
铅胁迫下红麻生理特性及DNA甲基化分析
2021-04-06 23:59李增强丁鑫超胡亚丽莫良玉
作物学报 2021年6期
李增强 丁鑫超 卢 海 胡亚丽 岳 娇 黄 震 莫良玉 陈 立 陈 涛 陈 鹏,*
专题
铅胁迫下红麻生理特性及DNA甲基化分析
李增强1丁鑫超1卢 海1胡亚丽1岳 娇1黄 震1莫良玉1陈 立1陈 涛2陈 鹏1,*
1广西大学农学院/ 广西高校植物遗传育种重点实验室, 广西南宁 530004;2广西壮族自治区亚热带作物研究所, 广西南宁 530004
DNA甲基化在植物响应生物和非生物胁迫中起重要作用, 但是有关铅胁迫下植物DNA甲基化水平变化的研究报道甚少。本研究以红麻P3A为材料, 采用水培法对幼苗进行不同浓度(0、200、400、600 µmol L-1) PbCl2处理, 测定幼苗农艺性状、根系ROS含量和抗氧化酶活性等变化情况; 利用甲基化敏感扩增多态性技术(methylation-sensitive amplification polymorphism, MSAP)分析600 µmol L-1铅胁迫条件下根系DNA甲基化水平变化, 回收差异甲基化片段并克隆测序, 采用qRT-PCR技术对DNA甲基化差异基因进行表达分析。结果表明, 不同浓度PbCl2胁迫均显著抑制幼苗的茎粗、根长和根表面积, 且400 µmol L-1及以上浓度PbCl2胁迫显著抑制红麻幼苗的株高和全鲜重。随着铅浓度的提高, 红麻幼苗根系的铅含量显著升高, O2܋和MDA含量显著增加, SOD活性显著升高, POD活性呈先降低后升高, CAT活性呈先升高后降低的趋势。对照及600 µmol L-1PbCl2处理下的幼苗根系DNA甲基化率分别为71.13%、62.20%, 其中全甲基化率分别为50.52%、37.80%, 半甲基化率分别为20.62%、24.40%, 即铅胁迫显著降低了红麻幼苗根系的DNA甲基化率和全甲基化率, 提高了根系的半甲基化率。qRT-PCR分析表明, 7个与抗性密切相关的DNA甲基化差异基因也存在表达量差异, 推测DNA甲基化水平变化在响应红麻铅胁迫中发挥重要作用。本结果为深入探索DNA甲基化响应植物非生物胁迫的潜在机制, 以及生产上利用红麻改良土壤铅污染提供了理论基础。
红麻; 铅胁迫; DNA甲基化; 甲基化敏感扩增多态性(MSAP); 实时荧光定量PCR (qRT-PCR); 抗氧化酶系统
土壤重金属污染严重影响农作物的产量和品质, 并且通过食物链传递严重危害人类的健康。铅是最严重的土壤污染重金属之一, 有效治理土壤中的铅污染是目前亟待解决的重大课题[1]。红麻是世界上重要的纤维作物, 其生长迅速、生物量大、对重金属耐受性强, 并且不进入食物链, 是重金属污染土壤修复的理想作物[2]。
DNA甲基化在调控植物生长发育和响应生物及非生物胁迫中起重要作用[3-4]。研究表明, 重金属胁迫下, 植物的DNA甲基化水平和模式会发生改变,这可能是植物对逆境的应急响应[5]。甲基化敏感扩增多态性(methylation sensitive amplified polymorphism, MSAP)技术是基于扩增片段长度多态性(amplified fragment length polymorphism, AFLP)技术的基础上发展起来的, 它的主要原理是采用限制性内切酶II和I对5′-CCGG-3′位点进行酶切, 由于这2种酶对该位点DNA甲基化的敏感性不同, 因此可以通过切割产生不同的片段来揭示DNA甲基化的程度, 该技术已广泛用于植物基因组DNA甲基化水平的检测[6-7]。何玲莉等[8]利用MSAP技术检测不同浓度铅胁迫对萝卜肉质根基因组DNA甲基化水平的变化发现, 其甲基化水平随着铅浓度的提高而上升, 推测可能是由于铅胁迫产生了大量的甲基自由基所致, 并且DNA甲基化水平的变化可以激活或者抑制相关基因的表达, 进而提高萝卜对铅胁迫的适应性。郭丹蒂[9]运用MSAP技术发现, 铅和镉胁迫均导致中华水韭DNA甲基化水平提高, 但与重金属的浓度无显著相关性。殷欣[10]也运用MSAP技术发现, 大豆基因组DNA甲基化水平随镉浓度的增加而逐渐提高, 并且甲基化差异基因广泛参与了植物响应重金属胁迫的过程。然而, 也有研究发现, 重金属胁迫导致基因组DNA甲基化水平降低的现象, 表明重金属胁迫下DNA甲基化水平变化和响应机制的复杂性[11]。
