
步云山森林植物群落物种多样性及其与海拔因子的关系
2021-04-02 13:10:02张恒庆王海双刘静怡李佳欣
辽宁师范大学学报(自然科学版) 2021年1期
张恒庆, 王海双, 焦 悦, 刘静怡, 李佳欣
(辽宁师范大学 生命科学学院,辽宁 大连 116081)
植物群落中蕴藏着丰富的野生植物种质资源,为人类和动物提供食物和栖息地,具有不可替代的作用[1].植物群落多样性是研究群落非常重要的内容,也是反映群落功能、群落结构以及物种类型的直接指标[2].植物群落多样性与植物所处区域内的环境气候因子、物种的进化、迁移、演变等因素密切相关[3],海拔梯度作为环境因子被认为是影响物种多样性格局的重要因素.有关步云山植物群落的相关研究还未见报道.本文从植物群落类型的划分、植物群落物种多样性及物种多样性与海拔因子的关系等方面对步云山森林植物群落进行研究,以期为步云山生物多样性保护和自然资源的可持续利用提供科学的依据.
1 研究区域概况
步云山南坡位于大连庄河境内,隶属步云山乡管辖,北坡位于营口盖州市境内,隶属盖州市矿洞沟镇,是营口与大连两市境内最高峰,号称“辽南第一峰”.步云山属长白山系,千山余脉,海拔1 130.7 m,山体陡峻,山势雄伟挺拔,平均坡度25°~35°.步云山地处北温带,属暖温带湿润大陆性季风气候,具有一定的海洋性气候特征,气候温和,四季分明.历年平均气温为9.3 ℃,极端最高气温36.0 ℃,极端最低气温-28.1 ℃,历年平均降水量为736.0 mm[4].优越的水热条件配比再加上较小的人为干扰造就了步云山丰富的植物资源,群落类型多样.
2 研究方法
2.1 植物调查
