
西藏工布自然保护区高山松林土壤种子库空间分布动态特征
2021-04-10 05:23:30李照青李江荣郑维列
林业科学研究 2021年2期
高 郯,李照青,卢 杰*,李江荣,郑维列
(1. 西藏农牧学院高原生态研究所,西藏 林芝 860000;2. 西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3. 西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000;4. 西藏绿景生态工程咨询有限公司,西藏 拉萨 850000)
种子是植物体进行有性繁殖,延续后代的重要阶段,土壤种子库是潜在种群阶段,是植被种群生活史中的一个重要阶段,能够反映过去、现在的种群结构特征及未来的演变趋势,并对种群的繁殖更新、物种多样性的维持、植被的演替和恢复等发挥重要作用[1-6]。土壤种子库的研究始于达尔文用幼苗数代表种子数[7],之后有关土壤种子库的研究涉及农田、草地、森林、湿地等生态系统的种子生产、散布、休眠及萌发特征、种子及种子库寿命、种子库与群落演替关系等诸多方面,并结合植物学、生态学、遗传学以及农学等进行研究,目前已成为恢复生态学的研究热点[8-11]。我国的土壤种子库研究工作始于20 世纪80 年代末,研究内容首先在林地,其次在草地、沙地和湿地生态系统上开展的一系列关于土壤种子库种类、多样性与地被植物间关系的研究,现阶段的研究内容多集中于“不同恢复阶段的土壤种子库特征及环境因子对土壤种子库时空分布特征的影响”上,旨在了解植被退化机制,为植被的恢复提供理论支撑[12-17]。
高山松(Pinus densataMast.)为我国特有种植物,是西藏工布自然保护区的主要建群种之一,在涵养水源、保持水土平衡及保护生物多样性等方面具有重要生态价值;同时因其松脂含量丰富达到国家Ⅰ级甚至特级标准,高山松具有较高的经济价值[18-19]。近年来,由于人类活动的破坏及严重病虫害的威胁,高山松种群发生严重的退化趋势[20]。高山松的研究多集中在生物量监测、种群结构等方面[21-24],较少涉及土壤种子库方面[25-26],尤其是土壤种子库动态变化及影响因素分析;而土壤种子库特征对高山松的更新、恢复又具有重要意义。故本研究以工布自然保护区高山松林分布较典型的3 个样点为研究对象,分析不同物候期高山松林土壤种子库的动态特征及影响因素,旨在阐明高山松林土壤种子库退化机制,为高原植被的保护、恢复及合理利用提供理论依据。
1 研究区概况
研究区位于林芝境内的工布自然保护区(92°09′~98°47′ E、26°52′~30°40′ N),是藏东南保存较完好的原生林区之一,属湿润山地暖温带和半湿润山地温带气候,受印度洋暖湿季风影响,干湿季分明,冬季干燥,夏季多雨。据西藏林芝高山森林生态系统国家野外科学观测研究站2017 年至2019 年气象数据可知,该地区年均气温3.9℃,年均相对湿度78.6%,年均降水量767.1 mm,6—9月为雨季,占全年降水的66.2%,全年日照1 418.8 h。