有关DNA甲基化响应红麻重金属胁迫的研究尚未见报道。本研究利用不同浓度PbCl2对红麻材料P3A幼苗进行胁迫处理, 研究幼苗农艺性状、ROS含量和抗氧化酶活性变化, 检测铅胁迫下基因组DNA甲基化水平变化, 并对甲基化差异片段进行克隆测序和表达水平分析, 以期为明确铅胁迫对红麻生长的影响, 揭示DNA甲基化响应红麻铅胁迫的潜在机制具有重要意义。
1 材料与方法
1.1 试验材料及处理
红麻材料P3A由广西大学农学院周瑞阳教授提供。挑选适量籽粒饱满的种子, 常温条件下用蒸馏水浸泡1 h, 经3%过氧化氢消毒10 min, 再用蒸馏水冲洗3次, 均匀地摆放在铺有2层纱布(注意定期定量添加1/4 Hoagland营养液, 始终保持纱布湿润)的塑料盒(27 cm´18 cm´9 cm)中, 放置于恒温光照培养箱(光/暗周期为10 h/14 h, 温度为白天27℃/夜晚25℃, 下同)中培养7 d后, 选取长势一致的幼苗240株, 随机分成12组, 每组20株, 移栽于12个底部含有托盘的育苗盘(21孔花卉育苗盘)中, 即为4个处理, 每个处理3次重复。向托盘中分别添加含有不同PbCl2浓度(0、200、400、600 µmol L-1)的1/4 Hoagland处理液, 注意每天定量地更换处理液。
1.2 农艺性状及生理指标的测定
胁迫处理7 d后, 全部材料用蒸馏水清洗干净并用吸水纸擦干, 分别用直尺、游标卡尺和电子天平测定各单株的株高、茎粗、全鲜重, 并利用根系扫描分析仪(EPSON EXPRESSION 10000XL)对根系进行扫描分析。之后将材料用液氮速冻后保存于-80℃冰箱, 用于后续试验。
分别采用羟胺氧化法[12]、硫代巴比妥酸法[13]测定根系的超氧阴离子(superoxide anion, O2܋)和丙二醛(malondialdehyde, MDA)含量; 分别采用氮蓝四唑法[13]、愈创木酚法[13]、紫外分光光度法[12]测定根系的超氧化物歧化酶(superoxide dismutase, SOD)、过氧化物酶(peroxidase, POD)、过氧化氢酶(catalase, CAT)活性。
1.3 甲基化敏感扩增多态性(MSAP)分析
参考Tang等[14]报道的MSAP方法并加以改良, 对部分处理(0、600 µmol L-1)的根系进行DNA甲基化水平分析。首先采用改良CTAB法[15]提取上述材料的基因组DNA, 之后分别用R I/II和R I/I 两种酶组合进行双酶切, 连接, 预扩增、选择性扩增, 6%聚丙烯酰胺凝胶电泳, 银染、显色, 拍照保存并进行MSAP多态性片段的统计。双酶切体系(3种酶均购于NEB公司)为R I 0.5μL、II或I 0.5 μL、CutSmart buffer 2 μL、模板DNA (100 ng μL-1) 4 μL、ddH2O 13 μL; 程序为37℃ 6 h, 80℃ (R I/II)或65℃ (R I/I) 20 min。连接体系为正反向引物(10 μmol L-1)各1 μL、T4DNA Ligase (350 U μL-1, TaKaRa公司) 2 μL、10×T4DNA Ligase buffer 2 μL、酶切产物14 μL; 程序为16℃ 14 h, 65℃ 20 min。预扩增和选择性扩增体系均为稀释10倍后的连接产物或预扩增产物5 μL、正反向引物(10 μmol L-1)各1 μL、2×RapidMaster Mix (南京诺唯赞生物科技有限公司) 10 μL、ddH2O 3 μL; 程序均为95℃ 3 min; 95℃ 15 s, 60℃ (以每对引物的平均m值为准) 15 s, 72℃ 30 s, 35个循环; 72℃ 5 min。接头序列、预扩增和选择性扩增引物序列见表1。
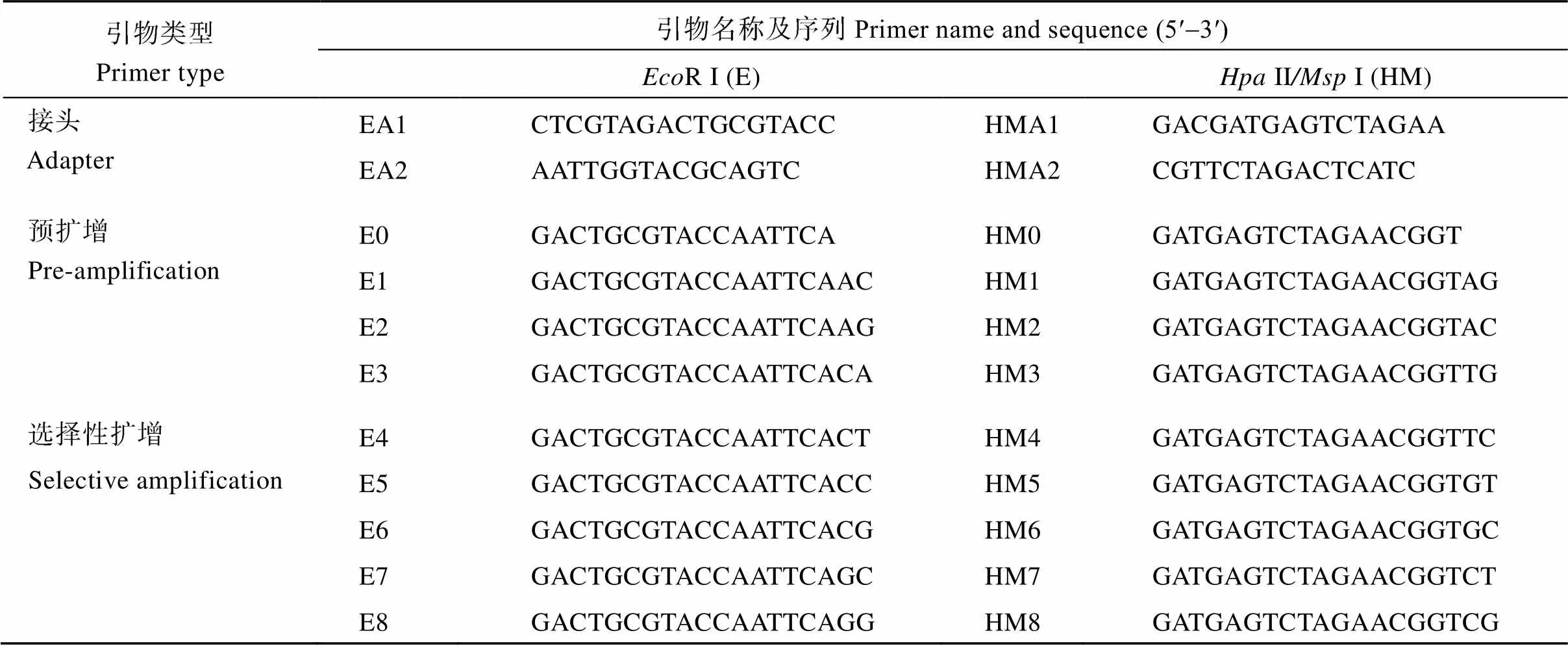
表1 接头和引物序列
MSAP多态性条带标记方法如下: 同一水平线上,R I/II和R I/I酶切都有带, 记为I型(无甲基化);R I/II酶切有带而R I/I酶切无带, 记为II型(半甲基化);R I/II酶切无带而R I/I酶切有带, 记为III型(全甲基化);R I/II和R I/I酶切都无带, 记为IV型(全甲基化)。DNA甲基化水平统计方法如下: 甲基化条带数 = II型+III型+IV型, 甲基化率 = (II型+III型+IV型)/(I型+II型+III型+IV型)´100%; 半甲基化条带数 = II型, 半甲基化率 = II型/(I型+II型+III型+IV)´100%; 全甲基化条带数 = III型+IV型, 全甲基化率 = (III型+IV型)/(I型+II型+III型+IV型)´100%[7]。