参照原国家环境保护部下发的《生物多样性观测技术导则陆生维管植物(HJ 710.1—2014)》,结合步云山的地形地貌特征及植物分布规律,确定科学的调查方案.本研究采用样方、样线相结合的方法于2019年、2020年多次对步云山不同海拔高度的植物进行调查.森林样方的取样单元为30 m×30 m,对30 m×30 m乔木样方内的乔木树种进行调查记录,在乔木样方四角分别设置4个10 m×10 m灌木样方并调查记录灌木树种,于乔木样方四角及中心位置设置5个2 m×2 m草本样方并调查记录草本植物.本研究共设置了18个森林群落样方和6条踏查样线,样线总长度共计4 864 m.步云山样方、样线设置见表1、表2.

表1 森林群落样方设置表
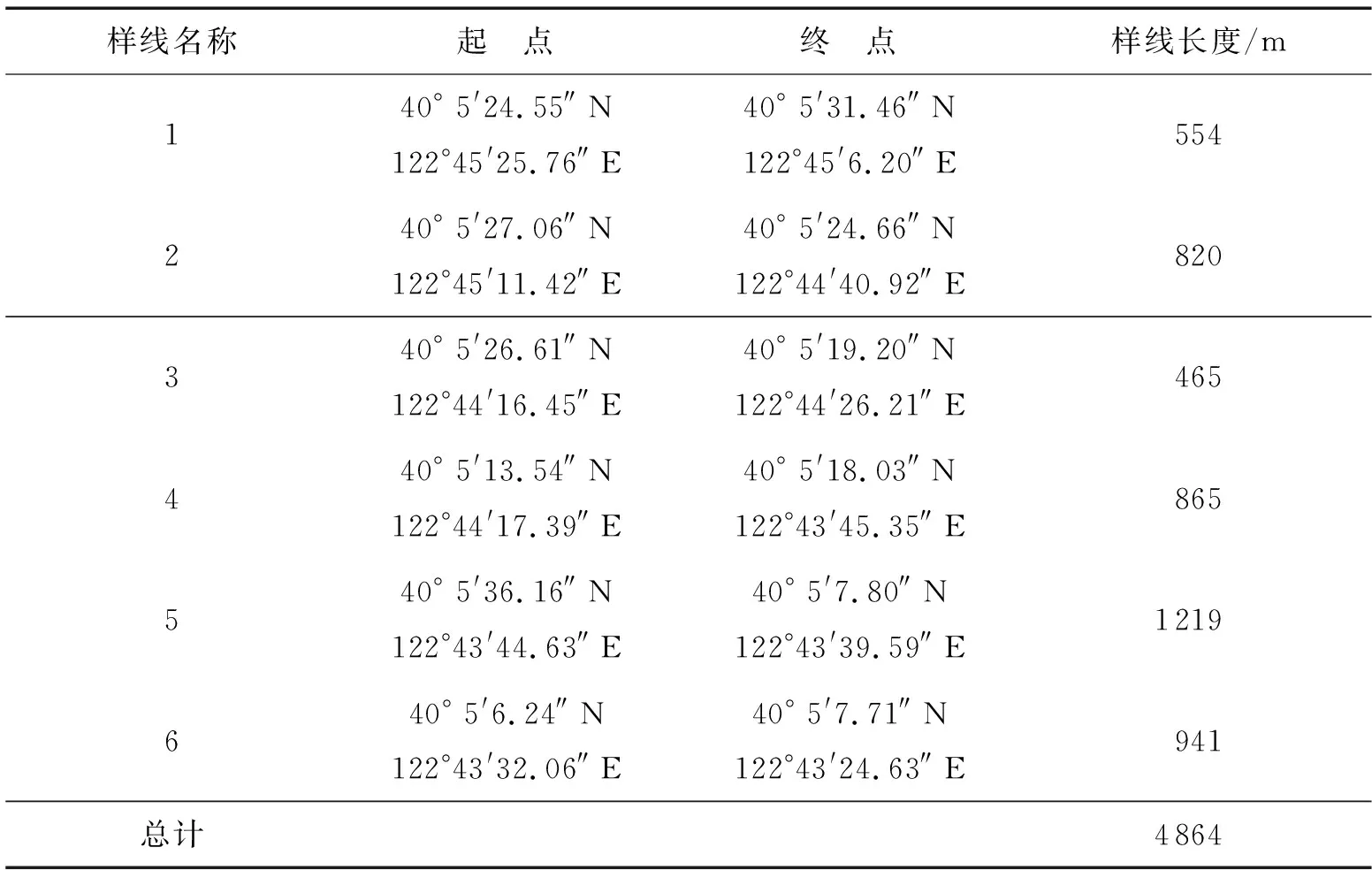
表2 样线设置表
2.2 数据处理
2.2.1 植物鉴定
在调查过程中记录物种相关的形态特征等方面信息,对野外未能准确识别的物种采集标本.植物种类鉴定主要依据《中国植物志》[5]《大连地区植物志》[6]《大连植物彩色图谱》[7],对仍有疑问的物种请两位以上专家鉴定.
2.2.2 植物群落分类
依据《中国植被》[8]《辽宁植被与植被区划》[9]等书籍中的分类系统及张恒庆等人[10]对植物群落分类的研究对步云山植被进行分类和命名.
2.2.3 物种多样性计算
本文采用物种丰富度指数、Shannon-Wiener指数、Simpson指数及Pielou均匀度指数[11]进行计算,计算公式如下:
物种丰富度指数(S):
S=N.
(1)
Shannon-Wiener指数(H′):
(2)
Simpson指数(D):
(3)
Pielou指数(E):
E=H′/lnS.
(4)
其中,Pi为第i个种的个体数占群落(样地)中所有种的总个体数的比例,S为物种数,N为群落(样地)中所有物种个体总数.
注:野外调查中草本植物很难完整地统计每种的个体数量,故本研究草本植物的Pi值用每种的种群盖度代替.
