
蔷薇属月季组植物的花粉形态学研究*
2021-04-07 00:59程璧瑄周利君蔡珩瑜潘会堂张启翔
云南农业大学学报(自然科学) 2021年2期
程璧瑄,于 超 ,周利君,蔡珩瑜,罗 乐,潘会堂,张启翔
(1.花卉种质创新与分子育种北京市重点实验室,北京 100083;2.国家花卉工程技术研究中心,北京 100083;3.城乡生态环境北京实验室,北京 100083;4.北京林业大学 园林学院,北京 100083)
蔷薇属(Rosaspp.)是世界最著名的观赏植物类群之一。全属约200 种植物,产自中国的蔷薇属植物有82 种[1]。月季组的物种是蔷薇属下的重要研究材料。月季组中包含了月季花(R.chinensis)、香水月季(R.odorata)和亮叶月季(R.lucidissima),在蔷薇属中具有极高的育种价值。香水月季自然分布于中国云南[1],其对现代月季的形成起到了关键作用。目前对于香水月季的记载主要依据《中国植物志》,即3 个变种:大花香水月季(R.odoratavar.gigantea)、橘黄香水月季(R.odoratavar.pseudlindica)和粉红香水月季(R.odoratavar.erubescens)[1]。然而野外调查发现:香水月季野生居群的表型变异极为丰富,且缺乏深入研究。
花粉具有个体小、质量轻、产量大和易于保存等特点[2]。花粉的形态特征相对稳定,是研究植物分类、系统演化的重要依据之一,在观赏植物分类和亲缘关系研究中起到重要的作用[3-7],也是探讨种和品种演化的重要依据[8-13]。
国内外对于蔷薇属的孢粉学有一定的研究[14-16]。冯立国等[17]研究了野生玫瑰及玫瑰品种的花粉形态及其演化趋势;白锦荣[14]对蔷薇野生种与中国传统月季品种花粉形态做过初步研究,依据花粉外壁纹饰的差异对材料进行聚类分析,根据孔穴密度可以区分表型相似的月季品种月月粉和月月红,并探讨了中国传统月季的品种起源和演化;罗乐等[18]对29 个蔷薇属种类的花粉进行分析,探讨了蔷薇属的亚属及组间的亲缘关系,为蔷薇属植物系统进化研究提供了孢粉学依据。
本研究对蔷薇属月季组的香水月季和月季花的25 份材料进行花粉形态学研究,包括野生资源和品种资源,探讨物种之间的亲缘关系,为相关的育种工作提供依据。
1 材料与方法
1.1 研究材料
选取的21 份香水月季材料于2012—2017 年间采自中国云南省的18 个地区,采样地及采样地经纬度、海拔如表1 所示。同时以中国四川省的2 份单瓣月季花(R.chinensisvar.spontanea)(13 号和22 号)、采自重庆的野生月季花(24 号)以及月月粉(R.chinensis‘Old Blush’)(25 号)为对照材料进行比较分析。

表1 研究材料采样地地理信息Tab.1 Geographic location of selected sites of the materials
1.2 研究方法
取尚未开放的花蕾,剥离花药后放在干燥通风的室内自然风干。散粉后装入有硅胶颗粒的离心管中并用棉花封口。试验前用烘干机在60 ℃条件下烘干至花粉达到绝干状态,避免因花粉粒中的水分影响拍摄[19]。真空镀膜后用扫描电子显微镜(日立S-3400N II)和场发射扫描电子显微镜(SU8010)扫描照相。分别于200~500× (群体)、2 000×、3 000× (个体、赤道面观、极面观)和8 000× (外壁纹饰)下观察并拍照。试验数据测量分析所用软件为AutoCAD 和SPSS 21.0,随机选取照片中状态良好的花粉,用AutoCAD 测量极轴长(P)、赤道轴长(E)、孔穴直径、条嵴间距、条嵴宽度、孔穴密度、条嵴密度、萌发沟长及萌发沟宽,每份材料的每种性状测量30 个样本数据并取平均值。另外观察并记录各花粉的赤道面观、极面观以及外壁纹饰的类型。