1.4 甲基化差异片段的回收、克隆和测序分析
对显色后的6%变性聚丙烯酰胺凝胶结果进行照相和MSAP多态性片段统计后, 切下观察到的甲基化差异片段于1.5 mL离心管中, 加入30 µL ddH2O, 煮沸10 mim, 用枪头将胶块捣碎, 短暂离心后取上清进行PCR扩增。体系包含上清10 μL、选择性扩增时的正反向引物(10 μmol L-1)各4 μL、2×RapidMaster Mix 50 μL、ddH2O 32 μL, PCR程序同选择性扩增。之后进行2%琼脂糖凝胶电泳及胶回收。使用北京全式金生物技术有限公司的T1 Cloning Kit进行克隆, 阳性克隆经PCR方法鉴定后, 送深圳华大基因科技服务有限公司测序。测序结果在红麻基因组数据库[16](https://bigd.big.ac.cn/gwh)及NCBI网站上进行Nucleotide BlAST同源性比对。
1.5 实时荧光定量PCR (qRT-PCR)分析
利用改良异硫氰酸胍法[17]分别提取相应根系材料(0, 600 µmol L-1)的RNA, 并进行1%琼脂糖凝胶电泳, 采用超微量紫外分光光度计检测RNA的质量和浓度, 之后使用诺唯赞反转录试剂盒(货号: R223-01)反转录成cDNA, 并以此为模板进行qRT-PCR分析。以组蛋白基因为内参基因, 采用2−ΔΔCT方法[2]计算基因的相对表达量, qRT-PCR引物序列见表2。
2 结果与分析
2.1 不同浓度PbCl2胁迫对红麻幼苗农艺性状的影响
对不同浓度PbCl2胁迫下红麻幼苗农艺性状的测定发现(表3), 低浓度(200 µmol L-1) PbCl2胁迫对幼苗的株高、全鲜重和根鲜重无显著抑制, 中浓度(400 µmol L-1)及高浓度(600 µmol L-1)胁迫下均出现显著抑制。说明低浓度PbCl2胁迫对红麻幼苗生长无明显的影响, 红麻具有较强的铅耐受性。不同浓度PbCl2胁迫均显著抑制了幼苗的茎粗、根长和根表面积。中浓度和高浓度PbCl2胁迫对幼苗生长的抑制除在株高方面呈显著性差异外, 在其他方面均无显著性差异。
2.2 不同浓度PbCl2胁迫对红麻幼苗根系铅含量的影响
由图1可知, 随着PbCl2浓度的逐渐增加, 根系中的铅含量显著升高。红麻幼苗根系的铅含量在对照条件下为3.59 mg kg-1, 低浓度(200 µmol L-1)、中浓度(400 µmol L-1)和高浓度(600 µmol L-1) PbCl2胁迫分别使根系的铅含量增加了34、138和203倍。结合对红麻幼苗农艺性状的测定结果得出, 本研究中不同浓度PbCl2胁迫虽然显著增加了幼苗根系中铅的积累, 但是并没有严重影响到红麻的生长, 说明红麻对铅胁迫具有较强的耐受性。
2.3 不同浓度PbCl2胁迫对红麻幼苗根系抗氧化系统的影响
由图2可知, 随着PbCl2胁迫浓度的升高, 红麻幼苗根系的O2܋(图2-A)和MDA含量(图2-B)、以及SOD活性(图2-C)总体上都呈逐渐上升的趋势。POD活性(图2-D)呈先下降后缓慢上升的趋势, 高浓度(600 µmol L-1) PbCl2胁迫下活性最高, 其次为对照条件和中浓度(400 µmol L-1), 低浓度(200 µmol L-1) PbCl2胁迫下活性最低。CAT活性(图2-E)呈先上升后降低的趋势, 中浓度PbCl2胁迫下活性最高, 其次为高浓度和低浓度, 对照条件下活性最低。各项指标在不同浓度PbCl2胁迫下都呈显著性差异。即不同浓度PbCl2胁迫均使红麻幼苗根系的O2܋和MDA含量升高, SOD和CAT活性上升。POD活性在高浓度胁迫下升高, 中浓度和低浓度胁迫下降低。说明铅胁迫使红麻幼苗根系发生了膜脂过氧化,但其自身的抗氧化酶活性也会相应提高, 以清除过量ROS的产生, 使铅胁迫对幼苗的伤害降到最低。

表2 实时荧光定量PCR引物序列
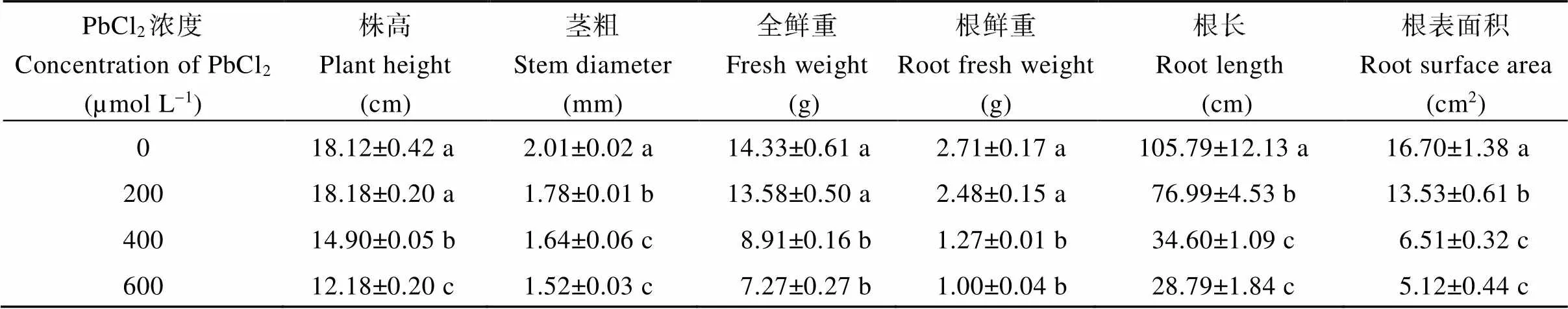
表3 不同浓度PbCl2胁迫对红麻幼苗农艺性状的影响
不同小写字母表示在0.05水平差异显著。
Values followed by different lowercase letters indicate significant difference at the 0.05 probability level.
