
杜仲ARF基因家族全基因组鉴定和表达分析*
2021-04-10 04:07:16刘闵豪周轩辕李周岐范睿深徐郡儡
林业科学 2021年3期
刘闵豪 李 龙 叶 靖 周轩辕 李周岐 范睿深 徐郡儡
(西北农林科技大学林学院 杨凌 712100)
杜仲(Eucommiaulmoides)是我国特有的以药用为主且具有其他多种用途的植物。杜仲树皮为传统中药材,其主要药用成分为环烯醚萜类、黄酮类、苯基丙烷、多糖、氨基酸、其他萜类和脂肪族(张京京等, 2014)。此外,杜仲树皮、树叶及果实中含有杜仲胶(反式聚异戊二烯),能够广泛应用于电器工业、化学工业和电讯器材工业中,拥有巨大的潜力。杜仲具有极高的开发利用价值,基因功能研究是其资源开发中一项十分重要的基础性工作,以往人们对于药用成分与杜仲胶合成途径的基因研究具有高度的热情,这使得杜仲生长发育相关基因的研究受到了忽视。2018年,杜仲首个全基因测序完成(Wuyunetal., 2018),标志着杜仲基因研究进入了一个新的阶段。利用杜仲全基因组测序数据,研究杜仲生长发育相关基因家族,能够弥补杜仲生长发育相关基因研究的不足,为杜仲的基因挖掘与物种特性研究奠定基础。
生长素(auxin, Aux)在植物中最主要的成分为吲哚乙酸(indole-3-acetic acid, IAA),是发现的第1种促进植物生长的植物激素。生长素的信号转导控制着植物各器官和组织对生长素的反应,在生长素对植物生长的调控中发挥重要作用。
生长素响应因子(auxin response factors, ARF)是生长素信号转导途径中重要的转录因子,其能够特异地结合生长素响应基因启动子区域的生长素响应原件(auxin response element, AuxRE)TGTCTC序列,调控生长素响应基因的表达,从而能够与多种途径的激素信号协同作用(Hagenetal., 2002)。ARF蛋白一般由3部分组成: N端B3型DNA结合区域(DNA-binding domain,DBD)、响应生长素应答的中间区域(middle region,MR)和C末端二聚化结构区域(C-terminal domain,CTD)(Tiwari, 2003)。ARF基因家族成员众多,表达模式复杂。目前的研究表明,大多数ARF基因对环境因子的反应较弱,其表达受microRNAs和反式作用siRNA(ta-siRNA)转录后调控(Liuetal., 2011a);并且发现了拟南芥(Arabidopsisthaliana)中microRNA167(miR167)靶向ARF6和ARF8(Wangetal., 2015), microRNA160(miR160)靶向ARF10、ARF16和ARF17(Linetal., 2015), TAS3 ta-siRNAs靶向ARF2、ARF3和ARF4(Williamsetal., 2005)。