该区内海拔高差大,植物种类丰富,乔木主要有以急尖长苞冷杉(Abies georgeivar.smithii(Viguie et Gaussen) Cheng et L)和林芝云杉(Picea likiangensisvar.linzhiensisCheng et L.K.Fu)为主的暗针叶林、以高山松为主的亮针叶林、以川滇高山栎(Quercus aquifolioidesRehd. et Wils.)为主的常绿硬阔叶林和以白桦(Betula platyphyllaSuk.)、山杨(Populus davidianaDode)为主的落叶阔叶林等[27-28]。样地林下植物灌木主要有川滇高山栎、峨眉蔷薇(Rosa omeiensisRolfe)、灰栒子(Cotoneaster acutifoliusTurcz.)、尼泊尔黄花木(Piptanthus nepalensis(Hook.) D. Don)、尖叶栒子(Cotoneaster acuminatusLindl.)、川滇野丁香(Leptodermis pilosaDiels )、 暗 红 小 檗(Berberis agricolaAhrendt)、多蕊金丝桃(Hypericum choisyanumWallich ex N. Robson)、 波 密 小 檗(Berberis gyalaicaAhrendt )、 腺 果 大 叶 蔷 薇(Rosa macrophyllavar.glanduliferaYü et Ku)、唐古特忍冬(Lonicera tanguticaMaxim.)、鸡骨柴(Elsholtzia fruticosa(D. Don) Rehd.)等;草本主要有早熟禾(Poaannua L.)、 灰 叶 香 青(Anaphalis spodiophyllaLing et Y. L. Chen)、窄叶野豌豆(Vicia sativasubsp.nigraEhrhart )、 角 盘 兰(Herminium monorchis(L.) R. Br.)、 双 参(Triplostegia glanduliferaWall. ex DC.)、高山珠蕨(Cryptogramma brunnoniana(Wall.) ex Hook. et Grev.)、粘毛鼠尾草(Salvia roborowskiMaxim.i)、西藏草莓(Fragaria nubicola(Hook. f.) Lindl.ex Lacaita)、 西 南 委 陵 菜(Potentilla lineataTreviranus)、轮叶黄精(Polygonatum verticillatum(L.) All.)、尼泊尔鸢尾(Iris decoraWall.)、金荞麦(Fagopyrum dibotrys(D. Don) Hara)等;另有藤本植物西藏铁线莲(Clematis tenuifoliaRoyle)等。林下土壤为山地棕壤。