不同小写字母表示在0.05水平差异显著。
Bars superscripted by different lowercase letters indicate significant differences at the 0.05 probability level.
不同小写字母表示在0.05水平差异显著。
Bars superscripted by different lowercase letters indicate significant differences at the 0.05 probability level.
2.4 PbCl2胁迫对红麻幼苗根系DNA甲基化水平的影响
运用MSAP技术分析了600 µmol L-1PbCl2胁迫和对照条件下红麻幼苗根系的DNA甲基化水平, 部分代表性聚丙烯酰胺凝胶电泳结果见图3, 统计结果见表4。对照条件下, 幼苗根系的DNA甲基化率为71.13%, 其中全甲基化率和半甲基化率分别为50.52%和20.62%。600 µmol L-1PbCl2胁迫下, 幼苗根系的DNA甲基化率为62.2%, 其中全甲基化率和半甲基化率分别为37.8%和24.4%。即600 µmol L-1PbCl2胁迫使幼苗根系的DNA甲基化率和全甲基化率降低, 半甲基化率升高。表明在整体水平上, 600 µmol L-1PbCl2胁迫降低了红麻幼苗根系的DNA甲基化水平。
2.5 甲基化差异片段的测序及qRT-PCR分析
本研究共扩增出291条MSAP多态性片段, 回收到97条DNA甲基化差异片段, 测序比对得到38条具有功能的DNA甲基化差异序列。与植物生长发育和响应逆境相关的甲基化差异序列测序结果见表5, 其中14条DNA甲基化差异序列比对到的同源基因与响应植物抗逆性密切相关, 例如AT-hook家族基因()、SGT家族基因()、S-腺苷-L-蛋氨酸依赖性甲基转移酶家族蛋白(SAM- MET)、NRT1/PTR家族蛋白(NPF5.4)、RaBa家族蛋白(RaBa1f)、Ran GTPase结合蛋白(GTPase)、WD40重复序列蛋白(WD40)、乙烯过表达蛋白(ETO1)、液泡分选蛋白(VPS13F)、β-甘露聚糖酶(β-man6)、类受体蛋白激酶(PLK)、纤维素合成酶(CesA2)、果胶酯酶抑制剂(PME7)、线粒体内膜转位因子(TIM21)。
挑选以上部分DNA甲基化差异基因对600 µmol L-1PbCl2胁迫和对照条件下的红麻幼苗根系进行qRT-PCR分析。由图4可知, 11个与响应植物抗性密切相关的DNA甲基化差异基因中, 有7个基因在PbCl2胁迫下的表达量与对照呈显著性差异。其中基因、、、、的表达量在PbCl2胁迫下显著降低, 基因、、的表达量在PbCl2胁迫下显著升高。说明这些基因的DNA甲基化情况变化与基因表达水平变化密切相关, 并且DNA甲基化变化对基因表达水平的影响是复杂多样的。DNA甲基化水平的升高大多数情况下抑制基因的表达, 但也存在对基因表达水平起促进作用, 或者无显著影响的情况[4]。
泳道1和3代表R I/II酶切, 泳道2和4代表R I/I酶切。1和2: PbCl2-0; 3和4: PbCl2-600。虚线方框: I型(无甲基化); 白色方框: II型(半甲基化); 黑色方框: III型(全甲基化); 虚点方框: IV型(全甲基化)。
The left lane 1, 3 and right lane 2, 4 represent digestion withR I/II andR I/I respectivelyThelane 1, 2, and 3, 4 represent thePbCl2-0, PbCl2-600 respectively. The dotted line frame, white frame, black frame and virtual point frame represent the type I (no methylation), type II (hemi-methylation), type III (full methylation), and type IV (full methylation), respectively.

表4 DNA甲基化水平统计分析
不同小写字母表示在0.05水平差异显著。
Bars superscripted by different lowercase letters indicate significant differences at the 0.05 probability level.