近年来,随着基因组和转录信息的不断完善,越来越多的植物物种进行了ARF的相关研究,在模式植物和作物中的研究比较深入,包括拟南芥(Okushimaetal., 2005)、水稻(Oryzasativa)(Wangetal., 2007)、玉米(Zeamays)(Xingetal., 2011)、大麦(Hordeumvulgare)(Huseyin, 2018)和番茄(Solanumlycopersicum)(Wuetal., 2011)等。由于木本植物基因组比较复杂,只有部分果树和用材树种鉴定过ARF基因,主要有苹果(Malusdomestica)(Luoetal., 2014)、甜橙(Citrussinensis)(Lietal., 2015)、葡萄(Vitisvinifera)(Wanetal., 2014)、毛果杨(Populustrichocarpa)(Kallurietal., 2007)和巨桉(Eucalyptusgrandis)(Hongetal., 2014)等。
本研究从杜仲全基因组测序数据中鉴定杜仲ARF基因家族全体成员,并从杜仲microRNA测序数据中获取杜仲的miR160与miR167序列,利用生物信息学方法对杜仲ARF基因家族成员进行分析,了解其基因外显子/内含子结构、蛋白结构、进化关系和miR160/167的调控预测等。同时利用杜仲转录本全长数据测序结果,分析杜仲ARF基因家族成员在叶片各生长发育时期的表达水平,再通过实时定量反转录PCR(qRT-PCR)方法分析杜仲ARF基因家族成员与miR160/167在杜仲不同发育时期的根、茎、叶中的表达量,分析ARF基因家族成员的时空表达模式和miR160/167的调控模式,初步预测其功能。本研究为EuARF基因的进一步功能研究奠定了基础,弥补了杜仲生长发育相关基因研究的不足。
1 材料与方法
1.1 试验材料 供试的植物材料为西北农林科技大学林学院实验苗圃(108.4°E,34.2°N)种植的杜仲优良品种杂交后代。于6月选取4年生同一亲本的杂交后代,混合采集其幼嫩叶片、成熟叶片、幼嫩茎段、成熟茎段、幼嫩根及成熟根,采摘后立即冻存到液氮中,随后带回实验室,并转存到-80 ℃冰箱中备用。所有样品均为3个生物学重复。
1.2 杜仲ARF基因家族的鉴定 从基因组数据网站Genome Warehouse下载发布的杜仲基因组测序数据(http:∥bigd.big.ac.cn/gwh/Assembly/13/show),构建本地数据库。以ARF结构域(PF06507)蛋白序列、拟南芥的AtARF蛋白序列以及水稻的OsARF蛋白序列为模板对基因组测序数据的蛋白数据库进行本地BlastP比对搜索,初步筛选出E≤10-5的蛋白序列为假定的EuARF蛋白。对这些假定的EuARF蛋白,使用SMART (http:∥smart.embl-heidelberg.de/)在线分析工具鉴定蛋白的保守序列,删除不含ARF蛋白特征结构域和特征结构域置信度低的基因。
1.3 杜仲ARF基因家族系统进化树分析 使用ClustalX软件对筛选出来的杜仲ARF基因家族蛋白与下载的拟南芥的AtARF蛋白序列、水稻的OsARF蛋白序列、欧美杨(Populus×euramericana)的PeARF蛋白序列进行多序列比对,蛋白序列均下载自PlantTFDB。比对结果使用 MEGA5.1软件构建系统进化树,构建采用邻接法(neighbor-joining method, NJ),自展值(bootstrap)设定为1 000。根据最后的进化树,将鉴定出的杜仲ARF基因家族按照与拟南芥AtARF基因家族比对结果的相似性进行命名。