根据观测记录结果,选取12 个性状,其中数量性状10 个,定性指标2 个,利用SPSS 21.0软件将所获得的数据进行指标化后,使用最远邻元素法对其进行聚类分析,获得孢粉形态学性状聚类图,并对其结果进行分析。结果与分析中的专业术语均参照文献[20]。
2 结果与分析
2.1 花粉外形
由扫描电子显微镜观察结果(图1~5)可知:研究观察的全部25 份供试材料的花粉均为单粒、等极和辐射对称。根据ERDTMAN[20]提出的NPC 分类系统,所有供试材料的花粉均属于N3P4C5型花粉。

图1 1~5 号供试材料花粉形态Fig.1 The pollen morphologies of No.1-5 materials of R. sect.chinenses

图2 6~10 号供试材料花粉形态Fig.2 The pollen morphologies of No.6-10 materials of R. sect.chinenses
由表2 所示:研究观察的大部分材料的花粉为超长球形(P/E值>2),仅有少数几份香水月季资源花粉为长球形(P/E值≤2)。所有花粉的极面观均为具三孔沟的三裂圆形;赤道面呈椭圆形或矩圆形,可见1~2 条萌发沟。萌发沟两端几乎达到两极,沟长范围在31.09~38.92 μm,而沟宽差异较大,范围在0.24~1.39 μm。具体花粉形态特征见表2。
2.2 花粉大小
花粉粒大小中等,极轴长(P)范围在36.06~42.81 μm 之间,赤道轴长(E)范围在16.39~20.07 μm 之间,P/E值为1.98~2.35。其中最大花粉粒来自14 号材料,P×E的平均值为42.81 μm×19.83 μm;8 号香水月季花粉的P×E平均值最小,为36.29 μm×16.66 μm。
2.3 外壁纹饰类型
供试材料的花粉外壁纹饰主要为孔穴—条纹型,而不同种类的孔穴和条纹具不同特点,可进一步细分为不同的类型。
2.3.1 条纹
条纹纹饰的主要区别在于其条嵴宽度和间距,条纹的深浅、密度以及条嵴走向。从表2 可以看出:花粉的条嵴宽度在0.14~0.24 μm,间距则在0.09~0.28 μm。其中条嵴密度最大的为3 号香水月季,密度最小的为18 号香水月季。条嵴走向也是判断花粉形态差异的一个重要因素,结果分析大致可分为三类:第1 类是条纹从两极起始,几乎与两极平行,如20 和23 号材料(图5);第2 类是条纹几乎与两极垂直,13 和14 号材料属于此类(图3);第3 类是条纹在两极处与其近似垂直,在赤道面处则与两极几乎平行,此种类型的花粉最多,涵盖了大部分供试材料。此外,本研究所测定的25 份材料的花粉均有清晰深刻的条纹和纹理,因此在条纹深度方面,供试材料间不存在较大差异。

图3 11~15 号供试材料花粉形态Fig.3 The pollen morphologies of No.11-15 materials of R. sect.chinenses
2.3.2 孔穴