3 讨论
3.1 红麻可以作为修复铅污染土壤的潜在作物
研究表明, 禾本科牧草和黄化枫林木的生物量随着土壤中铅浓度的逐渐增加呈先升高后降低的趋势[24-25], 低浓度铅胁迫可以促进小麦种子的萌发[25]。本研究表明, 低浓度(200 µmol L-1) PbCl2胁迫对红麻幼苗的生长无明显的影响, 而在较高浓度(400 µmol L-1以上)则表现出明显的抑制, 并且一定浓度范围内(400~600 µmol L-1)铅胁迫对幼苗生长的抑制并无显著性差异, 表明红麻具有较强的铅耐受性。由于红麻以收获营养器官为目的, 不进入食物链, 因此生产上可以利用红麻能够吸收土壤中的重金属铅, 并且对其产量无明显影响的优势来改良土壤。
植物在正常的生长发育条件下, 活性氧的产生与清除之间时常处于良性的动态平衡状态[27]。当植物遭受生物或非生物胁迫后, 这种动态平衡就会被打破, 导致植物体内积累大量的活性氧, 例如积累过量的O2–使膜发生过氧化作用, 产生过量丙二醛(MDA), 使细胞膜及细胞器破坏解体, 最终影响植物的生长发育[23-24,27]。与此同时, 当植物遭受逆境胁迫时, 负责清除活性氧的抗氧化酶等系统的活性会相应提高, 使对植物的伤害降到最低, 但当胁迫程度严重时, 会引起抗氧化酶系统紊乱, 无力清除更多的活性氧, 最终严重影响植物的生长发育[28-29]。本研究表明, 不同浓度PbCl2胁迫均显著提高了红麻幼苗根系的O2܋和MDA含量, 以及SOD和CAT的活性, POD活性也在高浓度铅胁迫下显著升高。表明红麻具有较强的抗氧化系统, 可以在一定范围内清除铅胁迫产生的ROS以缓解对红麻生长的影响。
3.2 铅胁迫降低了红麻幼苗根系的DNA甲基化水平
本研究首次检测了红麻幼苗根系的DNA甲基化水平, 并得出600 µmol L-1PbCl2胁迫使幼苗根系的DNA甲基化率和全甲基化率降低, 半甲基化率升高, 即整体上降低了幼苗根系的DNA甲基化水平。这与李雪林等[30]和高桂珍等[31]在其他作物中的研究结果一致, 但也有相反的结论[8-9]。这可能是由于不同作物和不同非生物胁迫的特性, 或者铅胁迫浓度和处理时间不同而导致的。也有研究表明, 枫树在不同浓度盐胁迫下的DNA甲基化水平变化情况不同, 并且相同浓度盐胁迫对不同组织DNA甲基化水平变化的影响也不尽相同[31], 说明植物响应非生物胁迫时, DNA甲基化情况变化的复杂性。
3.3 红麻铅胁迫下DNA甲基化变化参与调控抗性相关基因的表达
本研究发现了7个已报道的与植物抗逆性密切相关的基因在铅胁迫下同时存在DNA甲基化和基因表达水平的差异, 因此我们推测DNA甲基化水平的变化可能参与调控这些基因的表达, 进而调控红麻对铅胁迫的响应。在水稻中已经鉴定出49个与生长发育和响应胁迫相关的果胶酯酶抑制剂PMEI家族基因[21]。PMEI家族基因参与植物油菜素内酯的调节[33], 很多基因也抑制蔗糖转移酶的活性[21], 影响植物的抗逆性。本研究中甲基化差异基因在铅胁迫下几乎不表达, 因此可能对油菜素内酯和蔗糖的合成起到促进作用, 提高红麻幼苗对铅胁迫的适应性。纤维素合酶CesA在纤维素和细胞壁结构的合成中起重要作用[20,34], 拟南芥及基因表达水平的下调可以增强对葡萄孢菌的抗病性[35]。DNA甲基化可能通过调控纤维素合成酶基因的表达促进纤维素的合成, 最终可以提高拟南芥的抗盐性[36]。本研究中纤维素合酶的甲基化情况在铅胁迫下发生了改变, 基因表达量显著下调, 因此推测其甲基化情况的变化在调控基因表达、促进纤维素合成, 进而在响应铅胁迫方面起到了一定的作用。AT-hook家族基因在植物的生长发育、植物激素信号转导和响应逆境胁迫中起重要的作用, 部分AT-hook家族基因表达量的变化很可能参与到了番茄响应逆境胁迫中[19]。NRT1/PTR家族蛋白在植物转运硝酸盐、钾盐、氨基酸和植物激素过程中起重要的作用[37-38]。本研究中AT-hook家族的甲基化差异基因在铅胁迫下几乎不表达, NRT1/PTR家族基因在铅胁迫下的表达量显著降低, 可能会影响营养元素和植物激素的转运, 使红麻幼苗在铅胁迫下生长受抑。有研究报道SGT1家族基因与植物的抗病性密切相关[39], 李为民等[40]研究发现,过表达的转基因烟草对赤星病菌的抗性明显提高, 拟南芥基因突变体对霜霉病的抗性大大降低[39]。本研究中, 甲基化情况的变化使得抗病基因的表达量显著降低, 且该基因也可能参与到了植物非生物胁迫的响应中。
类受体蛋白激酶PLKs广泛参与植物细胞信号转导和逆境胁迫的响应过程, 水稻基因正向调控水稻的抗盐性, 高盐胁迫下其突变体的存活率降低, MDA含量升高, SOD和CAT活性降低[41]。本研究中甲基化差异基因在铅胁迫下的表达量显著升高, 因此可能抑制MDA的积累, 提高抗氧化酶的活性, 进而提高红麻幼苗对铅胁迫的适应性。RaBa家族基因在植物响应生物和非生物胁迫中发挥重要的作用[42], 转基因拟南芥受胁迫的影响更小[43]。核桃WD40转录因子基因参与到了逆境胁迫的响应当中[44], 夏凯文[23]也发现,WD40重复序列结构域蛋白是拟南芥生长发育和胁迫信号传递的关键调控因子。本研究中甲基化差异基因和在铅胁迫下的表达量均显著提高, 因此推测DNA甲基化水平的变化参与以上抗性基因的表达, 从而启动了红麻幼苗对铅胁迫的应急响应。
4 结论
红麻对铅胁迫具有较强的耐受性, 并且一定浓度范围内的铅胁迫对红麻生长的抑制无显著差异, 生产上可以利用红麻来改良重金属铅污染土壤。一定浓度范围内的铅胁迫下, 红麻幼苗可以通过提高自身的抗氧化酶活性来清除过量ROS的产生。600 µmol L-1PbCl2胁迫使幼苗根系整体的DNA甲基化水平降低, 与植物抗逆性密切相关的AT-hook家族基因、NRT1/PTR家族蛋白、RaBa家族蛋白、SGT家族基因、WD40重复序列蛋白、类受体蛋白激酶、纤维素合成酶等相关基因在铅胁迫下都发生了DNA甲基化和基因表达水平的显著变化。推测红麻幼苗可以通过自身的抗氧化酶系统, 及DNA甲基化水平变化来响应铅胁迫, 以消除或减轻铅胁迫对红麻生长的影响。
[1] Wadhwa S K, Tuzen M, Kazi T G, Soylak M, Hazer B. Polyhydroxybutyrate-b-polyethyleneglycol block copolymer for the solid phase extraction of lead and copper in water, baby foods, tea and coffee samples.,2014, 152: 75–80.
