
鹅掌楸天然林物种组成与林分空间结构特征研究
2021-04-10 05:23:30薛卫星郭秋菊艾训儒黄阳祥李玮宜
林业科学研究 2021年2期
薛卫星,郭秋菊*,艾训儒,姚 兰,朱 江,黄阳祥,李玮宜,罗 西,刘 毅
(1. 湖北民族大学生物资源保护与利用湖北省重点实验室,湖北 恩施 445000;2. 湖北民族大学林学园艺学院,湖北 恩施 445000;3. 湖北星斗山国家级自然保护区管理局,湖北 恩施 445000)
森林中物种的不同组成及其在空间分布的不同格局构成了森林的空间结构,而这些空间结构特征的变化会对森林的发展、演替产生重要影响。近几十年来,人们对森林结构的量化进行了大量研究[1],并提出不同的结构指数来量化森林的空间结构特征[2-3],其中一些学者提出的空间结构指数在实际调查中较为费时、费力,因此在应用中有一定限制。基于此,惠刚盈等[4]提出基于最近相邻木的林分空间结构指数,通过以最近相邻木组成的林分空间结构单元为基础构建林分空间结构参数体系,客观地分析评价林分空间结构状态。具体从描述林木空间分布格局的角尺度,体现林木大小差异的大小比数,描述树种混交程度的混交度以及描述林木密集程度的密集度4 个方面综合反应,然而,单一的林分结构参数不足以描述复杂的林分结构,所以,更多的研究中将两个或多个结构参数结合作为有机整体。通过多元分布更全面与客观地描述森林状态[5-6],更完整地体现森林结构和功能,且通过结构参数的多元分布可弥补林分结构参数与林木竞争指数之间的差距,可对森林整体结构进行深入全面的解释,为基于空间结构分析的森林经营方法提供科学依据,也为精准提高森林质量提供有效途径[7]。
鹅掌楸(Liriodendron chinense(Hemsl.) Sarg.)是我国特有珍稀植物,落叶大乔木,星散分布于长江流域以南的亚热带中、低山地区,我国11 省84 县均有鹅掌楸天然林的分布,由于其材型通直高大,屡遭滥伐,主要分布区鹅掌楸数量已逐渐减少,且因其繁殖特性导致天然更新困难[8-9],所以其种群处于濒危状态。目前,国内对鹅掌楸的研究较多,且主要集中在其遗传特性[10]、基因与组织表达[11-12],繁殖与栽培[13-14]等方面,旨在解决鹅掌楸繁育问题,而关于现存鹅掌楸天然林群落物种组成与空间结构特征的研究甚少。本研究以鹅掌楸天然林为研究对象,分析鹅掌楸林木个体在群落中的优势度,采用林分空间结构一元分布、二元分布及四元分布分析方法,量化林分结构指标,客观评价鹅掌楸天然林空间结构特征,为实现以鹅掌楸为目的树种的可持续经营提供理论依据。
1 研究区概况
研究区位于湖北恩施土家族苗族自治州鹤峰县(109°45′~110°38′ E,29°38′~30°14′ N)境内。该区属亚热带大陆性季风湿润气候,四季分明,降水充沛,年平均降水量在1 336.9~2 189.5 mm 之间,其中43%~45% 的降水集中在夏季。年平均气温9.8~15.4℃,年日照时数1 253~1 342 h,无霜期200~266 d。海拔194.6~2 095.6 m,平均海拔1 147 m,高差明显,达1 901 m。从高海拔到低海拔主要土壤分布依次为黄红壤、黄壤与山地棕黄壤。区域内植被类型丰富,群落类型多样,且垂直分布明显,海拔800 m 以下低山区主要分布中亚热带常绿阔叶、针叶混交林,海拔800~1 500 m 的中高山和高山地区主要分布有北亚热带常绿阔叶、落叶、针叶混交林,暖温带灌木草甸植被主要分布于海拔1 500~2 095 m 的高山区。