2.3 数据分析
数据采用Excel和Past软件,对不同数据进行整理计算、趋势分析、图形绘制等.
3 结果与分析
调查结果显示步云山共有植物79科226属379种,被子植物66科210属352种.其中,双子叶植物60科180属308种,单子叶植物6科30属44种.裸子植物1科1属1种,蕨类植物12科15属26种.在此基础上,对森林群落的划分、森林植物群落物种多样性及物种多样性与海拔因子的关系进行了分析.
3.1 森林植物群落类型的划分
依据群落分类原则,将步云山植物群落划分为13个典型森林群落类型,具体如下:
(1)日本落叶松林(Larixkaempferi)
为人工林,分布在海拔440~460 m范围内,坡度5°~10°,坡向北,乔木层郁闭度52%,以日本落叶松为优势种,高度在14 m左右,胸径18 cm左右,伴生少量紫花槭(Acerpseudosieboldianum)和色木槭(Acermono);灌木层植被盖度为7%,主要优势种为牛叠肚(Rubuscrataegifolius),伴生有早锦带花(Weigelapraecox)、阴山胡枝子(Lespedezainschanica)等;草本层盖度为30%,主要优势种为球果堇菜(Violacollina),混生有京黄芩(Scutellariapekinensis)等.
(2)花曲柳+色木槭林(Fraxinuschinensis+Acermono)
主要分布于海拔460~490 m范围内,部分地区也有少量分布,坡度30°~35°,坡向东北,乔木层郁闭度60%左右,花曲柳高度8~10 m,胸径8~11 cm,色木槭高度6~10 m,胸径4~9 cm,伴生有天女花(Oyamasieboldii)、千金榆(Carpinuscordata)等;灌木层植被盖度23%,主要优势种为毛榛(Corylusmandshurica),伴生有八角枫(Alangiumchinense)等;草本层植被盖度63%,主要优势种为兴安升麻(Cimicifugadahurica),混生有粗茎鳞毛蕨(Dryopteriscrassirhizoma)、蒙古堇菜(Violamongolica)等.
(3)花曲柳+蒙古栎林(Fraxinuschinensis+Quercusmongolica)
分布在海拔500~520 m范围内,坡度25°~40°,坡向北,乔木层郁闭度78%左右,花曲柳高度8~10 m,胸径6 cm左右,蒙古栎高度8 m左右,胸径7 cm左右.此群落类型中有多种伴生种,如山樱桃(Prunusserrulata)、紫椴(Tiliaamurensis)等;灌木层植被盖度为18%,主要优势种为大字杜鹃(Rhododendronschlippenbachii),伴生有光萼溲疏(Deutziaglabrata)等;草本层盖度为49%,宽叶薹草(Carexsiderosticta)为主要优势种,混生有黄花堇菜(Violaxanthopetala)、禾秆蹄盖蕨(Athyriumyokoscense)等.
(4)紫花槭+蒙古栎林(Acerpseudosieboldianum+Quercusmongolica)
分布在海拔530~570 m范围内,坡度20°~30°,坡向北,乔木层郁闭度62%,紫花槭高度在7 m左右,胸径5~6 cm,蒙古栎高度在8.5 m左右,胸径10~12 cm,伴生有黄檗(Phellodendronamurense)等;灌木层植被盖度为3%,主要植物为卫矛(Euonymusalatus);草本层盖度为15%,主要优势种为宽叶薹草,混生有矮丛薹草(Carexcallitrichos)、异叶车叶草(Asperulamaximowiczii)等.