1.4 杜仲ARF基因家族的生物信息学分析 使用基因结构显示系统(GSDS2.0)网站(http:∥gsds.cbi.pku.edu.cn/)比较每个EuARF基因的cDNA序列与基因组序列,生成其外显子/内含子结构。使用ProtParam在线软件(http:∥web.expasy.org/protparam/)分析鉴定出的EuARF蛋白的理化性质。利用MEME工具5.0.3版(http:∥meme-suite.org/tools/meme)分析EuARF蛋白的保守基序,预测的基序长度设为6~200个氨基酸,基序数量设为20,分布设为每个序列出现1次。
1.5 miRNA靶基因结合位点预测 使用本地Blastn对前期研究获得的杜仲miRNA测序数据进行比对,筛选杜仲miR160/167。使用RNAhybrid在线工具(https:∥bibiserv.cebitec.uni-bielefeld.de/rnahybrid)对eu-miR160/167和EuARFs的cDNA序列进行分析,预测可能的靶基因以及结合位点。
1.6EuARFs基因表达分析 利用前期研究获得的杜仲转录本全长数据库(测序使用4组生长时期叶片样本,分别为叶芽、3 cm长的生长叶、新的完全展开的幼叶和完全展开后30天的成熟叶,每组3个生物学重复)检索EuARFs的FPKM(fragments per kilobase million)值,使用HemI 1.0软件分析EuARFs在叶片生长过程中的表达谱。采用qRT-PCR方法分析EuARFs与eu-miR160s/miR167s在不同发育阶段(幼嫩和成熟)不同器官(根、茎、叶)中的表达量,从而验证EuARFs在叶片中的表达模式,初步分析EuARFs在根与茎中的表达模式,预测eu-miR160s/miR167s对EuARFs的调控作用。
1.7 RNA的提取与qRT-PCR的具体方法 使用Trizol法(Ambion, https:∥www.thermofisher.com/)提取样品中的总RNA,并使用TAKARA(TaKaRa, http:∥www.takara-bio.com/)的RNase-free DNase处理去掉残留的DNA。
对各样品的RNA使用TAKARA的反转录试剂盒进行反转录合成cDNA第1链,其中eu-miR160s/miR167s的反转录使用茎环法,茎环引物见表1。使用TAKARA的荧光定量试剂盒进行qRT-PCR试验,内参基因选择为EuUBC(基因登录号为: MH890470)(Yeetal., 2018),采用2-ΔΔCt法计算相对表达量。从EuARFs的保守区域设计用于qRT-PCR的引物,使用稀释了1、5、25、125和625倍的幼嫩叶片cDNA溶液进行每个引物的qRT-PCR试验,建立标准曲线,根据扩增效率对引物进行筛选,最后获得的引物见表2。eu-miR160/167的qRT-PCR使用的反向引物为通用引物,正向引物见表1。