2 研究方法
2.1 样地设置与调查
于2019 年7 月在鹤峰县中营镇白水沟村和五里乡下六峰村鹅掌楸天然群落分布区,使用罗盘仪布设40 个20 m × 20 m 调查样地,每个样地间隔距离至少大于20 m,其中白水沟村19 个,下六峰村21 个。样地建立后,使用不锈钢钢管在样地中心及四角进行埋桩标记,并记录样地生境概况:经纬度、海拔、坡度、坡向、坡位、土壤类型、受干扰程度及因素等。每个样地分为4 个10 m × 10 m 的样方,以样方为调查单元,对其中胸径 ≥ 5 cm 的乔木进行每木检尺和坐标定位,测量其胸径、树高、冠幅(取平均值),同时调查样方内灌木和草本物种组成、基径、高度及多度,层间植物物种组成等。
2.2 数据分析与处理
(1)乔木树种重要值计算:
重要值 =(相对多度 + 相对频度 + 相对显著度)/3
(2)空间结构特征指标:用以下4 个量化指标对林分空间结构特征进行描述。
角尺度:对林分空间分布格局的描述,指参照树i与4 株最近相邻木j之间所构成的夹角α小于标准角α0(α0= 72°)的个数占所形成4 个角的比例[15-16],其表达式为:

式中:Wi为角尺度,Zij为离散型变量,当第j个α角小于标准角α0时,Zij= 1;反之,则Zij= 0。
大小比数:对林木个体大小分化程度的描述,指与参照树i最近的四株相邻木j中大于参照树的株数占相邻木总株数的比例[17],其表达式为:

式中:Ui为大小比数,Kij为离散型变量,当相邻木大于参照树时,Kij= 1;反之,则Kij= 0。
混交度:对树种空间隔离程度的描述,指相邻木j中与参照树不属于同一树种的林木株数占相邻木总株数的比例[3]。其表达式为:

式中:Mi为大小比数,Vij为离散型变量,当相邻木与参照树不属于同一树种时,Vij= 1;反之,则Vij= 0。
密集度:对林木密集程度的描述,指4 株最近相邻木中与参照树树冠投影相连接的林木株数占相邻木总株数的比例[18]。其表达式为:
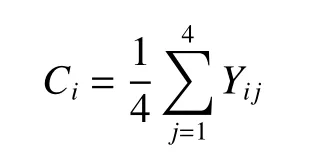
式中:Ci为密集度,Yij为离散型变量,当相邻木与参照树树冠投影相重叠时,Yij= 1;反之,则Yij= 0。
利用林分空间结构分析软件Winkelmass 与R3.5.1[19]进行分析,采用Microsoft Excel 对数据进行处理并用Origin2017 作图。
3 结果与分析
3.1 鹅掌楸天然林群落物种组成及主要乔木树种数量特征