孔穴的区别主要在于其孔径大小、密度、穿透力以及分布的差别上。25 份香水月季材料中,孔穴的形状多为圆形或近圆形。不同材料的孔穴穿透力具有一定的差别,可大致分为两类:一类是孔穴均较为深刻,具有一定的穿透力,此种孔穴几乎分布在花粉的各个位置;另一类是孔穴深浅不一,很多孔穴仅在外壁表面形成凹槽而无法深入,此种现象多发生在条纹细密的纹饰中,同时孔穴多分布在两极或萌发沟处。由表2 可见:不同材料的孔穴密度具有很大差异,而其中孔穴的分布也存在着不同,孔穴密度最大的17 号香水月季的孔穴遍布整个花粉粒(图4),而密度最小的12 号香水月季的孔穴多分布于萌发沟边和两极处,同时其条纹密集,其孔穴也深浅不一(图3)。
2.4 聚类分析

图4 16~20 号供试材料花粉形态Fig.4 The pollen morphologies of No.16-20 materials of R. sect.chinenses
采用最远邻元素法对孢粉学数据进行聚类分析。由图6 所示:所有材料整体被聚为4 个类群。聚类结果将2 份月季花资源(24、25 号)和大多数香水月季资源归在第I 类群,其中21 号资源与野生月季花(24 号)距离很近,15 号资源与月月粉(25 号)距离很近,此结果与课题组对于供试材料在表型水平的聚类分析一致,推测这2 份资源在遗传关系上与月季花类资源更为接近。第II 类群中包含了3 份香水月季资源,这3 份资源的花粉均为超长球形,且表型性状也十分相似,均为重瓣香水月季。在基于表型性状的聚类分析中,这3 份也被一同聚在1 个类群中且遗传距离十分相近。第Ⅲ类群中包含了4 份单瓣资源(1、2、13 和22 号),结果将2 份野生单瓣月季花资源一同聚在这一类群中。但在第Ⅲ类群中,香水月季资源与月季花资源没有完全分开。本次聚类结果将大花香水月季(3 号)和粉红香水月季(6 号)同聚为第Ⅳ类群,两者均为《中国植物志》中所记载的香水月季变种,外壁条纹走向均为复合性状,在两极处垂直于极面,越靠近赤道面处的条嵴越几乎与极面平行,结果表示两者遗传关系很近。
3 讨论
3.1 花粉演化趋势
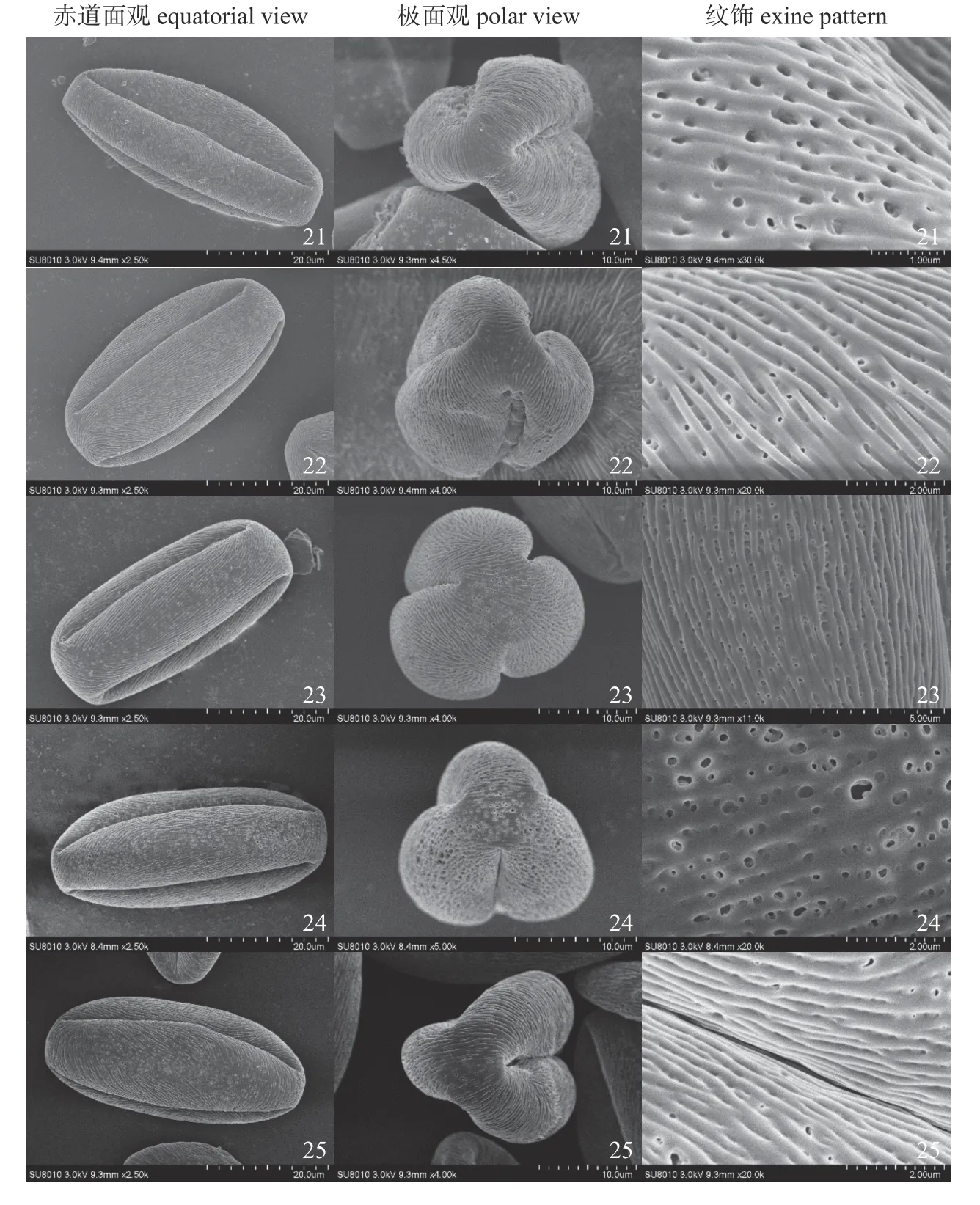
图5 21~25 号供试材料花粉形态Fig.5 The pollen morphologies of No.21-25 materials of R. sect.chinenses
ERDTMAN[20]认为:被子植物花粉外壁纹饰的演化由无结构层(光滑)向穿孔(孔穴)发展,再由穿孔继续演化成条纹类型。WALKER[21]对毛茛科类植物花粉的主要演化趋势进行分析,认为在进化上花粉形态的演变规律为由大到小、由球形到超长球形,外壁纹饰从无穿孔覆盖层到穿孔覆盖层。WODEHOUSE[22]分析花粉功能和进化的关系时指出:花粉越长,其体积与表面积之比越小,调节功能便越强,表明花粉越进化。周丽华等[23]研究认为:穴状纹饰是蔷薇科花粉中较为原始的类型,而条纹—网状和条纹状纹饰依次是比较进化的类型。因此,花粉的大小、性状和外壁纹饰上条嵴的有无、宽窄以及间距等都可以在一定层面上反映出被子植物的进化程度。LUO 等[24]对新疆疏花蔷薇的表型变异和孢粉形态进行综合分析,发现疏花蔷薇不同变种的花粉粒形状及外壁纹饰差异较大,表明花粉形态与表型性状具有一定的相关性。本研究显示:不同采集地的香水月季的花粉在条纹走向、条嵴密度、条嵴宽度和孔穴密度等方面存在差异,很可能这些细微的差异使得香水月季产生了野外丰富的花色及重瓣性等变异。对供试香水月季资源的花粉进行比较可以看出:在单瓣香水月季材料中,花粉的P/E值普遍较小,部分为长球形;而重瓣的香水月季资源花粉P/E值较大,均为超长球形。因此,单瓣较重瓣的香水月季可能更为原始,这也反映了花粉的进化程度与植物形态分类上的联系。