2 结果与分析
2.1 EuARF蛋白的鉴定及理化性质 经过转录本数据本地BlastP比对和SMART保守结构域分析后,共鉴定出18个杜仲ARF蛋白序列,通过生物信息学进一步验证获得的EuARF蛋白的基本信息和理化性质(表3)。所有EuARF蛋白均含有DBD与ARF结构域,为真正的ARF蛋白,大部分EuARF蛋白含有CTD结构域,不含CTD结构域的EuARF蛋白为EuARF3.1、EuARF3.2、EuARF8.2、EuARF10.1、EuARF10.2、EuARF16 和EuARF17; EuARF蛋白的长度为273(EuARF8.2)~1 062个(EuARF19.1)氨基酸,预测的分子质量为30.64~117.90 kDa; 理论等电点(pI)为5.36(EuARF10.2)~9.68(EuARF8.2),其中16个EuARF蛋白的等电点都小于7,呈弱酸性,在酸性细胞环境中发挥作用,仅有EuARF2.1和EuARF8.2的等电点大于7,呈碱性。
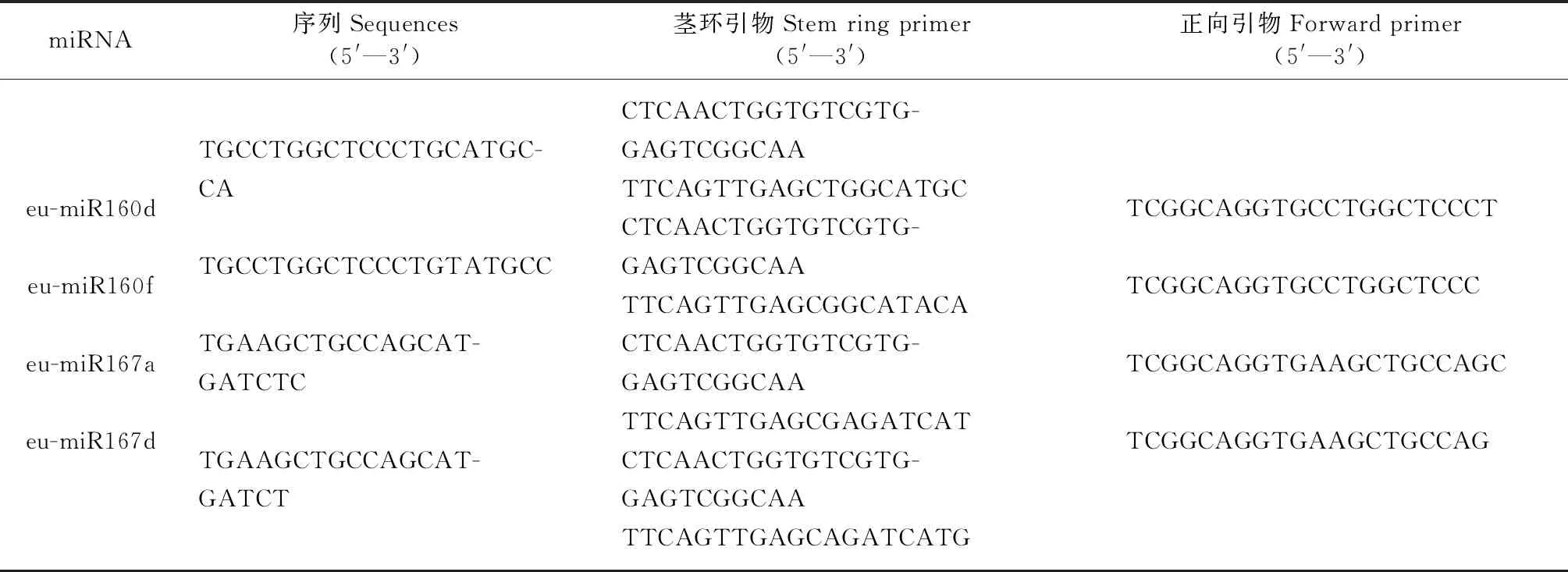
表1 eu-miR160s/eu-miR167s的序列与引物

表2 EuARFs基因的引物
2.2 EuARF蛋白的系统进化 利用邻接法(NJ)构建了拟南芥、水稻、欧美杨和杜仲ARF基因家族的系统发育进化树(图1),可以将18个EuARFs分到5个亚族。EuARF3.1、EuARF3.2和EuARF4属于Ⅰ亚族(AtARF3/4同源),EuARF10.1、EuARF10.2、EuARF16和EuARF17属于Ⅱ亚族(AtARF10/16/17同源),EuARF1、EuARF2.1、EuARF2.2、EuARF9和EuARF18属于Ⅲ亚族(AtARF1/2同源),EuARF6.1、EuARF6.2、EuARF8.1和EuARF8.2属于Ⅴ亚族(AtARF6/8同源),EuARF19.1和EuARF19.2为Ⅵ亚族(AtARF7/19同源),EuARF蛋白中没有Ⅳ亚族蛋白(AtARF5同源)。
从进化树的分支上来看,在Ⅰ亚族和Ⅵ亚族中,EuARFs蛋白与PeARFs的亲缘关系较近; 在Ⅱ亚族和Ⅴ亚族中,EuARFs蛋白与OsARFs和AtARFs的亲缘关系更近; 而在Ⅲ亚族中,EuARF2.1蛋白与AtARF2亲缘关系较近,EuARF9、EuARF18与PeARF9、PeARF18亲缘关系更接近。
2.3 EuARF蛋白的保守结构域和基序 SMART分析的EuARF蛋白保守结构域以及meme分析的保守基序结果如图2所示,图2a是根据多重序列比较构建的18个EuARF蛋白的进化树,以方便比较各ARF亚族的保守结构域和基序特征。保守结构域分析(表3,图2b)表明大多数EuARF蛋白都包含3个结构域(DBD、ARF和CTD),而EuARF10.1、EuARF10.2、EuARF16、EuARF17、EuARF3.1、EuARF3.2和EuARF8.2不含CTD结构域,其中EuARF3.1、EuARF3.2属于Ⅰ亚族,EuARF10.1、EuARF10.2、EuARF16和EuARF17属于Ⅱ亚族。比较其基序(图2c)发现,DBD结构域由Motif 1、2、9和11组成,ARF结构域由Motif 5、6和13或Motif 5、6和19组成,而CTD结构域由Motif 7和10组成,每个基序的氨基酸组成如图2d所示。