鹅掌楸天然林群落中物种总计251 种,其中乔木88 种、灌木61 种、草本77 种、木质藤本25 种,隶属于76 科172 属。物种数较多的科主要有:蔷薇科(Rosaceae)12属27种、忍冬科(Caprifoliaceae)3 属14 种、樟科(Lauraceae)5 属13 种、菊科(Compositae)9 属13 种、百合科(Liliaceae)7 属11 种、禾 本 科(Gramineae)7 属8 种、豆科(Leguminosae)7 属8 种,等。在172 个属中,悬钩子属(Rubus)含有最多物种数,为14 种,荚蒾属(Viburnum)次之,为11 种,且76.16%的属只含有1 个种,群落物种丰富度较高。

图1 各树种的重要值(前20 种)Fig. 1 The important values of different tree species(The first 20 kinds of)
为了全面反映群落中主要乔木树种的数量特征,分析各乔木树种在林分中的地位和影响,计算各树种的重要值,重要值排名前20 的乔木树种,如图1 所示。由图可知,重要值位于第1 位的鹅掌楸为14.38%,比第2 位的杉木(Cunninghamia lanceolata(Lamb.)Hook.)高1.99%,这表明鹅掌楸为该群落的优势种群,但优势不明显。其它树种如枹栎(Quercus serrateMurray)、川陕鹅耳枥(Carpinus fargesianaH. Winkl.)、 四 照 花(Cornus kousavarchinensisOsborn)的重要值也较高,都达到6%及以上,因此该区鹅掌楸天然群落主要为鹅掌楸 + 杉木-枹栎、川陕鹅耳枥、四照花群落。柳杉(Cryptomeria japonicasinensisMiquel)、多脉青冈(Cyclobalanopsis multinervisCheng et T.Hong)、大叶杨(Populus lasiocarpaOliv.)等为群落的主要伴生种,作为林分中不可或缺的一部分,对提高林分稳定性具有重要作用。
3.2 林分空间结构一元分布特征
通过计算林木4 个结构参数(W、U、M和C)各自的不同取值等级(0.00、0.25、0.50、0.75、1.00)相对频率获得空间结构参数的一元分布(图2)。由图2 可知,鹅掌楸天然林中林木4 个结构参数呈现不同的频度变化。群落中54.29%林木以随机分布为主,以不均匀分布次之(20.45%)。林木角尺度均值为0.566,在最适经营范围[0.475,0.517]之外且大于0.517,林分整体呈现轻微团状分布。不同大小比数等级相对频率在4 个结构参数中相对变化最小,都为20%左右,均值为0.508,即不同优势程度的林木数量较为接近,表明林分生长整体处于中庸状态。林木隔离程度在不同频度的取值有明显差异。多数林木个体属于强度混交与极强度混交,总计达到71.10%,中度混交与低度混交次之,零度混交所占比例最少。整体林木混交度均值为0.742,说明林分内树种单种聚集的情况极少,整体混交良好。林分密集度均值为0.865,其中很密集林木最多,达69.02%,接下来依次是比较密集和中等密集的林木,处于稀疏和很稀疏的林木相对较少,表明其林分内林木整体较密集,林木间对营养空间的竞争较大。