[2] Chen P, Ran S, Li R, Huang Z, Qian J, Yu M, Zhou R. Transcriptomeassembly and differentially expressed genes related to cytoplasmic male sterility in kenaf (L).,2014, 34: 1879–1891.
[3] Finnegan E J, Peacock W J, Dennis E S. DNA methylation, a key regulator of plant development and other processes., 2000, 10: 217–223.
[4] Chinnusamy V, Zhu J K. Epigenetic regulation of stress responses in plants., 2009, 12: 133–139.
[5] 葛才林, 杨小勇, 刘向农, 孙锦荷, 罗时石, 王泽港. 重金属对水稻和小麦DNA甲基化水平的影响. 植物生理与分子生物学学报, 2002, 28: 363–368. Ge C L, Yang X Y, Liu X N, Sun J H, Luo S S, Wang Z G. Effects of heavy metal on the DNA methylation level in rice and wheat., 2002, 28: 363–368 (in Chinese with English abstract).
[6] Michael M C, Michael N, Eberhard R. Effect of site-specific modification on restriction endonucleases and DNA modification methyltransferases., 1994, 22: 3640–3559.
[7] 李增强, 史奇奇, 孔祥军, 汤丹峰, 廖小芳, 韦范, 何冰, 莫良玉, 周瑞阳, 陈鹏. 红麻不育系与保持系基因组DNA甲基化比较分析. 中国农业大学学报, 2017, 22(11): 17–27. Li Z Q, Shi Q Q, Kong X J, Tang D F, Liao X F, Wei F, He B, Mo L Y, Zhou R Y, Chen P. Comparative analysis on the kenaf (L.) genomic DNA methylation of its male sterility line and maintainer line., 2017, 22(11): 17–27 (in Chinese with English abstract).
[8] 何玲莉, 沈虹, 王燕, 王娟娟, 龚义勤, 徐良, 柳李旺. 铅胁迫下萝卜基因组DNA甲基化分析. 核农学报, 2015, 29: 1278–1284. He L L, Shen H, Wang Y, Wang J J, Gong Y Q, Xu L, Liu L W. Analysis of genomic DNA methylation level in radish under lead stress., 2015, 29: 1278–1284 (in Chinese with English abstract).
[9] 郭单蒂. 重金属Pb、Cd对中华水韭() DNA甲基化的影响. 哈尔滨师范大学硕士学位论文, 黑龙江哈尔滨, 2014. Guo S D. The Effect of Heavy metals of Pb and Cd on DNA Methylation in. MS Thesis of Harbin Normal University, Harbin, Heilongjiang, China, 2014 (in Chinese with English abstract).
[10] 殷欣. 镉胁迫下大豆生理生化特性及DNA甲基化变异的研究. 哈尔滨师范大学硕士学位论文, 黑龙江哈尔滨, 2016. Yin X. Soybean Physiological and Biochemical Characteristics and DNA Methylation Variation under Cadmium Stress. MS Thesis of Harbin Normal University, Harbin, Heilongjiang, China, 2016 (in Chinese with English abstract).
[11] Sciandrello G, Caradonna F, Mauro M, Barbata G. Arsenic- induced DNA hypomethylation affects chromosoma instability in mammalian cells., 2004, 25: 413–417.
[12] 曹翠玲, 麻鹏达. 植物生理学教学实验指导. 西安: 西北农林科技大学出版社, 2016. pp 96–99. Cao C L, Ma P D. Experimental Guidance of Plant Physiology. Xi’an: Northwest A&F University Press, 2016. pp 96–99 (in Chinese).
[13] 王学奎, 黄见良. 植物生理生化实验原理与技术. 北京: 高等教育出版社, 2015. pp 282–286. Wang X K, Huang J L. The Principle and Technology of Plant Physiology and Biochemistry Experiment. Beijing: Higher Education Press, 2015. pp 282–286 (in Chinese).
[14] Tang X M, Tao X, Wang Y, Ma D W, Li D, Yang H, Ma X R. Analysis of DNA methylation of perennial ryegrass under drought using the methylation-sensitive amplification polymorphism (MSAP) technique., 2014, 289: 1075–1084.
[15] 李荣华, 夏岩石, 刘顺枝, 孙莉丽, 郭培国, 缪绅裕, 陈健辉. 改进的CTAB提取植物DNA方法. 实验室研究与探索, 2009, 28(9): 14–16. Li R H, Xia Y S, Liu S Z, Sun L L, Guo P G, Miao S Y, Chen J H. CTAB-improved method of DNA extraction in plant.,2009, 28(9): 14–16 (in Chinese with English abstract).
[16] Zhang L W, Xu Y, Zhang X T, Ma X K, Zhang L L, Liao Z Y, Zhang Q, Wan X B, Chang Y, Zhang J S, Li D X, Zhang L M, Xu J T, Tao A F, Lin L H, Fang P P, Chen S, Qi R, Xu X M, Qi J M, Ming R. The genome of kenaf (L.) provides insights into bast fibre and leaf shape biogenesis., 2020, 18: 1796–1809.
[17] 王春晖, 赵云雷, 王红梅, 陈伟, 龚海燕, 桑晓慧. 适用于转录组测序的棉花幼根总RNA提取方法筛选. 棉花学报, 2013, 25: 372–376. Wang C H, Zhao Y L, Wang H M, Chen W, Gong H Y, Sang X H. Screening of methods to isolate high-quality total RNA from young cotton roots., 2013, 25: 372–376 (in Chinese with English abstract).
