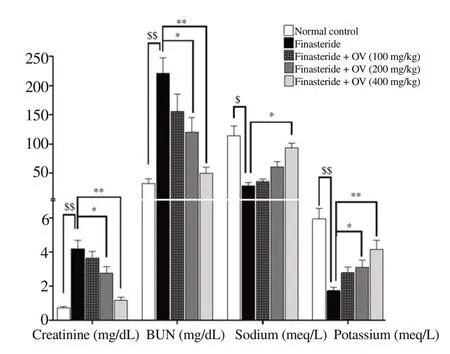
Figure 3. Serum levels of Cr, BUN, Na+ and K+ after treatment with Origanum vulgare extract. The data are expressed as mean±SD. $P < 0.05, $$P < 0.01 compared with the normal group; *P < 0.05, **P < 0.01 compared with the finasteride group. Cr: creatinine; BUN: blood urea nitrogen.
3.9. Histopathological findings
Pyknotic heterochromatin in the nuclei and the cytoplasm filled with eosinophilic granules were seen in the liver of finasteride-treated mice. Also, around the liver central venules and portal triad, dense lymphocytic infiltration was observed. The destruction of hepatocytes inhibited the bile and blood flow into sinusoidal spaces. Swelling, obstruction, and congestion were also seen in central venules (Figure 5 A1 & A2). Treatment with O. vulgare extract improved the hexagonal structure of hepatic lobules and cytoplasmic structure of hepatocyte. There were no lymphocytic infiltrations, congestion, and obstruction around the central venules(Figure 5B-D).
In the kidney, finasteride increased lymphocytic infiltrations around arteries and glomeruli, destructed the normal architecture of renal tissue, and finally induced necrosis in cortical and medullary renal tubules. Finasteride also damaged renal physiological function and resulted in congestion and hyperemia in renal arteries (Figure 6 A1 & A2). However, O. vulgare extract protected the renal tissue and preserved the normal histological structure. The extract also reduced lymphocytic infiltration, congestion, and hyperemia in renal arteries (Figure 6B-D).
4. Discussion
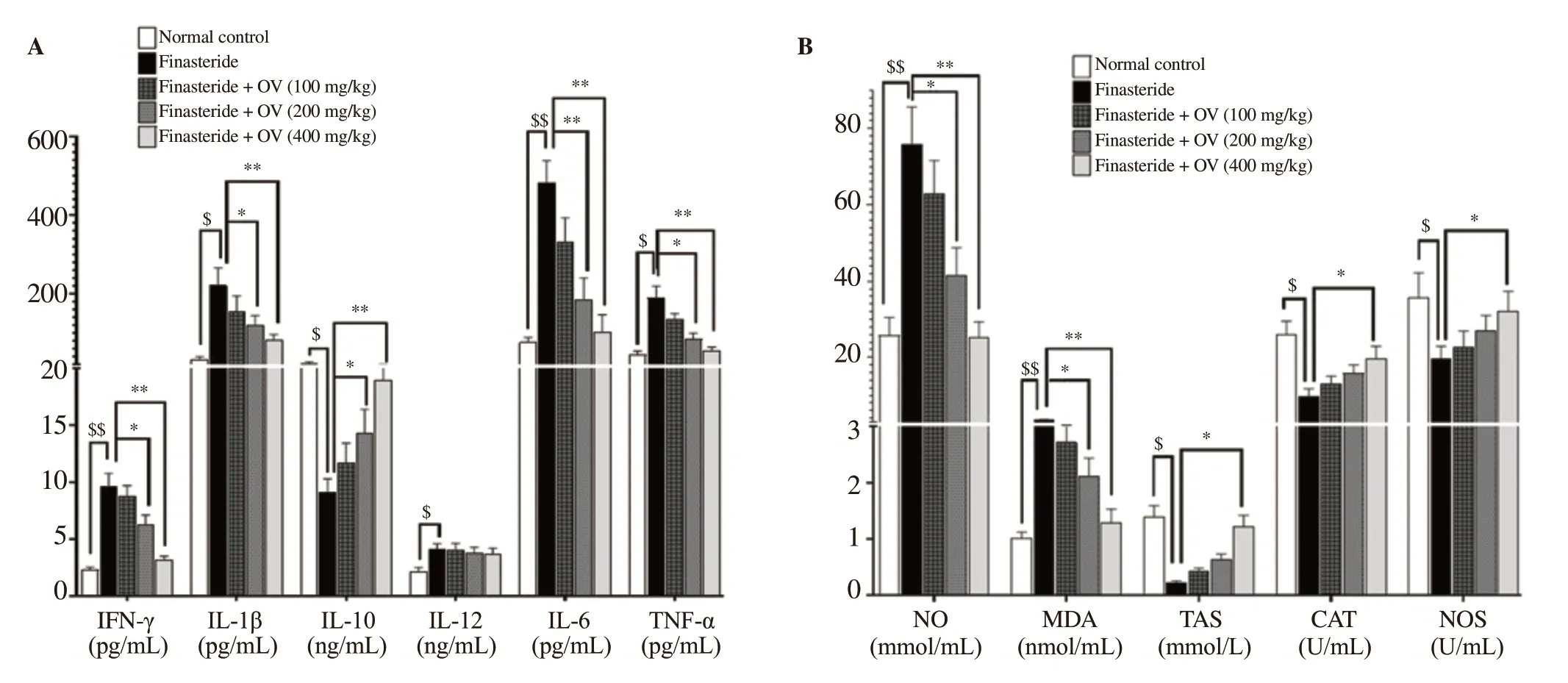
Figure 4. Effect of Origanum vulgare extract on oxidative stress and inflammatory cytokines in finasteride-exposed mice. (A) Serum levels of cytokines including IFN-γ, IL-6, TNF-α, IL-1β, IL-10, and IL-12. (B) Serum levels of CAT, NOS, NO, TAS, and MDA. The data are expressed as mean±SD. $P < 0.05, $$P < 0.01 compared with the normal group; *P < 0.05, **P < 0.01 compared with the finasteride group. CAT: catalase; TAS: total antioxidant status; NOS: nitric oxide synthase; MDA: malondialdehyde; NO: nitric oxide; IFN-γ: interferon-γ; IL-6: interleukin-6; TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β; IL-10: interleukin-10; IL-12: interleukin-12.