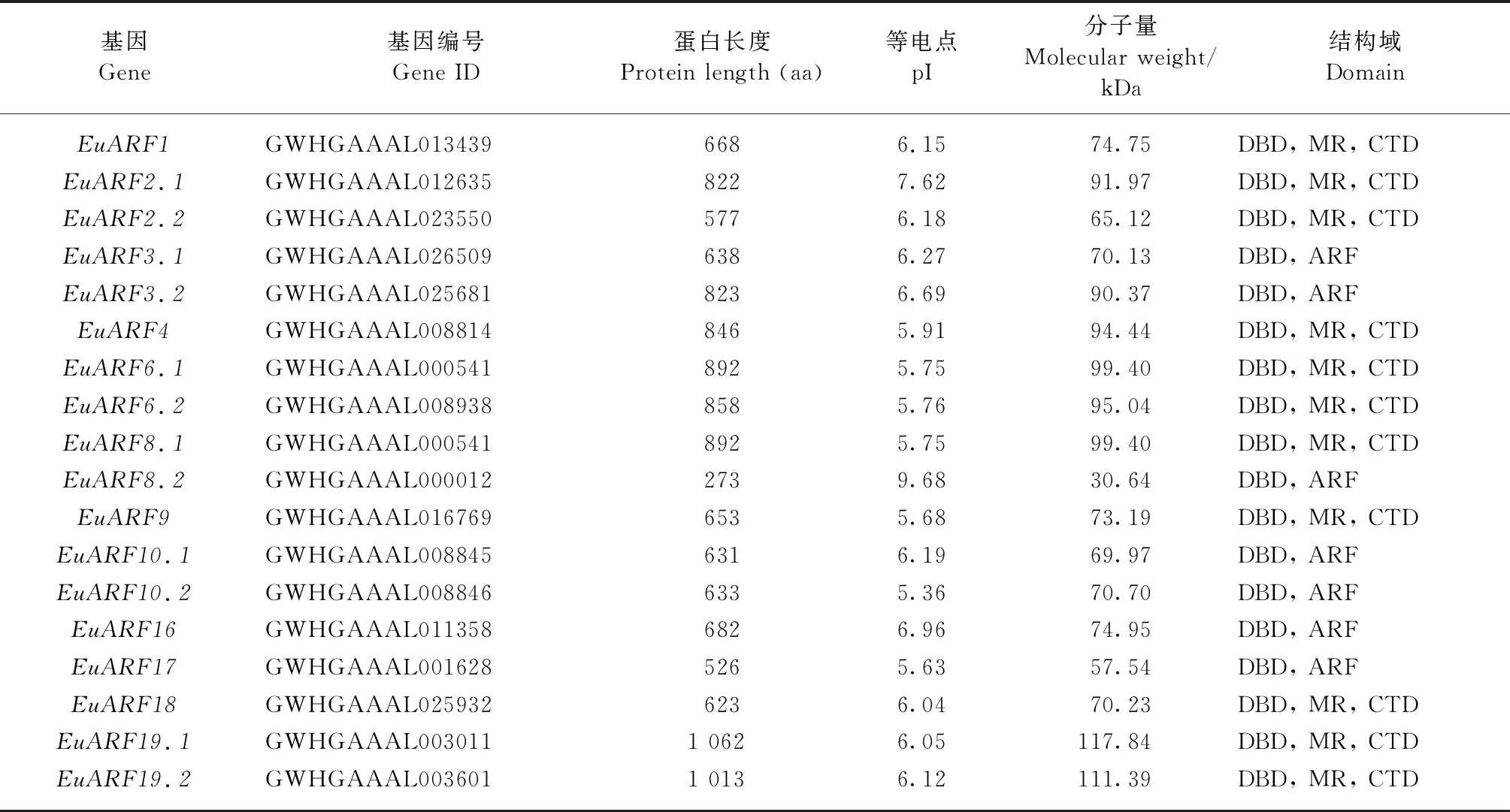
表3 杜仲ARF基因家族基本信息①
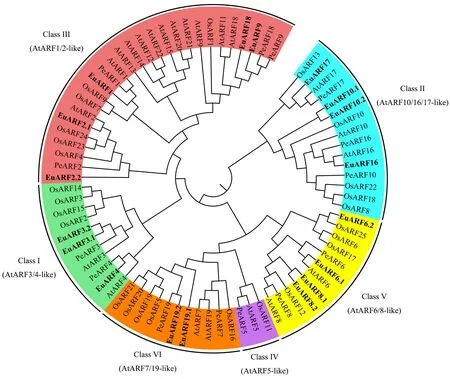
图1 ARF基因家族系统发育进化树

图2 EuARF蛋白的保守结构域与基序
2.4EuARF基因结构 通过比较EuARF基因的cDNA序列和基因组序列,可以获得EuARF基因的外显子/内含子结构(图3),其中图3a是根据多重序列比较构建的18个EuARF蛋白的进化树,以方便将基因结构分为亚族比较。从基因结构中可以看出EuARF基因均含有2个或2个以上的外显子,同一亚族的EuARF基因的外显子/内含子结构比较相似,例如第Ⅱ亚族的EuARF基因只有2或3个外显子,而其他亚族EuARF基因的外显子更多。

图3 EuARF基因的外显子/内含子结构
2.5EuARF基因受miR160/167调控的靶位点 在前期研究获得的杜仲miRNA测序数据中共鉴定到4条miR160/167,命名为eu-miRNA160d、eu-miRNA160f、eu-miRNA167a和eu-miRNA167d,eu-miR160/167的序列见表1。RNAhybrid靶位点预测的结果(图4)显示EuARF基因家族中有7个基因受到eu-miR160/eu-miR167的调控,其中eu-miR160s调控Ⅱ亚族的EuARFs(EuARF10.1、EuARF10.2、EuARF16和EuARF17),eu-miR167s调控Ⅴ亚族的EuARFs(EuARF6.1、EuARF6.2和EuARF8.1),该结果与拟南芥中miRNA对AtARFs的调控情况相同(Wangetal., 2015; Linetal., 2015)。
2.6EuARFs和eu-miR160s/167s的表达模式 在前期研究中,本课题组通过单分子测序获得了杜仲叶片不同生长时期的转录本全长数据。4个不同生长时期的叶片分别为叶芽(L1)、生长叶(3 cm长,L2)、幼叶(新的完全展开的叶,L3)和老叶(完全展开后30天,L4)。在转录本全长序列数据中共检索到17个EuARF基因的FPKM(EuARF17没有检索到),通过HemI 1.0构建的基因表达谱如图5所示,可以看出: 在叶片的所有生长发育时期,Ⅵ亚族的EuARF19.2的表达明显高于其他EuARFs; 在幼叶(L3)中,Ⅴ亚族的EuARFs的表达量较高;EuARF1、EuARF2.1、EuARF2.2、EuARF10.1在叶芽(L1)和老叶(L4)中表达较高; Ⅴ亚族的EuARFs除EuARF8.2外均在生长叶(L2)中表达较高。此外,Ⅰ亚族EuARFs和Ⅲ亚族的EuARF9在叶片不同生长时期的表达均较低。EuARFs在叶片不同生长时期的表达热图(图5)可以初步反映其在叶片生长过程中存在调控作用。
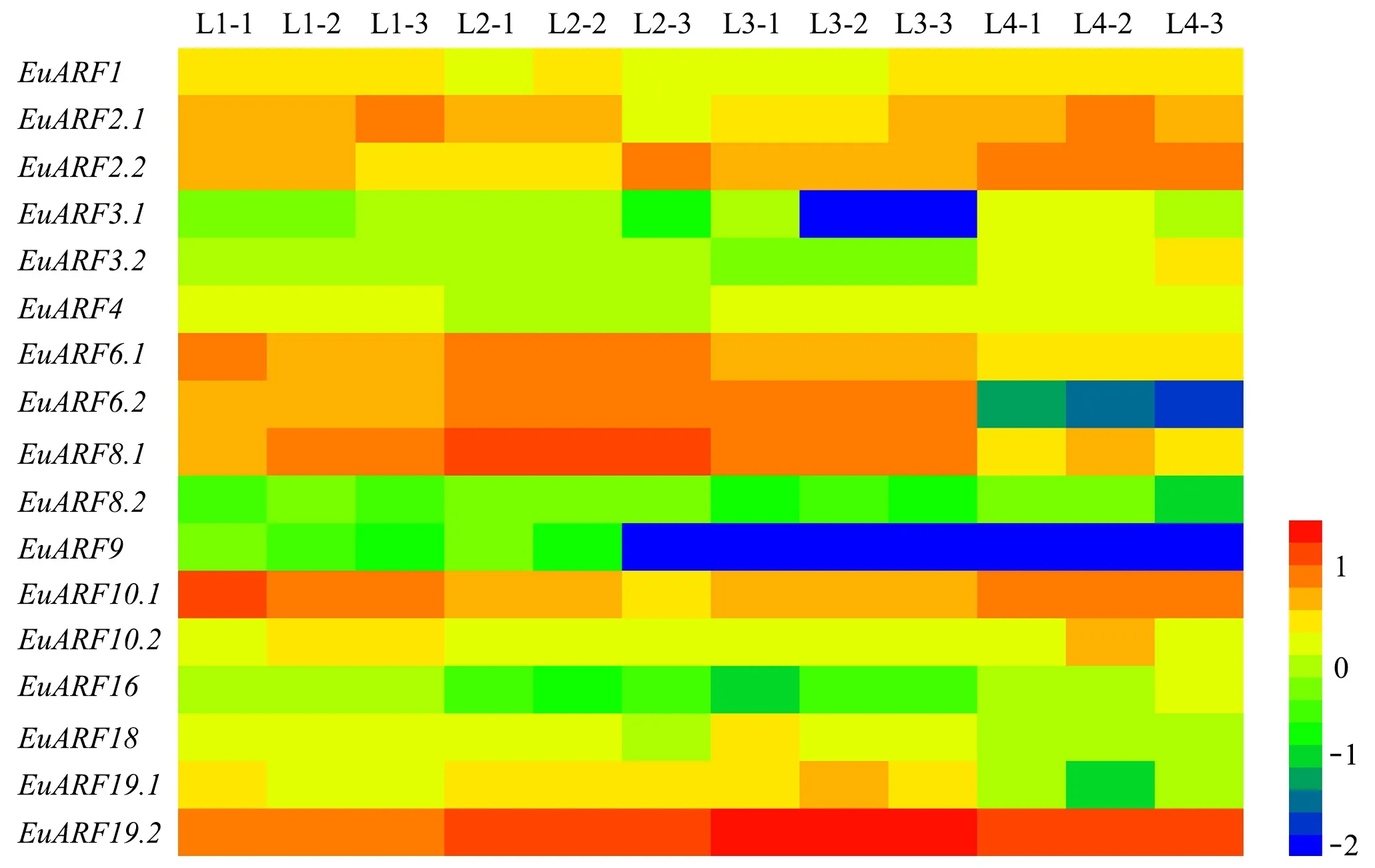