3.3 林分空间结构二元分布特征
对鹅掌楸天然林空间结构参数的二元分布进行分析,即将林分空间结构参数进行两两组合(WU、W-M、W-C、U-M、U-C和M-C),任一结构参数与另一结构参数的可能取值(0.00、0.25、0.50、0.75、1.00)交叉分类,计算不同结构参数不同取值组合的相对频率。在鹅掌楸天然林中,随着角尺度的增大(Wi= 0.00→1.00),同一大小比数的林木株数呈现先增加后减少的趋势,在Wi=0.50 时达到峰值;且同一角尺度值下不同大小比数的林木株数变化差异较小,表明相同分布格局的林木在群落中生长的优劣势程度差异较小(图3a)。同一混交度的林木株数随着角尺度的增大同样呈现先增加后减少的趋势,并在Wi= 0.50 时取得最大值,而同一角尺度时随着混交度的增大,林木株数逐渐增加,且Wi= 0.50,Mi= 1.00 时的林木株数最多,占群落林木总数的23.57%,表明鹅掌楸天然林中以随机分布且极强度混交的林分结构单元最常见(图3b)。W-C二元分布与W-M类似,处于很密集状态的林木占比达69.02%,其中处于随机分布、很密集的林木个体占比最大,达37.56%,说明鹅掌楸天然林中随机分布的林木所处小环境树冠紧密相连,林木间的竞争激烈(图3c)。图3d显示同一大小比数下,随着混交度等级的增大,林木株数逐渐增加,而同一混交度时不同大小比的林木株数相差不大;在大小比数一定时,密集程度越大的林木个体株数越多,且林分中密集度等级为很密集且处于绝对优势的林木占83.07%,比较密集且处于优势的林木的占71.9%(图3e),表明鹅掌楸天然林中占优势的林木周围林冠密集,竞争激烈;林分中混交度与密集度均随各自取值的增大而呈递增趋势,其中极强度混交且很密集的林木个体最多,占比达到了28.73%(图3f)。
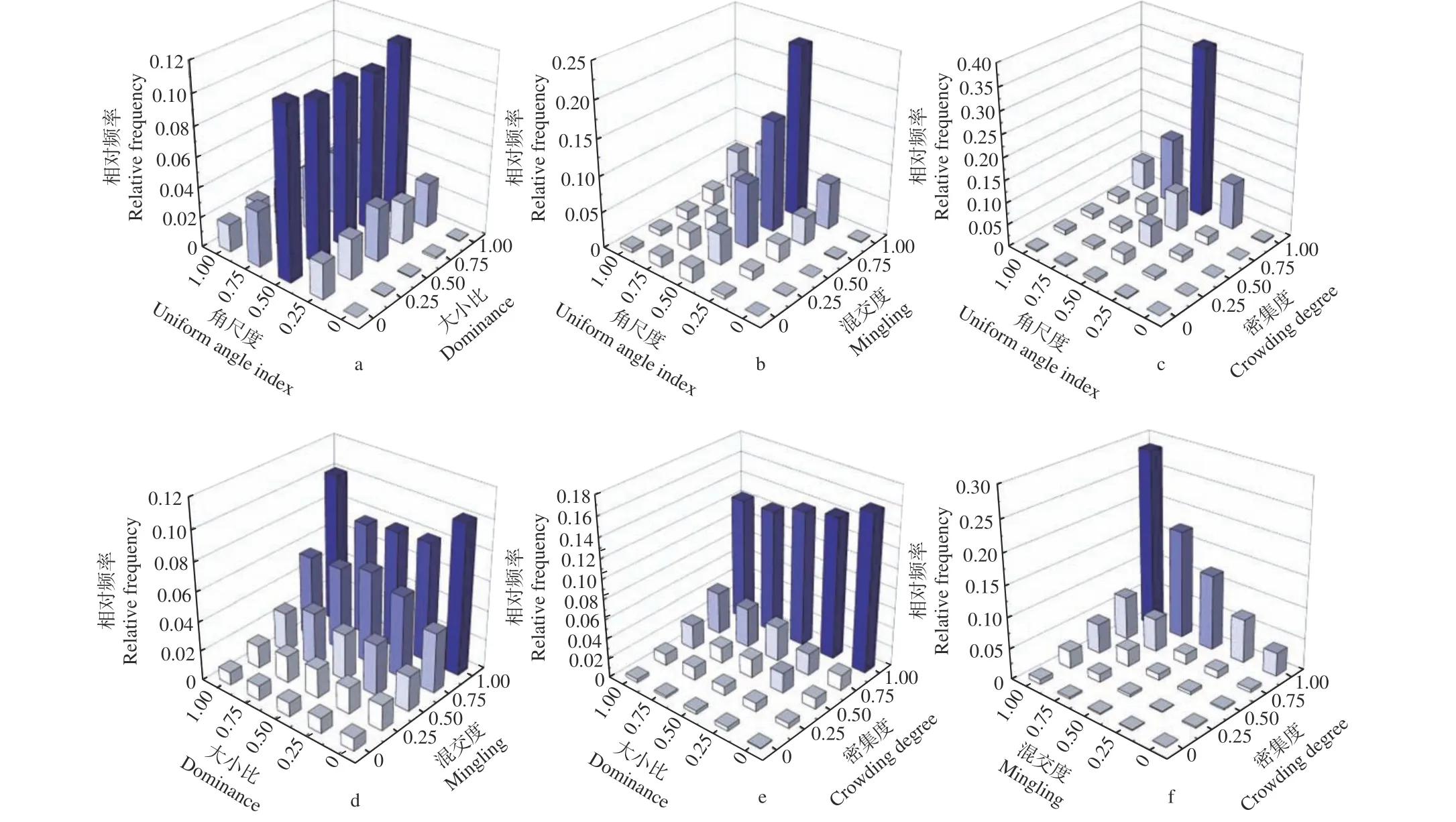
图3 空间结构特征二元分布Fig. 3 The bivariate distribution of stand structure characteristics
3.4 林分空间结构四元分布特征
通过将两个结构参数的可能取值进行交叉分类组合作为X轴,另外两个结构参数的可能取值进行交叉分类组合作为Y轴,各结构参数相互组合的相对频率为Z轴,计算组合后的625 种不同参数不同取值等级组合的相对频率,得出鹅掌楸天然林林分结构四元分布[20]。如图4 所示,在鹅掌楸天然林中结构参数为Ci= 1,Mi= 1,Wi= 0.5,Ui= 0→1的林木个体所占比例较大,占总株数的15.9%,其中处于随机分布且占绝对优势的林木个体(周围都为不同种且树冠很密集相连)最多,占总株数的4.9%;其次为随机分布,很密集且处于极强度混交和绝对劣势状态的林木个体,占总株数的3.25%;再者为随机分布,很密集且处于极强度混交的中庸林木,占总株数的2.82%。