[18] 原晓龙, 陈剑, 陈中华, 华梅, 王娟, 王毅. 滇牡丹类黄酮7-O-葡萄糖基转移酶基因的鉴定与表达分析. 西部林业科学, 2018, 47(5): 19–25. Yuan X L, Chen J, Chen Z H, Hua M, Wang J, Wang Y. Characterization and expression analysis of a favonoid 7-O-glucosyltransferase gene in.2018, 47(5): 19–25 (in Chinese with English abstract).
[19] 丁丽雪, 李涛, 李植良, 徐小万, 李颖, 王恒明, 王永飞, 马三梅, 黎振兴. 番茄AT-hook基因家族的鉴定及胁迫条件下的表达分析. 植物遗传资源学报, 2016, 17: 303–315. Ding L X, Li T, Li Z L, Xu X W, Li Y, Wang H M, Wang Y F, Ma S M, Li Z X. Genome-wide identification and expression analysis in oxidative stress of AT-hook gene family in tomato., 2016, 17: 303–315 (in Chinese with English abstract).
[20] Ana Caño-Delgado, Penfield S, Smith C, Catley M, Bevan M. Reduced cellulose synthesis invokes lignification and defense responses in., 2003, 34: 351–362.
[21] Nguyen H P, Jeong H Y, Kim H, Kim Y C, Lee C. Molecular and biochemical characterization of rice pectin methylesterase inhibitors (OsPMEIs)., 2016, 101: 105–112.
[22] 陈江淑, 邓治, 刘辉, 范玉龙, 姜达, 夏立琼, 夏志辉, 李德军. 巴西橡胶树基因克隆、进化及表达分析. 植物生理学报, 2015, 51: 1735–1742. Chen J S, Deng Z, Liu H, Fan Y L, Jiang D, Xia L Q, Xia Z H, Li D J. Cloning, phylogenetic and expression analyses of-gene in., 2015, 51: 1735–1742 (in Chinese with English abstract).
[23] 夏凯文. 拟南芥WD40家族蛋白TAWD在抵御高温胁迫中的功能研究. 华中师范大学硕士学位论文, 湖北武汉, 2019. Xia K W. Function Study of the WD40-repeat Protein TAWA inThermotolerance. MS Thesis of Central China Normal Unversity, Wuhan, Hubei, China, 2019 (in Chinese with English abstract).
[24] 李慧芳, 王瑜, 袁庆华, 赵桂琴. 铅胁迫对禾本科牧草的生长及体内酶活性的影响. 种子, 2014, 33(8): 1–7 Li H F, Wang Y, Yuan Q H, Zhao G Q. The impacts of lead stress on the growth of forage grasses and their enzyme activities., 2014, 33(8): 1–7 (in Chinese with English abstract).
[25] 张博宇. 铅胁迫对黄花风铃木幼苗生长和生理的影响. 广西大学硕士学位论文, 广西南宁, 2019. Zhang B Y. Effects of Lead Stress on Growth and Physiology ofSeedlings. MS Thesis of Guangxi University, Nanning, Guangxi, China, 2019 (in Chinese with English abstract).
[26] 周秋峰, 于沐, 赵建国, 张果果. 重金属胁迫对小麦生长发育及相关生理指标的影响. 中国农学通报, 2017, 33(33): 7–14. Zhou Q F, Yu B, Zhao J G, Zhang G G. Heavy metal stress on growth and development and physiological indexes of wheat., 2017, 33(33): 7–14 (in Chinese with English abstract).
[27] Roychoudhury A, Basu S, Sarkar S N, Sengupta D N. Comparative physiological and molecular responses of a common aromaticrice cultivar to high salinity with non-aromaticrice cultivars., 2008, 27: 1395–1410.
[28] Liang W, Ma X, Wan P, Liu L. Plant salt-tolerance mechanism: a review., 2017, 495: 286–291.
[29] Zhu J K. Abiotic stress signaling and responses in plants., 2016, 167: 313–324.
[30] 李雪林, 林忠旭, 聂以春, 郭小平, 张献龙. 盐胁迫下棉花基因组DNA表观遗传变化的MSAP分析. 作物学报, 2009, 35: 588–596. Li X L, Lin Z X, Nie Y C, Guo X P, Zhang X L. MSAP analysis of epigenetic changes in cotton (L.) under salt stress., 2009, 35: 588–596 (in Chinese with English abstract).
[31] 高桂珍, 应菲, 陈碧云, 李浩, 吕晓丹, 闫贵欣, 许鲲, 伍晓明. 热胁迫过程中白菜型油菜种子DNA的甲基化. 作物学报, 2011, 37: 1597–1604. Gao G Z, Ying F, Chen B Y, Li H, Lyu X D, Yan G X, Xu K, Wu X M. DNA methylation of seed in response to heat stress inL, 2011, 37: 1597–1604 (in Chinese with English abstract).
[32] Mastan S G, Rathore M S, Bhatt V, Yadav P, Chikara J. Assessment of changes in DNA methylation by methylation-sensitive amplification polymorphism inL. subjected to salinity stress., 2012, 508: 125–129.
[33] Wolf S, Mravec J, Greiner S, Grégory M, Hofte H. Plant cell wall homeostasis is mediated by brassinosteroid feedback signaling., 2012, 22: 1732–1737.
[34] Ellis C. Themutanthas constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens., 2001, 13: 1025–1033.
[35] Vicente R, Javier G A, Pablo V. Enhanced disease resistance to Botrytis cinereal inplants is associated to an early downregulation ofgenes., 2011, 6: 911–913.
[36] 王津, 韩榕. DNA甲基转移酶赋予拟南芥盐胁迫耐受性. 江苏农业学报, 2019, 35: 1028–1031. Wang J, Han R. DNA methyltransferases confer salt stress tolerance in., 2019, 35: 1028–1031 (in Chinese with English abstract).
[37] Li H, Yu M, Du X Q, Wang Z F, Wu W H, Quintero F J, Jin X H, Li H D, Wang Y. NRT1.5/NPF7.3 functions as a proton-coupled H+/K+antiporter for K+loading into the xylem in., 2017, 29: 2016–2026.
[38] Criado M V, Roberts I N, Echeverria M, Barneix A J. Plant growth regulators and induction of leaf senescence in nitrogen-deprived wheat plants., 2007, 26: 301–307.