2 研究方法
2.1 样地设置
选取高山松林分布较典型的东久乡东久桥(DJ)附近、林芝镇林芝桥(LZ)附近以及巴宜区比日神山(BR) 景区为实验样点,记录样点的海拔、坡向、坡度等信息,样点概况详见表1。在每个样点,分别在高山松林分布的山脚、山腰和山顶各3 个20 m × 30 m 的乔木样方,对样方内的乔木进行每木检尺,记录树种、高度、年龄、胸径、坐标、冠幅等信息;在每个乔木样方内采取对角线五点取样法设置5 个2 m × 2 m 的灌木样方,记录灌木的名称、多度、高度、盖度等信息;在每个灌木样方内随机设置1 个1 m × 1 m 的草本样方,记录草本植物的名称、多度、高度、盖度等信息。
2.2 野外取样
为明确不同物候期(种子散落后、种子越冬后、种子散落前)高山松林土壤种子库的分布特征及其对环境的响应,于2017 年12 月(种子散落后,Ⅰ期)、2018 年3 月(种子越冬后,Ⅱ期)和2018 年9 月(种子散落前,Ⅲ期)分别在选定的3 个样点进行3 次取样。在每个样点选择3 株独立性较好(周围5~8 m 范围内没有其他结实高山松)的高山松作为母树,共选取27 株母树。然后以母树为中心,在其东、南、西、北4 个方向上向外延伸,分别在1、3、5 m 处采样,取样面积为20 cm × 20 cm。据前期对高山松林的研究结果,发现高山松林土壤5 cm 以下极少有种子存在,因此,取样深度为地下5 cm,且分3 层取样:枯落物层、0~2 cm 土层及2~5 cm 土层,装袋后带回实验室待用。3 次共计取土壤种子库样品2 916 份。
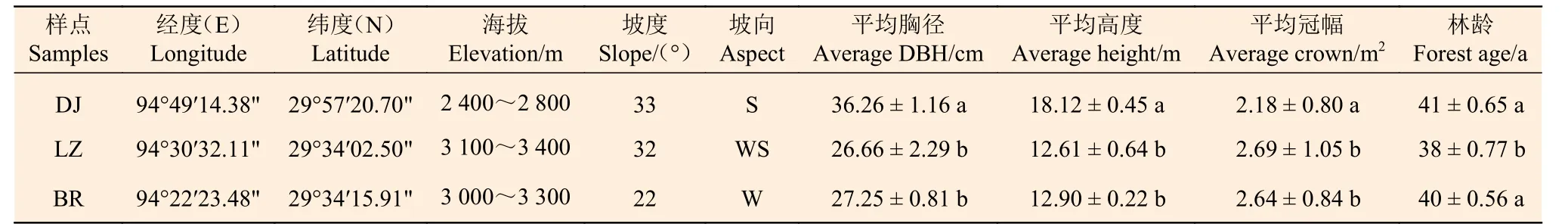
表1 高山松林土壤种子库样点基本情况Table 1 Basic overview of soil seed bank plots in P. densata forest
2.3 种子质量分类及活性鉴定
首先,对野外采集的高山松林土壤种子库样品分别过5 mm 和2 mm 的土壤筛,然后对样品进行人工分拣,挑拣出样品中的高山松种子,并根据高山松种子的外表情况,分为虫蛀种子、败育种子(种子因发育畸形而明显小于或轻于正常完好种子)、腐烂种子(外种皮明显霉变发黑的种子)及完好种子4 类统计,最后采用氯代三苯基四氮唑(TTC)染色法[29]对高山松完好种子进行种子活性鉴定。
2.4 数据处理
高山松林土壤种子库种子密度(粒·m-2)用土壤种子库中的种子数量除以对应采样面积来表示。采用Excel 2010 对数据进行统计整理,并运用SPSS 17.0 对高山松林不同方向、不同距离、不同垂直深度的高山松种子密度进行单因素方差(oneway ANOVA)和Duncan’s 多重比较分析。
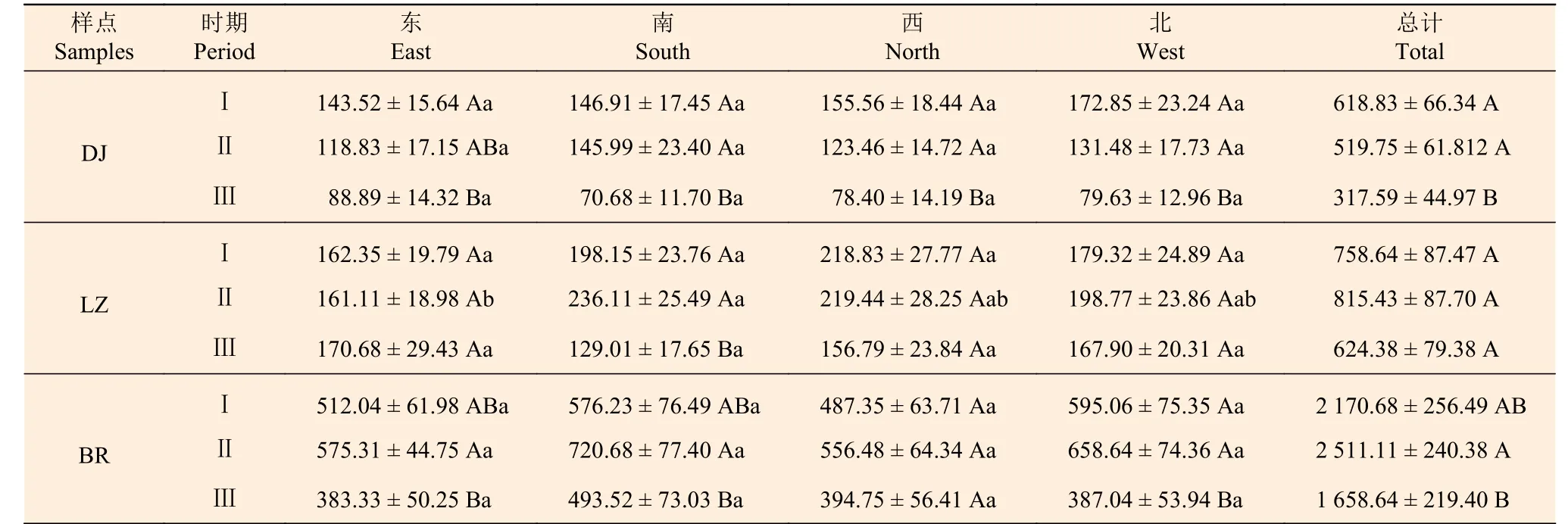