图5 EuARF基因在不同生长时期叶片中的表达谱
为了验证EuARF基因在叶片中的表达模式,初步分析EuARF基因在其他器官中的表达模式,采用qRT-PCR方法,比较了正常生长条件下不同发育阶段(幼、熟)器官(叶、根、茎)中EuARF基因的表达水平。结果(图6a、b)表明,在正常生长条件下不同发育阶段(幼嫩和成熟)的器官(根、茎和叶)中EuARFs均有表达,不过表达量有一定差异。在根中,EuARFs的表达量最高,并且在成熟根中的表达量普遍高于幼嫩根,而在叶和茎段中却相反,表现为在幼嫩的器官中表达量更高。在表达水平上,EuARFs之间存在较大区别。EuARFs在叶片中的表达与转录本测序数据的表达谱相似,其中EuARF19.2在幼叶中的表达量最高; V亚族的EuARFs(EuARF6.1、EuARF6.2和EuARF8.1)在叶和茎中的表达水平较高,并表现为在幼嫩和成熟期均有表达; Ⅱ亚族的EuARFs(EuARF10.1、EuARF10.2、EuARF16和EuARF17)和Ⅴ亚族的EuARF8.2相较于其他EuARFs在所有器官中表达水平都较低。
eu-miR160s/eu-miR167s在不同发育阶段器官中的表达模式(图6c)显示,eu-miR160s在幼根中的表达较高,其靶基因Ⅱ亚族的EuARFs在根中均有一定表达,但与其他亚族的EuARFs相比,这些靶基因在根中的表达量属于中等偏低水平,这可能说明这些基因在根中的表达受到了eu-miR160s的负调控。eu-miR167s也存在相同的结果,eu-miR167s在成熟根和茎中几乎不表达,其靶基因EuARF6.1、EuARF6.2和EuARF8.1在成熟根和茎中表达水平最高,这表明eu-miR167s可能也起到负调控作用。
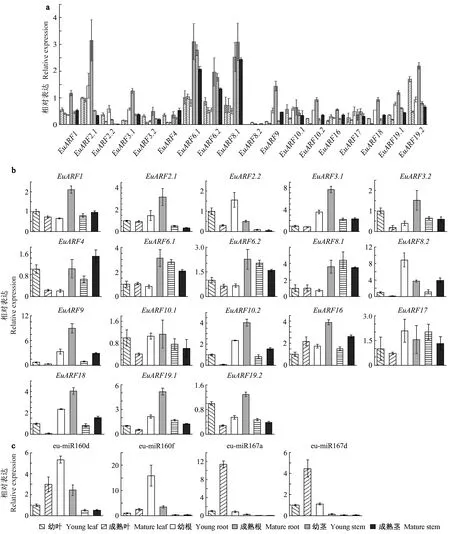
图6 EuARFs基因和eu-miR160s/eu-miR167s在不同发育阶段不同器官的表达
3 讨论
ARF转录因子在植物中以家族的形式存在,一般情况下被分为6个亚族(Xingetal., 2011; Tangetal., 2018; Huseyin, 2018),在有的研究中也被分为5个亚族(Lietal., 2015; 董晨等, 2017)。本研究共鉴定出18个EuARFs,根据6个亚族的分类可以分到其中5个亚族中(图1),只有IV亚族中没有EuARF基因,同亚族的EuARF基因都具有相似的外显子/内含子结构(图3),同亚族的EuARF蛋白也具有相似的保守结构域和基序(图2),这些结果验证了基于多序列比对后构建的进化树的可靠性。
在家族成员数量上,EuARFs小于AtARFs,主要原因是Ⅲ亚族的EuARFs数量(3个)远小于AtARFs(12个),这种现象也存在于其他植物种,包括大麦 (Huseyin, 2018)、番茄(Wuetal., 2011)、苹果(Luoetal., 2014)、甜橙(Lietal., 2015)、巨桉(Hongetal., 2014)和荔枝(Litchichinensis)(董晨等, 2017)等。在早期的研究中,这些多出来的AtARFs被单独分为一组(Xingetal., 2011; Wuetal., 2011; Wanetal., 2014; Yangetal., 2014)。这表明,杜仲可能与这些植物一样,在进化过程中丢失了一小部分的ARF基因。