Figure 5. Histopathological results of the liver in mice. (A1-A2) The finasteride group shows parenchymal inflammation (*), hepatocellular necrosis (#), and sinusoidal obstruction ($). Endothelial injuries in small hepatic venules (&) lead to endothelial swelling, thrombosis, and luminal occlusion (^). (A2 & C) Sinusoidal dilatation and congestion, as well as the destruction of hepatocytes’ nuclei and some necrotic hepatocytes with the basophilic (1) and residual eosinophilic (2) bodies filled with granules are seen. (B, C and D) The treatment groups (100, 200 and 400 mg/kg/day of Origanum vulgare extract) show improved structure of the liver, which is more pronounced at the dose of 400 mg/kg. Hepatocytes have a normal structure (without eosinophilic granules). No congestion and obstruction of the central venule or lymphocyte infiltration is observed. (E) The normal group shows the normal structure of the hepatocytes, central venules, and liver arteries. n: normal hepatocytes; 1: basophilic hepatocytes; 2: eosinophilic hepatocytes; CV: central venule and NL: normal lobule [A1, A2, B, C, D, and E, H&E ×100 (scale bar = 100 μm)].

Figure 6. Histopathological results of the kidney in mice. (A1-A2) The finasteride group shows hyperemia in the renal arteries (*) and tubular necrosis (TN) in PCT and DCT in the glomeruli, and lymphocytic infiltration (LI) between the tubules. (B, C and D) The treatment groups (100, 200 and 400 mg/kg/day of Origanum vulgare extract) show improved structure of the liver, which is more pronounced at the dose of 400 mg/kg. No tubular necrosis, lymphocyte infiltration, and vascular hyperemia are observed. (E) The normal group shows the normal structure of the tubules, glomeruli, and renal arteries. G: normal glomerulus; BV: blood vessel; PCT: proximal convoluted tubule; DCT: distal convoluted tubule; US: urinary space; DG: glomerular degeneration; CT: collecting tubules; MD: macula densa [A1, A2, B, C, D, and E, PAS×100, (scale bar = 100 μm)].