表2 不同时期高山松林各样点种子密度在水平方向上的分布特征Table 2 Horizontal distribution characteristics of seed density of plots of P. densata forest at different periods
3 结果与分析
3.1 不同时期各样点高山松林土壤种子库水平分布
不同时期高山松林各样点种子密度在母株周围水平方向上的分布特征见表2。不同时期,各样点高山松林土壤种子库在东、南、西、北4 个方位上的分布总体上差异不显著(P> 0.05),但LZ 样点第Ⅱ时期的东、南方向除外。各样点土壤种子库密度上表现为BR > LZ > DJ。DJ 样点在东向上第Ⅱ时期与第Ⅲ时期差异不显著(P> 0.05),其他各方位上均表现为Ⅰ、Ⅱ时期与第Ⅲ时期差异显著(P< 0.05);LZ 样点除南向上Ⅰ、Ⅱ时期与第Ⅲ时期差异显著外(P< 0.05),其他各方向上在3 个时期间差异均不显著(P> 0.05);BR 样点除在西向上各个时期差异均不显著(P> 0.05)外,其他3 个方向均为第Ⅱ时期与第Ⅲ时期差异显著(P< 0.05)。随着取样时间的变化,DJ 样点各时期高山松土壤种子库的种子密度呈递减趋势,而LZ 样点和BR 样点随着取样时间的改变,其各时期高山松土壤种子库的种子密度先增加后减少(LZ 样点东向除外)。
各时期高山松林土壤种子库母树周围不同尺度下的分布特征见图1。不同时期各样点距高山松母株不同距离上的土壤种子库密度均表现为1 m > 3 m > 5 m。第Ⅰ时期各样点在距母树1 m 和3 m 时差异不显著(P> 0.05),而5 m 处高山松土壤种子库数量显著低于1 、3 m(P< 0.05);第Ⅱ、Ⅲ时期各样点在距母株不同尺度下,其差异性各异,除DJ 样点在3 个尺度下均差异不显著(P>0.05)外,其他2 个样点总体上表现为1 m 处高山松种子数量显著高于5 m(P< 0.05),且LZ、BR 2 个样点1 m 处高山松种子数量分别是5 m处的1.69、1.80 倍(Ⅱ 期)和1.60、 1.63 倍(Ⅲ期)。不同时期各样点高山松土壤种子库密度均表现为BR > LZ > DJ。
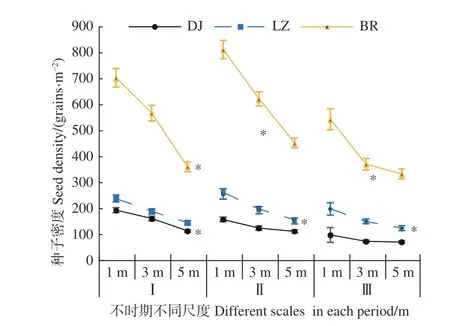
图1 不同时期高山松林各样点种子密度随尺度变化规律Fig. 1 Variation of seed density with scale in different plots of P. densata forest in different periods
3.2 不同时期各样点高山松林土壤种子库在不同土层分布格局
不同时期各样点高山松林土壤种子库种子密度在不同土层分布特征见图2。不同样点在各时期的土壤种子库分布均表现为随土壤深度的增加而减少。除第Ⅱ和第Ⅲ时期的DJ 样点在0~2 cm 土层和2~5 cm 土层差异不显著(P> 0.05)外,其他样点在各时期均表现为枯落物层显著高于0~2 cm 土层(P< 0.05),2~5 cm 土层显著低于0~2 cm 土层(P< 0.05)。