(5)胡桃楸林(Juglansmandshurica)
胡桃楸林分布在海拔570~600 m范围内,坡度15°~25°,坡向南,乔木层郁闭度40%左右,主要优势种为胡桃楸,高度在9 m左右,胸径10~12 cm,伴生有大果榆(Ulmusmacrocarpa)、山楂(Crataeguspinnatifida)等;灌木层植被盖度为14%,主要优势种为牛叠肚,伴生有金银忍冬(Loniceramaackii)、锦带花(Weigelaflorida)等;草本层盖度为80%,甘野菊(Chrysanthemumboreale)为主要优势种,种群盖度50%,混生有益母草(Leonurusjaponicus)、茵陈蒿(Artemisiacapillaris)等.
(6)花曲柳+灯台树林(Fraxinuschinensis+Cornuscontroversa)
分布在海拔600~620 m范围内,坡度40°~45°,坡向东北,乔木层郁闭度56%,花曲柳和灯台树两者高度均在10 m左右,胸径10~15 cm,但花曲柳数量较多,伴生有裂叶榆(Ulmuslaciniata)等;灌木层植被盖度为27%,主要优势种为八角枫;草本层植被盖度为9%,主要物种有二苞黄精(Polygonatuminvolucratum)等.
(7)蒙古栎+紫花槭林(Quercusmongolica+Acerpseudosieboldianum)
分布在海拔620~650 m范围内,坡度25°~35°,坡向西北,乔木层郁闭度75%,蒙古栎高度在10~12 m,胸径15~20 cm,紫花槭高度6 m左右,胸径8~11 cm,伴生有朝鲜槐(Maackiaamurensis)等;灌木层植被盖度为1%,主要植物为卫矛;草本层植被盖度为13%,主要优势种为龙常草(Diarrhenamandshurica),混生有凤凰堇菜(Violafunghuangensis)等.
(8)天女花+千金榆林(Oyamasieboldii+Carpinuscordata)
分布在海拔650~680 m范围内,坡度10°~20°,坡向西南,乔木层郁闭度75%,天女花高度6~8 m,胸径9~11 cm,千金榆高度6~7 m,胸径6~9 cm左右,伴生有花曲柳、黄檗等;灌木层植被盖度为3%,主要优势种为卫矛,伴生有毛榛、光萼溲疏等;草本层盖度为11%,主要物种有毛筒玉竹(Polygonatuminflatum)、辽宁堇菜(Violarossii)等.
(9)紫花槭+日本落叶松+蒙古栎林(Acerpseudosieboldianum+Larixkaempferi+Quercusmongolica)
分布在海拔680~710 m范围内,坡度10°~20°,坡向东北,乔木层郁闭度70%,紫花槭高度6.5 m左右,胸径4~6 cm,日本落叶松高度11 m左右,胸径15~20 cm,蒙古栎高度约9 m,胸径约10 cm;灌木层植被盖度为7%,主要优势种为卫矛,伴生短翅卫矛(Euonymusrehderianus)、东北接骨木(Sambucusmanshurica)等;草本层盖度为45%,以蔓孩儿参(Pseudostellariadavidii)为优势种,混生有掌叶铁线蕨(Adiantumpedatum)等.
(10)紫花槭+千金榆林(Acerpseudosieboldianum+Carpinuscordata)
分布在海拔720~750 m范围内,坡度25°~30°,坡向北,乔木层郁闭度65%,紫花槭高度约7 m,胸径6~8 cm,千金榆高度约7 m,胸径约6 cm,伴生有花楸树(Sorbuspohuashanensis)等;灌木层植被盖度为5%,主要优势种为毛榛;草本层植被盖度为8%,主要优势种为苦荬菜(Ixerispolycephala)等.
(11)蒙古栎+色木槭林(Quercusmongolica+Acermono)
分布在海拔850~890 m范围内,坡度30°~35°,坡向东,乔木层郁闭度67%,蒙古栎高度8~9 m,胸径10 cm左右,色木槭高度6~8 m,胸径4~6 cm,伴生水榆花楸(Sorbusalnifolia)等;灌木层植被盖度为13%,主要优势种为卫矛,伴生短翅卫矛等;草本层植被盖度为47%,主要植物为荨麻叶龙头草(Meehaniaurticifolia)、牧根草(Asyneumajaponicum)等.