图4 林分结构特征四元分布Fig. 4 The quadrivariate distribution of stand structure characteristics
4 讨论
4.1 群落物种组成与主要乔木树种数量特征
研究区鹅掌楸天然林群落中群落分层明显,分为乔木层、灌木层、草本层与木质藤本4 个层次,整体物种组成与姚兰等对鄂西南常绿落叶阔叶混交林物种组成相似[21]。林分中乔木树种丰富,但大部分树种数量较少,44.3%的树种在整个研究区内只有1~2 株,这也是导致群落整体混交度较高的原因之一[22];其中鹅掌楸、杉木、枹栎、川陕鹅耳枥占据优势,累计重要值达到34.75%;与其他学者[23-24]对鹅掌楸天然林的研究相比,研究区内物种丰富度相对较高,而林分内其他优势树种因地理条件不同具有明显差异。鹅掌楸树种重要值最大但优势不是很明显,这与孤生殖现象导致的自然条件下鹅掌楸种子萌发率低,以及林分中幼树幼苗极少息息相关,且鹅掌楸多数以大径级形式存在,DBH≥20 cm 个体占鹅掌楸总株数的54.3%,年龄结构呈现老龄化,因此进行适当的人为干预以保持鹅掌楸在群落中的地位对其种群繁衍是十分必要的。
4.2 林分结构一元分布特征
一元分布是描述林分某方面结构特征在不同等级的频率分布[20]。鹅掌楸天然林林分角尺度W=0.566,从林分中不同树种的空间结构单元角度分析,以杉木、川陕鹅耳枥、柳杉、鹅掌楸、四照花、枹栎、灯台树为参照树的结构单元平均角尺度与整个林分平均角尺度相似,且以上树种为该群落的主要优势种。该结果表明同种聚集分布可能是导致此种空间分布格局的原因,因此以这几个树种作为参照树的林木分布格局在很大程度上决定了整个天然群落的林分林木分布格局,且杉木、柳杉、四照花的同种聚集现象较为突出,在今后的森林经营过程中可重点调整这3 个树种的空间结构,以加快群落由团状分布向随机分布转变的进程。林分整体呈现高度混交状态,且样地中优势树种的混交度处于极强度混交与强度混交之间的频率很高,说明所处林分树种隔离程度较高,群落向稳定的方向发展[25]。林分整体生长处于中庸状态,且林分中鹅掌楸大小比数为0 和0.25 的频率达到了73.1%,大小比数为0 的个体数更是达到了48.9%,表明大部分鹅掌楸在整个林分结构单元中处于优势地位。林分整体密集度较高,林木树冠较紧密,相互之间竞争激烈[26],可根据不同的森林经营目的进行调整,去除部分林木,降低竞争势,增加林下光照。
4.3 林分结构二元分布特征
二元分布采用林分结构参数两两组合的方法来同时描述两方面的林分结构特征及其之间的相关性,具体且直观地表现出了林分空间结构状态及结构参数之间的相互关系[27]。鹅掌楸天然林林分二元分布中,角尺度对大小比数、混交度与密集度均有一定影响,呈现随机分布的林分其混交程度与密集程度相对频率均达到最高,而同一角尺度值,不同大小比数林木个体数量并无明显差异,说明林木空间分布类型并不是决定林木优劣态势的关键因素,这与毛沂新等[28]的研究结果相似,可见合理补植与采伐林木改变林分空间分布格局可有效改善林分混交状态与郁闭度。