在Ⅰ、Ⅱ、Ⅲ 3 个时期,BR 样点枯落物层高山松种子密度均最大,分别是0~2 cm 土层的3.61、2.47、5.90 倍。第Ⅰ时期,各土壤深度均表现为BR 采样点种子密度显著高于其他2 个采样点(P< 0.05),而DJ 和LZ 采样点之间各土层差异均不显著(P> 0.05);第Ⅱ、Ⅲ时期,各样点在2~5 cm 土层之间的差异性同第Ⅰ时期一致,但各样点在枯落物层和0~2 cm 土层均表现为BR 显著高于LZ,LZ 又显著高于DJ(P< 0.05)。在不同时期,BR 枯落物层中的种子密度分别为LZ 和DJ 的2.80、2.77、2.73和3.75、4.30、5.32 倍。

图2 不同时期高山松林各样点种子密度在不同土层分布特征Fig. 2 Different soil layers distribution characteristics of seed density in different plots of P. densata forest in different periods
3.3 不同时期高山松林土壤种子库种子密度对海拔的响应
BR 样点和LZ 样点海拔相近,LZ 样点坡度、坡向等基本地形因子与DJ 样点相似,为保证外在环境条件一致,以DJ 和LZ 样点为研究对象,分析不同采样时期高山松林土壤种子库密度对海拔梯度的响应(图3)。随海拔的升高,高山松林土壤种子库数量先减少后升高,至3 100 m 达到极大值,之后又降低,随后又略微升高。总体上,高山松林的土壤种子库种子密度随海拔的升高呈波动趋势,但从DJ 和LZ 样点分开看,各山体均表现为山腰小于山顶及山脚,即呈“V”字型。除3 100 m以上,第Ⅱ时期略高于第Ⅰ时期外,其他各海拔处均为第Ⅰ时期 > 第Ⅱ时期 > 第Ⅲ时期。

图3 各时期高山松林土壤种子库种子密度对海拔梯度响应Fig. 3 Response of seed density of P. densata forest soil seed bank to altitude gradient
3.4 不同时期各样点高山松林种子质量构成
不同时期各样点高山松林土壤种子库种子质量成分构成见图4。不同时期各样点高山松林土壤种子库主要由虫蛀种子、败育种子、腐烂种子和完好种子构成,其中,虫蛀种子和完好种子占比较大,二者共占比65%以上,最高可达85.79%;腐烂种子占比最小,并随着采样时期的变化而增大,在第Ⅲ个采样期,腐烂种子占比增加到11%,败育种子的比例亦在增加,最高可达21.64%。经不同土层分析可知:土壤种子库中具有潜在更新能力的种子即有活力种子的数量极少,仅占种子库中种子总量的1.43%,其中,73.38% 的有活力种子储存在枯落物中,且随土壤深度的增加而不断减少,2~5 cm土壤层中有活力的种子仅占该类种子总量的5.84%。
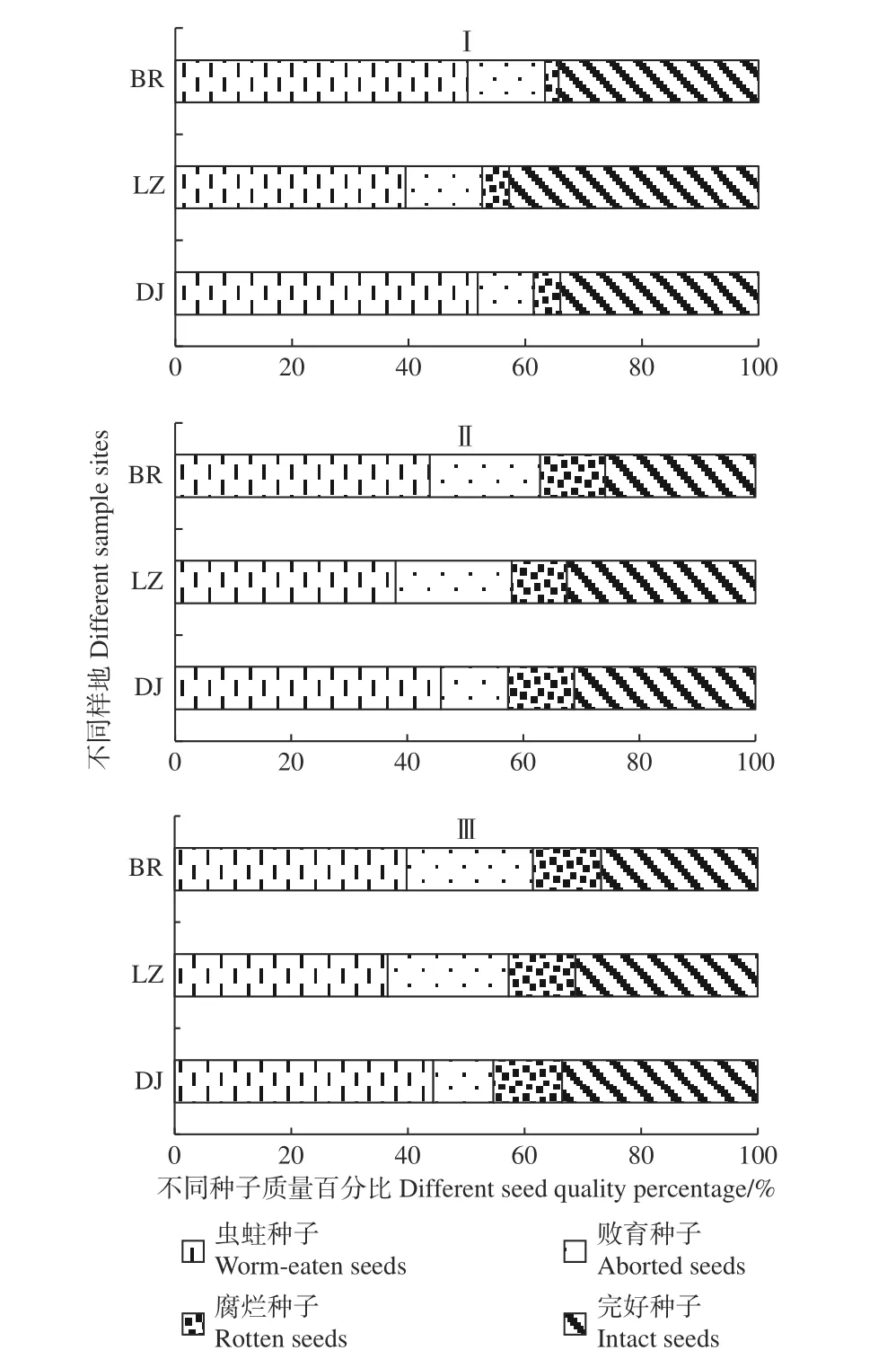