(12)蒙古栎林(Quercusmongolica)
分布在海拔900~940 m范围内,坡度25°~35°,坡向东南,乔木层郁闭度85%,主要优势种为蒙古栎,高度在7 m左右,胸径10~12 cm;灌木层植被盖度为26%,主要优势种为大字杜鹃,伴生锦带花等;草本层盖度为33%,主要植物有菴闾(Artemisiakeiskeana)、大叶柴胡(Bupleurumlongira-diatum)等.
(13)花曲柳林(Fraxinuschinensis)
花曲柳纯林分布在海拔1 000~1 060 m左右,坡度30°~40°,坡向东,乔木层郁闭度57%,主要优势种为花曲柳,高度在6 m左右,胸径5~7 cm,伴生有少量色木槭和朝鲜槐;灌木层植被盖度为7%,主要灌木植物有大叶小檗(Berberisferdinande-coburgii)等;草本层植被盖度为49%,蹄叶橐吾(Ligulariafischeri)为主要优势种,混生有返顾马先蒿(Pedicularisresupinata)等.
3.2 森林植物群落的物种多样性
分别计算步云山13个森林群落乔、灌、草3层的物种多样性指数,结果见表3.
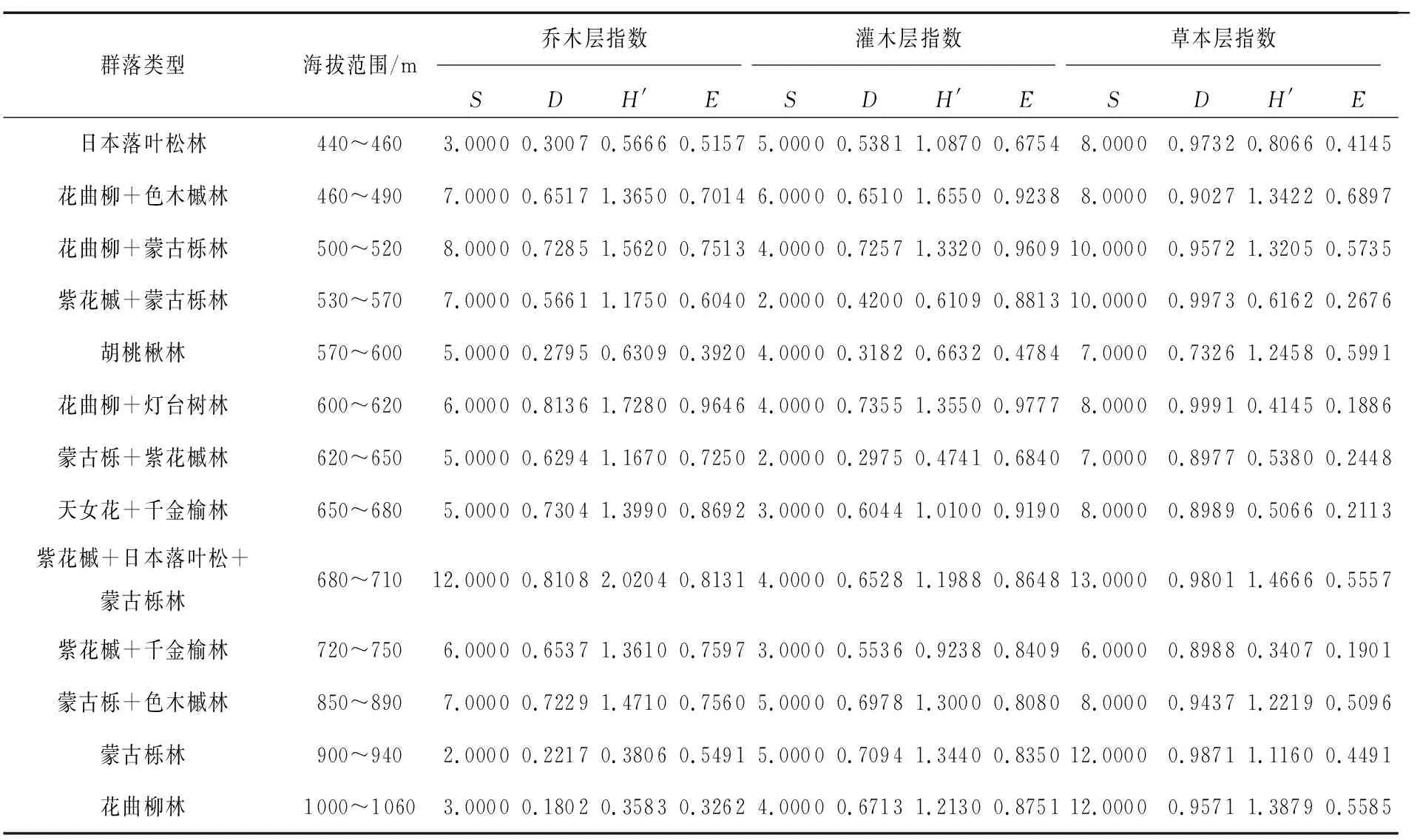
表3 各群落类型乔木层、灌木层和草本层的物种多样性指数
从表3可以看出,步云山各植物群落不同层次的丰富度指数和Simpson指数整体呈草本层>乔木层>灌木层.草本层植物丰富度指数和Simpson指数最大的原因在于草本植物对极端环境的适应性很强,无论在低海拔还是高海拔区域均能长出适应环境的草本植物,而乔木层丰富度指数和Simpson指数大于灌木层指数与以往卢鹏林[12]、尹达[13]、何亚珍[14]等人对不同山体的研究结果灌木层>乔木层的物种多样性指数并不一致.