The present study investigated the potential protective effects of O. vulgare extract against finasteride-induced liver and kidney injuries. As biochemical and histopathological examinations showed, O. vulgare extract effectively improved the structural and functional defects observed in the liver and kidney of finasteride-treated mice.
In the present study, 25 mg/kg of finasteride had adverse effects on biochemical parameters and histological structure of mouse liver and kidneys. Previous studies revealed that high (5 and 25 mg/kg/day and higher) or low (1 mg/kg/day) doses of finasteride increased the production of reactive oxygen species (ROS) and caused oxidative stress[25,26]. ROS then induced destructive effects via activating apoptotic pathways (i.e. ROS-mediated activation of GRP-78, p-IRE1, p-JNK, procaspase-3, p53, and caspase-3) in hepatocytes and epithelial cells of renal proximal convoluted tubule and distal convoluted tubule[27]. In this study, CAT and NOS activities were significantly reduced in finasteride-treated animals, which was in agreement with previous reports. Although apoptosis was not directly determined in the present research, remarkable apoptotic changes were observed in histopathological evaluation of the liver and kidney. Finasteride triggered kidney damage such as glomerulosclerosis, tubulointerstitial fibrosis, and infiltration of mononuclear cells via affecting androgen homeostasis and reducing androgen receptor expression[5,25]. On the other hand, androgen receptor has been described as a factor responsible for the deterioration of renal function[7].
Plants with anti-inflammatory, antioxidant, and anti-apoptotic properties inhibit the production of inflammatory cytokines and ROS and thereby protect hepatic and renal parenchymal cells against the damage caused by these factors and ultimately maintain their physiological function[28,29]. Protecting hepatocytes and renal tubular epithelial cells against damages on DNA and other cellular components that were induced by chemical drugs and toxins is attributed to the presence of phytocompounds such as flavonoids which have anti-oxidative and anti-inflammatory properties[30]. ROS results in oxidative stress accompanied by various injuries to tissues and organs, especially the liver[31], which was found in the current study. A previous study showed that treatment with O. vulgare extract prevented the increase of liver enzymes[32], which is in agreement with our results. In this study, treatment with O. vulgare leaf extract not only significantly decreased serum ALT, AST, and ALP but also prevented liver damage in finasteride-exposed mice. Moreover, O. vulgare extract normalized the increased levels of inflammatory cytokines and suppressed finasteride-induced oxidative stress. Our findings also indicated that O. vulgare promoted antioxidant activities by enhancing antioxidant enzymatic storage. Furthermore, O. vulgare extract with its antioxidant properties inhibited lipid peroxidation as evidenced by decreased levels of MDA and inflammatory cytokines. A previous study showed that O. vulgare leaf extract was rich in rosmarinic acid, caffeic acid, and other phenolic compounds such as flavonoids, which could inhibit lipid peroxidation[33].
Based on the results of this study, the significant elevation in the levels of pro-inflammatory cytokines including IFN-γ, IL-12, IL-6, TNF-α, and IL-1β was accompanied by tissue infiltration by immune cells. It has been reported that T cells and macrophages express testosterone-binding sites[34], and macrophages also demonstrate 5α-reductase activity similar to finasteride[35]. Thus, androgens could have directly affected immune cells (especially T and B lymphocytes) and contributed to the tissue infiltration. Besides, sex hormones not only control renal hemodynamics, mesangial cell proliferation, and extracellular matrix metabolism, but also affect the synthesis and release of some cytokines and other growth factors, which in turn affect the progression of renal diseases[36]. Furthermore, humoral and cellular immune responses are considered to be regulated by sex hormones[35]. Therefore, an impaired sex hormones ratio in finasteride-treated animals can directly modulate the function of immune cells and the production of pro-inflammatory factors. It has been shown that finasteride can induce apoptosis in hepatic and renal cells via increasing the secretion of inflammatory cytokines through c-Jun amino-terminal kinase pathways[37,38]. Therefore, the adverse effects of finasteride can be partly attributed to the inhibition of testosterone-dihydrotestosterone conversion, leading to impaired androgen/estrogen homeostasis and eventually characteristic pathomorphological and functional changes.