[39] Peart J R, Lu R, Sadanandom A, Malcuit I, Moffett P, Brice D C, Baulcombe D C. Ubiquitin ligase-associated protein SGT1 is required for host and nonhost disease resistance in plants., 2002, 99: 10865–10869.
[40] 李为民. 海岛棉、、、基因及中棉基因启动子的克隆和功能分析. 中国农业科学院博士学位论文, 北京, 2003. Li W M. Cloning and Functional Analysis of,,,Genes andPromoter. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2003 (in Chinese with English abstract).
[41] 路凯, 张亚东, 朱镇, 陈涛, 赵庆勇, 赵凌, 周丽慧, 姚姝, 赵春芳, 梁文化, 魏晓东, 王才林. 水稻类受体蛋白激酶RLKN1在调节植物耐盐性中的功能研究. 见: 江苏省遗传学会2019年学术研讨会论文集. 南京: 江苏省遗传学会, 2019. p 10. Lu K, Zhang Y D, Zhu Z, Chen D, Zhao Q Y, Zhao L, Zhou L H, Yao S, Zhao C F, Liang W H, Wei X D, Wang C L. Function of rice receptor-like protein kinase PLKN1 in regulating plant salt tolerance. In: Proceedings of the 2019 Academic Seminar of Jiangsu Genetics Society. Nanjing: Jiangsu Genetics Society, 2019. p 10 (in Chinese).
[42] Asaoka R, Uemura T, Ito J, Fujimoto M, Ito E, Ueda T, Nakano A.RABA1 GTPases are involved in transport between the trans-Golgi network and the plasma membrane, and are required for salinity stress tolerance., 2013, 73: 240–249
[43] 代丽红. 杨树盐逆境响应基因的功能研究. 黑龙江大学硕士学位论文, 黑龙江哈尔滨, 2014. Dai L H. Study on the Function of Salt Stress Response Genein Poplar. MS Thesis of Heilongjiang University, Harbin, Heilongjiang, China, 2014 (in Chinese with English abstract).
[44] 陈淑雯, 郝茜珣, 贾彩霞, 赵爱国, 李大培, 杨桂燕. 核桃WD40转录因子基因的克隆及逆境响应. 植物遗传资源学报, 2018, 19: 979–986. Chen S W, Hao Q X, Jia C X, Zhao A G, Li D P, Yang G Y. Identification and stress response analysis of a WD40 transcription actorgene from., 2018, 19: 979–986 (in Chinese with English abstract).
Physiological characteristics and DNA methylation analysis under lead stress in kenaf (L.)
LI Zeng-Qiang1, DING Xin-Chao1, LU Hai1, HU Ya-Li1, YUE Jiao1, HUANG Zhen1, MO Liang-Yu1, CHEN Li1, CHEN Tao2, and CHEN Peng1,*
1College of Agriculture, Guangxi University / Guangxi Colleges and Universities Key Laboratory of Plant Genetics and Breeding, Nanning 530004, Guangxi, China;2Guangxi Subtropical Crops Research Institute, Nanning 530004, Guangxi, China
DNA methylation plays an important role in response to plant biotic and abiotic stresses, but there are few reports on the changes of plant DNA methylation level under lead stress. In this study, kenaf (L.) P3A was used as the material, the seedlings were cultured in Hoagland solution, and treated with at different concentrations (0, 200, 400, and 600 µmol L-1) of PbCl2. The changes of agronomic traits, ROS content and antioxidant enzyme activity of root were investigated. The changes of DNA methylation level in roots under 600 µmol L–1lead stress were determined by methylation-sensitive amplification polymorphism (MSAP). The differentially methylated genes (DMGs) were cloned, sequenced, and functionally annotated. In addition, the expression levels of DMGs were investigated by qRT-PCR. The results showed that the stem diameter, root length and root surface area of seedlings were significantly inhibited by different concentrations of PbCl2stress. And the plant height and total fresh weight of kenaf seedlings were significantly reduced under 400 µmol L-1concentration or more of lead stress. The content of lead, O2܋and MDA, and activities of SOD were increased significantly in kenaf seedlings roots, CAT activity increased first and then decreased with the increase of lead concentration, the POD activity showed a trend of decreasing first and then increasing. MSAP analysis of roots treated with 0 µmol L-1and 600 µmol L-1PbCl2showed that DNA methylation rates were 71.13%, 62.20%, fully methylated ratio was 50.52%, 37.80%, and hemi-methylated ratio were 20.62%, 24.40%, respectively. In other words, lead stress significantly reduced DNA methylation rate and total methylation rate, whereas increased the hemi-methylation rate of roots of kenaf seedlings. qRT-PCR analysis showed that there were also differences in gene expression of seven DMGs closely related to resistance. It suggested that the change of DNA methylation level played an important role in kenaf response to lead stress in kenaf. This study provides a theoretical basis for further exploring the potential mechanism of DNA methylation in response to plant abiotic stress, and improving soil lead pollution in kenaf production.
kenaf; lead stress; DNA methylation; MSAP; qRT-PCR; antioxidant enzyme system
10.3724/SP.J.1006.2021.04104
本研究由国家自然科学基金项目(31560341, 31960368)和国家现代农业产业技术体系建设专项(CARS-16-E14)资助。
This work was supported by the National Natural Science Foundation of China (31560341, 31960368) and the China Agriculture Research System (CARS-16-E14).
陈鹏, E-mail: hustwell@gmail.com
E-mail: 1134485681@qq.com
2020-05-08;
2020-08-19;
2020-09-10.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20200908.1800.005.html
猜你喜欢
现代园艺(2022年7期)2022-11-19
林业科技(2022年5期)2022-10-08
上海师范大学学报·自然科学版(2022年3期)2022-07-11
昆明医科大学学报(2022年1期)2022-02-28
今日农业(2021年10期)2021-11-27
今日农业(2021年9期)2021-11-26
诗潮(2021年1期)2021-03-15
魅力中国(2020年23期)2020-12-08
湖北农业科学(2019年22期)2019-12-23
中国火炬(2015年12期)2015-07-31