但根据对步云山的实地调查发现,该山体人为干扰较小,乔木层植物群落占绝对优势,多为落叶阔叶混交林,主要建群种的个体数量很多并有大量伴生乔木,再加上本研究将乔木幼树也归为了乔木,所以乔木层物种多样性较高.乔木层植物长势良好又会使整个群落林下光照条件变差,所以灌木种类和数量都很少,物种多样性较低,故步云山的研究结果符合当地实际情况.日本落叶松林、胡桃楸林、蒙古栎林、花曲柳林这4种群落类型中丰富度指数和Simpson指数呈草本层>灌木层>乔木层,是因为4种纯林位于较高或较低海拔区域,而高海拔和低海拔区域水热配比条件并不适合所有乔木层植物的生长,造成乔木层植物相对低矮,种类单一,物种多样性低.乔木层植物生长不良对灌木层植物来说非常有利,它们可以获得更多的光照等生长要素,故物种多样性较高.均匀度指数是指群落中不同物种的多度分布的均匀程度[15],步云山各植物群落不同层次的均匀度指数整体呈灌木层>乔木层>草本层.灌木层均匀度指数偏大是因为步云山灌木层物种丰富度很低,每一物种的个体数也很少,物种间个体数目差异不明显,所以均匀度指数偏大;乔木层的物种相对于灌木层较丰富,每一物种的个体数有一定差异,所以均匀度次于灌木层;草本植物多成簇分布,导致样地内物种之间个体数差异较大,分布不均匀,所以均匀度指数最小.Shannon-Wiener指数在各群落的乔、灌、草3层差异不明显,看不出明显的变化趋势.
3.3 森林植物群落物种多样性与海拔因子的关系分析
海拔梯度综合了温度、湿度、光照等多种环境因子,是物种多样性梯度格局研究的重要方面.物种多样性沿海拔梯度的分布格局一般有5种形式[16]:分别是沿海拔梯度先降后升、单峰曲线、单调升高、单调下降和无明显格局[17].本研究以上文3.2计算的13个森林群落乔、灌、草3层的物种多样性指数为纵坐标,海拔梯度为横坐标进行趋势曲线拟合,图1~图3显示了不同群落的乔、灌、草3层物种多样性指数在海拔梯度上的变化趋势及多项式拟合效果.