在大部分植物的ARF基因家族中都存在不含CTD结构域的ARF蛋白,杜仲的ARF基因家族中也同样如此。SMART结果显示,Ⅰ亚族和Ⅱ亚族的EuARF蛋白以及Ⅴ亚族的EuARF8.2不含CTD结构域。在拟南芥和其他物种也有相似的情况,与Ⅰ亚族EuARF3.1、EuARF3.2同源的AtARF3也不含CTD结构域,并与EuARF3.1、EuARF3.2一样表现为完全缺失CTD结构域; Ⅱ亚族的AtARFs(AtARF10、16和17)也不含CTD结构域,并且与Ⅱ亚族的EuARFs一样表现为CTD结构域不完整。这些不含CTD结构域的ARF蛋白可能不参与生长素的信号转导途径而行使独立的功能(Liuetal., 2011b),在本研究中则发现这些不含CTD结构域的EuARFs表达水平相对较低,尤其是Ⅱ亚族的EuARFs,这可能与受到eu-miR160s的负调控有关,表明这类EuARFs可能在eu-miR160s的负调控下参与其他信号转导途径。此外,通过比较基序(图2c)可以发现EuARF8.2蛋白相比于其他Ⅴ亚族的EuARF蛋白除了缺少CTD结构域外,还缺少了Motif 3、4、8、12、15和17,这些基序位于DBD与ARF结构域两侧,其丢失可能使EuARF8.2丧失了部分功能。
本研究中发现EuARF基因家族中有7个基因受到eu-miR160s/eu-miR167s的调控,其中eu-miR160s调控Ⅱ亚族的EuARFs,eu-miR167s调控除了EuARF8.2的V亚族的EuARFs,这与拟南芥中的调控情况相同(Wangetal., 2015; Linetal., 2015)。通过表达模式分析(图6c)可以看出eu-miR160s在幼嫩的根中表达水平极高并且在其他不同发育阶段的器官中也有表达,而Ⅱ亚族的EuARFs在根中均有一定表达,但与其他亚族的EuARFs相比,其在根中的表达量属于中等偏低水平,这表明eu-miR160s在根的生长发育中对Ⅱ亚族EuARFs可能有负调控作用,从而调控根的生长; 在拟南芥的研究中发现Ⅱ亚族的AtARFs中,AtARF10和AtARF16对根冠以及根部分生组织的发育起到调控作用(Wangetal., 2005),杜仲的Ⅱ亚族EuARFs可能拥有相似的功能。eu-miR167s则是主要在叶片中表达,尤其是在成熟的叶片中,其调控的Ⅴ亚族EuARFs在根、茎、叶中的表达水平均较高(图6a),不过对比其他EuARFs基因在根、茎、叶的相对表达水平(图6b)可以发现,Ⅴ亚族EuARFs在叶片中的表达水平远低于根和茎,推测Ⅴ亚族的EuARFs在叶片中的表达受到了负调控,这表明Ⅴ亚族EuARFs可能在eu-miR167s的负调控下在叶片发育中发挥重要的作用。