In the current study, finasteride-induced toxicity was associated with increased levels of TNF-α and NO, indicating intensified inflammatory and oxidative stress processes, respectively. On the other hand, the levels of these mediators were diminished significantly in the animals receiving O. vulgare extract compared to control groups. These findings were consistent with the results of similar studies[39,40]. Decline in the serum levels of TNF-α and NO proved that the nephroprotective and hepatoprotective effects of O. vulgare extract might be partly due to its high antioxidant capacity (as shown in DPPH and FRAP assays). These results were also in line with previous reports. Sharifi-Rigi et al. reported that the oral administration of O. vulgare extract increased renal superoxide dismutase and CAT activities and decreased serum level of MDA and the gene expression of TNF-α in rat models of paraquat-induced nephrotoxicity[41]. In another study, O. vulgare extract protected the liver against paraquat-induced hepatotoxicity in rats partly by scavenging free radicals and improving hepatic structure and function[42]. In histopathological study, we observed mononuclear cell infiltration and fibrosis around renal atrophic lesions. It has been reported that T cells can express TGF-β and directly induce adjacent tubular epithelial cells to transform into proliferating fibroblasts that migrate across the tubular basement membrane and generate fibrotic lesions in the renal interstitium[43].
In line with the results of other studies, LC-ESI/MS analysis showed that O. vulgare hydroalcoholic extract contained flavonoids such as quercetin, apigenin, genistein, α-tocopherol, kaempferol, coumaric acid, epigallocatechin, and daidzein, some of which have potent antioxidant properties, and others are effective in reinforcing the hypothalamic-pituitary-gonadal axis. Studies have shown that some polyphenolic compounds of O. vulgare extract, such as apigenin[44], quercetin[45], and ferulic acid[46], regulate the secretion of testosterone from Leydig cells and protect Sertoli cells and the spermatogenic lineage against ROS destructive effects, which can reduce the release of inflammatory cytokines from damaged cells and ultimately maintain the structure of tissues and cells.
In conclusion, the present study showed that the hydroalcoholic extract of O. vulgare prevented the nephrotoxic and hepatotoxic effects induced by finasteride. We also demonstrated that these protective effects were executed through the antioxidant and antiinflammatory properties of the extract. This plant can be used to improve tissue injuries in patients with kidney and liver problems. It is recommended to purify the effective compounds of this plant and investigate their therapeutic effects in drug discovery studies.
There were some limitations in the current study. Pro-apoptotic and anti-apoptotic factors including NF-κB, Bax, p53, and Bcl-2 were not assessed in the present study. Therefore, it is advisable to investigate the possible anti-apoptotic properties of O. vulgare leaf extract in future studies to further understand its mechanisms of action.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Acknowledgments
The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University, Abha, KSA for funding this work through Research Group Project under grant number R.G.P.2/80/41 and also Wuxi Municipal Health and Family Planning Commission (WXQ201832).
Funding
This work was financially supported by King Khalid University, Abha, KSA through Research Group Project under grant number R.G.P.2/80/41 and Wuxi Municipal Health and Family Planning Commission (WXQ201832).
Authors’ contributions
QFS, SXC, AE, and HK designed and conducted the experiment, and performed data analysis. ZFT and XYS contributed to manuscript writing and performing experimental study. FJ, HTW, AE, HK, and ZYD participated in definition of intellectual content, literature search, statistical analysis, and manuscript preparation. AE got the research fund and DM performed language editing. All the authors reviewed the data, read, and approved the manuscript.
 Asian Pacific Journal of Tropical Biomedicine2021年5期
Asian Pacific Journal of Tropical Biomedicine2021年5期
- Asian Pacific Journal of Tropical Biomedicine的其它文章
- Protective effects of rice bran hydrolysates on heart rate variability, cardiac oxidative stress, and cardiac remodeling in high fat and high fructose diet-fed rats
- Aloin attenuates chronic constriction injury-induced neuropathic pain in rats by inhibiting inflammatory cytokines and oxidative stress
- Argemone mexicana extract alleviates gastrointestinal disorders by stimulating muscarinic receptors and blocking voltage-gated L-type calcium channels
- Honokiol attenuates oxidative stress-induced cytotoxicity in human keratinocytes via activating AMPK signaling