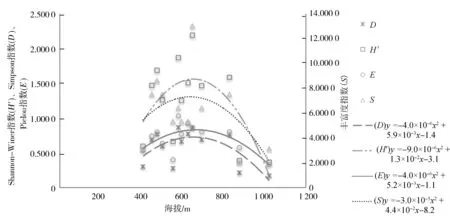
图1 不同群落类型的乔木层物种多样性指数沿海拔高度的变化趋势Fig.1 Variation trend of tree layer species diversity index of different community types along elevation

图2 不同群落类型的灌木层物种多样性指数沿海拔高度的变化趋势Fig.2 Variation trend of shrub layer species diversity index of different community types along elevation
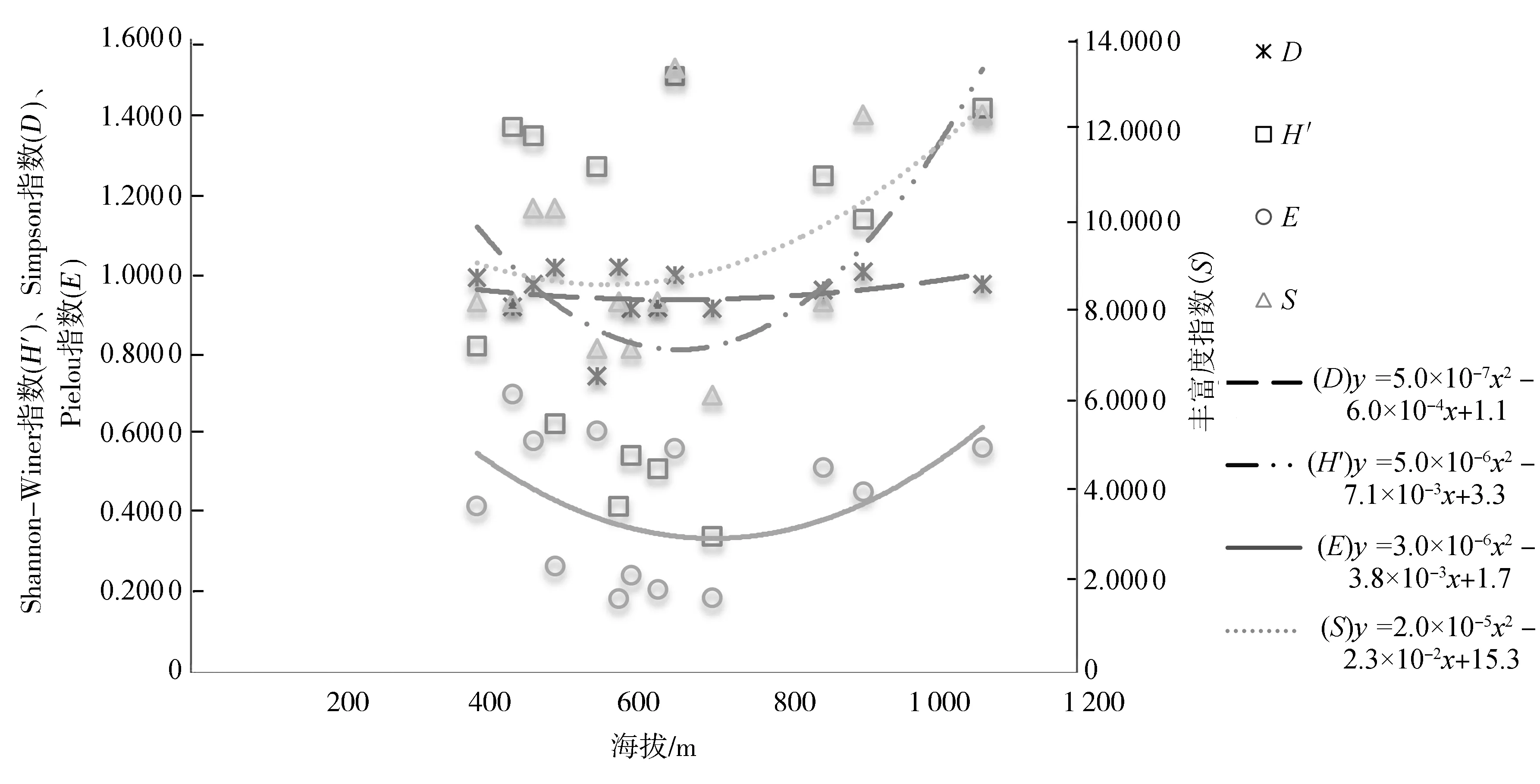
图3 不同群落类型的草本层物种多样性指数沿海拔高度的变化趋势Fig.3 Variation trend of herb layer species diversity index of different community types along elevation
从图1~图3可以看出,步云山乔木层物种多样性指数均随海拔的升高呈先上升后下降的单峰模式,最高值出现在海拔600~800 m的中海拔地段.在大多数情况下,低海拔区域温度较高而水分相对缺少,加之受人为干扰的影响较大而产生负面影响[18],使低海拔地区乔木层植物生长受限;中海拔地区的温度和降水量适宜,土壤深厚发育好,保水性较好,其水热条件更适宜植被生长,加之人为干扰相对较少[19],使中海拔地区乔木层植物生长良好,不但建群种数量庞大,还拥有较多的伴生乔木,郁闭度高,物种多样性指数大;高海拔地区主要由于热量限制,太阳辐射强、昼夜温差大等环境条件严重影响乔木层植物生长发育,因此物种多样性较低.灌木层和草本层的物种多样性呈现先下降后上升的趋势,最低值出现在海拔600~800 m.林下层的灌木和草本植物更容易受到微环境,例如人为干扰和森林郁闭度的影响[20],森林郁闭度会影响林下层的光照强度、湿度和温度[21].