进一步比较EuARFs的表达水平与其同源ARFs的功能研究结果可以对EuARFs的具体功能进行预测,这为进一步研究这些基因的功能奠定了基础。例如,研究已经证实AtARF8通过调节GH3基因的表达对幼苗生长产生影响(Wuetal., 2006),本研究中也发现与AtARF8同源的EuARF8.1在幼嫩的茎中表达水平极高,并且在叶片发育中受到eu-miR167s的负调控; Zhang等(2015)发现OsARF19控制着水稻叶夹角和侧根发育,并且AtARF7/AtARF19被证实可激活LBD/ASLS基因从而促进叶片的增大(Okushimaetal., 2007),本研究则发现与OsARF19和AtARF7/AtARF19同源的EuARF19.2在叶和根中有很高的表达水平; 先前的研究表明,AtARF2除了参与花器官的发育,还对拟南芥叶和根的发育有调节作用(Okushimaetal., 2005),与AtARF2同源的EuARF2.1在叶和根中也有很高的表达水平。
4 结论
杜仲是重要的叶用经济树种,研究杜仲叶片的生长发育对杜仲育种研究有重要意义。本研究在杜仲全基因组测序数据库中共鉴定出18个杜仲ARF基因,这些EuARFs可以分到5个亚族中,基因结构、理化性质以及蛋白结构分析说明同亚族的EuARFs具有相似的结构与功能,其中Ⅱ亚族的EuARFs受eu-miR160s调控,V亚族的EuARFs除了EuARF8.2均受eu-miR167s调控。EuARFs的转录本数据表达谱显示,在幼叶(L3)和生长叶(L2)中,V亚族的EuARFs的表达量较高,此外,EuARF19.2的表达明显高于其他EuARFs。qRT-PCR试验结果显示,EuARFs在正常生长条件下不同发育阶段(幼嫩和成熟)的器官(根、茎和叶)中均有表达,在根中表现为在成熟阶段上调表达,叶和茎段中表现为在成熟的器官中下调表达。eu-miR160s主要在幼嫩的根中表达,eu-miR167s则是主要在成熟的叶片中表达。综合来看,V亚族中EuARFs对幼苗的生长发育具有调控作用,在幼嫩的茎中表达水平极高,在叶片中受到eu-miR167s的负调控从而对其生长发育产生影响; 此外,EuARF19.2在叶片中的表达显著高于其他EuARFs,可以推测其对杜仲叶片生长具有重要的调控作用,具有极高的研究价值。本研究初步揭示了大部分EuARFs在植物生长发育中发挥的作用,为杜仲EuARFs基因功能的进一步研究奠定了基础。
猜你喜欢
江苏农业科学(2022年17期)2022-09-24 11:42:26
四川大学学报(自然科学版)(2021年4期)2021-07-15 09:10:48
中国农业科学(2020年5期)2020-03-30 02:02:46
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
中成药(2018年8期)2018-08-29 01:28:14
上海师范大学学报·自然科学版(2016年5期)2016-12-14 06:31:20
陕西画报(2016年1期)2016-12-01 05:35:28
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
小雪花·小学生快乐作文(2015年9期)2015-09-29 23:06:01