图4 不同时期高山松林土壤种子库种子质量百分比Fig. 4 Percentage of seed quality of P. densata forest soil seed bank in different periods
4 讨论
4.1 不同时期各样点高山松林土壤种子库分布动态
植物的繁殖方式分为有性繁殖和无性繁殖,其中,种子萌发是实现植物有性生殖,种群延续发展的重要和敏感阶段[1,30]。种子的分布特征受多种因素的影响而呈现不同的分布特征,如自身重力、风力、地形因素及动物的干扰等[29,31]。在本研究中,各时期不同样点高山松林土壤种子库在不同方向上的分布总体上无显著差异,这与李照青等[25]、Lu 等[26]的研究结果一致,可能是各样点坡度、坡向对其影响较小,而自身种子较小,且带有种翅,易受风力等的影响而向除坡向以外的方向飘落,使四个方向的种子密度分布相近;且林内并没有形成强风或流水对种子传播影响的外部条件,高山松林土壤种子库最终呈现为靠母株分布。随着取样时间的变化,DJ 样点各时期高山松林土壤种子库的种子密度逐渐减少,这可能与物种的物候期及环境因素有关[32]。据Lu 等[26]对高山松林的研究结果表明,高山松种子雨期为10 月初至11 月底。第1 次采样时间为12 月初,此时种子密度最大,经过一个冬季后,万物复苏,动物活动频繁,种子数量减少,随着生长季的到来,较多种子萌发,种子数量降低,这与卢彦磊等[30]对各坡向锐齿栎土壤种子库动态特征研究结果一致,与Russi 等[33]和Mayor等[34]最大值出现在种子散落期,最小值出现在种子散落之前相似。LZ 样点和BR 样点随着取样时间的改变,其各时期高山松林土壤种子库的种子密度先增加后减少,即第2 次采样期,高山松土壤种子库密度最大,这可能存在2 类持久性土壤种子库(PSSB)和2 类短暂性土壤种子库(TSSB),与植物的植冠种子库特征有关[35];也可能是外界环境的变化引起植物结实的周期性改变[36]。
4.2 不同时期高山松林土壤种子库种子密度垂直分布及对海拔的响应
种子成熟后,首先下落在土壤表面的枯落物层中,再根据种子自身特性存在于不同位置,高山松种子较小,长4~6 mm,宽3~4 mm,容易藏匿在枯落物层中,在地球引力、雨水冲刷、动物等的扰动下,掉落在表层土壤中,随土壤深度的增加,土壤种子库密度逐渐减少。本研究结果表明,主要集中在枯落物层和0~2 cm 土层,二者占比89%以上,这与吴敏等[37]对不同生境中栓皮栎土壤种子库垂直分布研究结果一致。
海拔是一种综合了温度、湿度、光照、水分的环境因子指标,海拔的变化,可引起植物生长发育所需的非生物因子的改变,从而对植物生长特性产生影响[15,38]。为保证外在环境条件一致,综合DJ 和LZ 两个样点,研究海拔对土壤种子库的影响。总体上,高山松林的土壤种子库种子密度随海拔的升高呈波动变化,这与黄瑞灵等[29]和郭连金等[39]对青海省拉鸡山不同植被土壤种子库随海拔变化不同,可能是高原地区辐射强,海拔越高越有利于吸收光照,从而有利于强阳性树种结实,但同时高海拔所带来的低温亦会影响种子的结实在海拔、光照、温度的综合作用下表现出波动的趋势,同时也要考虑坡位等因素。