低海拔地区降水缺乏限制了乔木植物的生长,从而为灌木和草本植物提供了较多的生存空间和较有利的生态环境;中海拔地区乔木长势良好,郁闭度较大,遮挡住了灌木和草本植物生长所需要的光照,不利于灌木和草本植物的生长,所以灌木和草本植物物种多样性较低;高海拔地区虽然温度低,但乔木层植物矮小,这使得整个群落林下的光照强度增加,灌木层和草本层植物能更好地吸收阳光,物种多样性变高.
4 结 论
步云山共有植物79科226属379种,划分为13个森林群落类型,森林群落多样,物种多样性较高.各植物群落的丰富度指数和Simpson指数整体呈现草本层>乔木层>灌木层的趋势;Shannon-Wiener指数在各群落的乔、灌、草3层中波动较小;而均匀度指数则整体呈灌木层>乔木层>草本层.不同群落的乔木层植物物种多样性沿海拔梯度整体呈先上升后下降的趋势,灌木层和草本层的物种多样性呈现先下降后上升的趋势.
整体来看,步云山森林植物群落类型多样,物种多样性高,乔木层植物物种多样性受海拔梯度的影响变化较大,灌木层和草本层植物受乔木层植物的高度、郁闭度等的影响,所以也间接受海拔梯度的影响.通过对步云山森林植物群落物种多样性及其与海拔因子关系的研究,揭示了步云山各森林群落类型的物种组成、海拔分布范围及沿海拔梯度的物种多样性变化,为步云山森林生态系统生物多样性和生态环境的保护提供了科学的理论依据.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
少儿美术(2020年8期)2020-11-06 05:21:38
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
福建林业(2020年5期)2020-03-18 08:23:02
水土保持研究(2019年6期)2019-10-19 03:33:32
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