在大小比-混交度二元分布中,在同一大小比系列上,混交度与密集度相对频率逐渐升高,且同一混交度与密集度系列上,大小比数的相对频率差距较小,与张岗岗等[29]的研究结果相似,即相同密集程度、相同混交程度的林木在林分中等比例的处于不同优劣程度,且处于绝对劣势且弱度混交的林木占8.7%,在森林经营过程中,这部分林木可作为首要采伐木。在混交度-密集度二元分布中,林木混交状态与林分密集程度呈现相互促进的状态,且处于极强度混交与非常密集的林木相对频率最高,而过高的林分郁闭度不利于林下幼苗更新,进行合理调整,改善林下光照,补植珍贵乡土树种,提高物种多样性与林分结构稳定性的同时增加森林多重效益[30]。
4.4 林分结构四元分布特征
目前,对林分空间结构四元分布的研究较少,四元分布结合了角尺度、大小比、混交度和密集度4 个林分结构参数,包含了625 种林分结构参数组合。本研究得出,在鹅掌楸天然林中,处于绝对优势,4 株最近相邻木皆与参照树不同种且相邻木随机的密集分布于参照树周围的林木个体所占比例最大,可见,林分空间结构四元分布可对林分进行全方位的、整体性的精准描述[19],这对林分整体调控具有极强的指导意义。在森林经营活动中,可根据林分现状与经营目的要求,制定以角尺度、混交度、大小比数和密集度为指标体系的林分优化方案,保留优势木,促进林分结构向随机分布、强度混交转变,筛选出林分结构不合理的林木加以调整,提高林分质量,加快现实林分向理想林分转化的速度,优化经营,促进森林经营目标的实现[31]。
5 结论
针对鄂西南鹅掌楸天然林物种组成与林分结构的研究发现,鹅掌楸天然林群落物种丰富,分层明显。群落中鹅掌楸重要值最大,且该群落主要为鹅掌楸 + 杉木-枹栎、川陕鹅耳枥、四照花群落。林分整体呈现轻微团状分布,但随着自然演替发展将逐渐呈现随机分布,林木混交度较高,且呈现密集分布,林分生长中庸。天然林中以随机分布、极强度混交的林分结构单元最常见,相同分布格局的林木在群落中生长的优劣势程度差异变化较小,占优势的林木周围林冠密集,竞争激烈。群落中以优势木为目标树构成的随机、密集、强度混交的结构单元分布最多,因此选择性地伐除非目标树、且处于绝对劣势且弱度混交的林木个体,可有效调整林分结构,改善林内光照条件,促进天然更新,提高森林生态系统稳定性,增强森林多功能效益。
猜你喜欢
小星星·阅读100分(低年级)(2022年9期)2022-09-23 07:30:52
现代园艺(2021年12期)2021-12-07 03:59:22
鸭绿江(2021年17期)2021-10-13 07:06:16
临床检验杂志(电子版)(2020年1期)2020-04-03 04:00:16
新农民(2019年9期)2019-02-19 14:19:13
金色少年(奇趣科普)(2017年10期)2017-12-14 03:22:18
三明学院学报(2016年2期)2016-10-14 08:43:29
学苑创造·B版(2015年12期)2016-06-23 09:36:36
福建农业科技(2015年3期)2015-02-27 10:20:44