根据前人的研究[18]可以得知:月月粉在月季组中的遗传水平较为进化,对比月月粉的孢粉学信息可知,大花香水月季(R.odoratavar.gigantea)和粉红香水月季(R.odoratavar.erubescens)较月月粉更为进化。两者都具有超长球形进化趋势;单位面积内的条嵴和孔穴数量也更多,花粉外壁纹饰的图案更为复杂,其条纹为复合型走势,而孔穴则深浅不一,整个外壁显示从穿孔型向条纹型演化的趋势。
与蔷薇属孢粉学相关的桂味组、芹叶组和合柱组结果相比[18],本研究的月季组种类的花粉形态大部分为超长球形。前人研究中芹叶组植物的花粉普遍孔穴较少甚至没有孔穴,桂味组植物的孔穴密度也较小[18],白锦荣[14]认为芹叶组可能为蔷薇属较原始的类型,桂味组次之,合柱组和月季组均较为进化且比较相近。这与本研究结果相符,并进一步证明了香水月季是蔷薇属中一个较为进化的物种。


图6 基于12 个花粉形态性状的25 份供试材料聚类分析Fig.6 The cluster analysis of 25 materials based on 12 pollen morphology traits
3.2 遗传关系探讨
聚类结果表明:部分香水月季资源与月月粉、单瓣月季花聚在同一类群,参考表型研究结果,可将15 与21 号归为月季花类资源。在表型性状的聚类结果中,单瓣月季花与月月粉被聚在同一类群[27],但本研究结果却将两者分开。在基于SSR 分子标记和单拷贝基因的蔷薇属植物系统发育研究中,用SSR 构建的系统进化树与本研究聚类结果相似,即将大花香水月季与粉红香水月季单独聚在一起,同时将月月粉与单瓣香水月季资源分开,而基于GAPDH基因构建的系统发育树却有着相反的结果[25]。单瓣月季花被认为是月月粉品种起源的重要亲本,而香水月季也与参与了月月粉的起源[26-27]。综上可知,月月粉、单瓣月季花和香水月季之间有着复杂的遗传相关性。
MENG 等[28]在基于叶绿体基因及单拷贝核基因的分析认为,部分重瓣的香水月季资源是月季花和大花香水月季的后代。推测在本研究中,部分香水月季资源与月季花类群遗传关系很近,因此没有被完全分开。
4 结论
本研究中供试的25 份香水月季和月季花花粉均为单粒花粉,呈长球形或超长球形(P/E在1.98~2.35 之间),大小为(36.06~42.81) μm×(16.39~20.07) μm。萌发孔三孔沟,外壁纹饰均为孔穴—条纹型,覆盖层具穿孔。不同种类之间花粉的外壁纹饰在条纹走向和孔穴穿透等性状方面有所差异。聚类分析将供试材料划分为4 个类群。本研究认为:单瓣月季花是蔷薇属中较为原始的物种,而香水月季是蔷薇属中1 个较为进化的物种。蔷薇属内亲缘关系十分复杂,因此还需结合表型与分子水平的研究结果进行更为全面的分析探究。
猜你喜欢
音乐天地(音乐创作版)(2022年9期)2022-02-15
基层中医药(2020年6期)2020-09-11
小学生学习指导(低年级)(2020年6期)2020-07-25
农业工程学报(2020年11期)2020-07-22
计算机时代(2019年12期)2019-12-23
人大建设(2018年7期)2018-09-19
小天使·一年级语数英综合(2018年4期)2018-06-22
机械制造与自动化(2018年1期)2018-03-19
作文周刊·小学一年级版(2016年11期)2016-05-10
火花(2015年7期)2015-02-27