从DJ 和LZ 样点分开看,各山体均表现为山腰 < 山顶及山脚,即呈“V”字型,且下坡位种子密度高于上坡位和中坡位,这可能是下坡位土壤养分等易聚集,植被发育较好,有性繁殖能力较强,也可能与雨水冲刷使种子聚集有关;而上坡位种子密度多于中坡位,可能是上坡位光照良好,更有利于强阳性树种的生长,需要进一步研究。
4.3 高山松林土壤种子库质量特征
种子库组成既反映了种子质量,又反映了有性生殖能力,在种群的更新、繁衍中发挥重要的作用[37]。种子丧失活力的主要原因是动物的捕食、潮湿环境引起的霉变腐烂等[40-41]。本研究表明,虫蛀、完好类占比较大,二者共占比65%以上,最高可达85.79%,这与李照青等[25]对高山松及刘彤等[42]对红豆杉的种子质量研究结果一致。同时,随着采样时间的变化,腐烂、败育的种子数量在增多,完好的种子数量在下降,这与吴敏等[37]的研究结果相似,可能是随着天气的变暖及雨季的到来,枯落物层中的种子生存环境潮湿,易于霉变,这极大影响高山松更新。虫蛀种子数量略微下降,可能是林下草本、灌木随着生长季的变化,产生了一定数量种子,减小了动物对高山松种子捕食的压力;也可能是动物与取食植物之间的互利共生关系,有益于种子的传播而不是活力丧失,如鲁长虎[43]认为,鸦科鸟类以及啮齿类动物既是植物种子的捕食者,也是传播者,相互之间是一种比较复杂的互利共生关系,具体原因有待于进一步考究。
5 结论
各样点高山松林土壤种子库在不同时期水平分布上无明显差异,而在距母树不同距离及不同土层有明显的近母株分布效应和垂直分布特征,表明高山松种子自身特性和外界风等因素对其种子库的分布特征影响较大。海拔作为一种光照、温度的综合因子对高山松林的土壤种子库分布特征也有一定的影响,而坡度、坡向等无明显效应。随着时间的变化,腐烂种子率有所增加,完好种子百分比降低,有活力的种子数量也较少,表明受工布自然保护区高湿多雨的气候特征影响,高山松种子更新能力较弱;同时,林分特征、灌草分布等均会对种子的分布及质量产生影响,有待后续研究。
猜你喜欢
Plasma Science and Technology(2022年9期)2022-08-29 00:43:22
湖北植保(2022年4期)2022-08-23 10:51:52
鸭绿江(2021年35期)2021-11-11 15:25:02
土壤(2021年1期)2021-03-23 07:29:06
军事文摘(2018年24期)2018-12-26 00:57:56
土壤学报(2017年5期)2017-11-01 09:21:27
连环画报(2016年10期)2016-12-16 05:13:34
心潮诗词评论(2016年1期)2016-12-01 05:48:31
乡村地理(2016年2期)2016-06-15 20:29:26
西部广播电视(2015年7期)2016-01-16 03:45:18
