
中国泥鳅属和副泥鳅属鱼类的分类整理
2021-04-01 07:56:12王银肖杨慧兰谭慧敏陈咏霞
水生生物学报 2021年2期
张 慧 王银肖 杨慧兰 谭慧敏 陈咏霞
(河北大学生命科学学院, 保定 071002)
泥鳅属(Misgurnus)、副泥鳅属(Paramisgurnus)隶属于鲤形目(Cypriniformes)、鳅科(Cobitidae), 喜栖息于静水或缓流水底部淤泥中。世界上泥鳅属鱼类有8种, 副泥鳅属有1种[1]。在中国泥鳅属分布有3种, 即泥鳅(Misgurnus anguillicaudatus)、黑龙江泥鳅(Misgurnus mohoity)和北方泥鳅(Misgurnus bipartitus)[2,3]。副泥鳅属有大鳞副泥鳅(Paramisgurnus dabryanus)。历年来关于北方泥鳅的有效性以及副泥鳅属的有效性存在争议。李雅娟等[4,5]基于微卫星标记及形态测量学分析, 认为北方泥鳅为有效种, 且与泥鳅亲缘关系较近, 而与黑龙江泥鳅亲缘关系较远。而Perdices等[6]利用Cytb和Rag1作为分子标记对鳅属(Cobitis)与泥鳅属的研究中, 认为北方泥鳅是黑龙江泥鳅的次定同物异名, 副泥鳅属为泥鳅属的次定同属异名。Yi 等[7]基于形态测量学与DNA条形码对泥鳅属鱼类及其邻近种的研究中, 认为北方泥鳅应为有效种。为了理清中国泥鳅属鱼类的物种多样性以及副泥鳅属鱼类的分类地位, 本研究基于外部形态特征及内部解剖特征, 结合线粒体基因(Cytb和COⅠ)序列, 对中国的泥鳅属及副泥鳅属鱼类进行了分类整理。
1 材料与方法
1.1 实验材料
2017年8—10月, 采用地笼捕捞方式, 采自重庆市丰都县(29°86′N, 107°69′E)、江西省南昌市(28°89′N, 116°01′E)、广东省东莞市(23°04′N,113°74′E)、内蒙古自治区赤峰市(42°34′N,118°72′E)、河北省易县(39°43′N, 115°18′E)、河北省雄安新区(38°87′N, 116°05′E)、湖南省平江县(29°41′N, 113°13′E)、湖北省通城县(29°26′N,113°83′E)、辽宁省岫岩县(40°26′N, 123°31′E)、浙江省杭州市(30°25′N, 119°73′E)、辽宁省凤城市(40°46′N, 124°06′E)、辽宁省沈阳市(41°72′N,123°31′E)、黑龙江省塔河县(52°32′N, 124°67′E)及宝清县(46°34′N, 132°25′E), 共计96尾, 标本浸泡于95%的乙醇中, 于-20℃低温保存在河北大学博物馆。
1.2 形态学分析
常规方法制作骨骼标本, 共解剖标本16尾, 其中泥鳅、北方泥鳅及黑龙江泥鳅各3尾, 大鳞副泥鳅7尾。采用数字游标卡尺测量(精确到0.1 mm)了23个可量性状, 测量方法参照陈咏霞等[8]的方法。使用连接到Leica M205A立体显微镜的Leica DFC295数码摄像头拍摄标本的脑颅骨骼、鳞片以及口唇结构等形态特征。
文中使用的英文缩写: HU (Hebei University,HU), 河北大学博物馆; SL (Standard Length, SL), 体长; HL (Head Length, HL), 头长; ML (Maximum Likelihood, ML), 最大似然法; BI (Bayesian Inferences, BI), 贝叶斯; sec (Supraethmoid-ethmoid complex), 上筛骨-中筛骨复合体; pv (Vomer), 梨骨;fr (Frontal), 额骨; pa (Parietal), 顶骨; so (Supraoccipital, 上枕骨; fon (Frontoparietal), 囟门; eo (Exoccipital), 外枕骨; oc (occipital condyle), 枕髁; bo (basioccipital), 基枕骨; ps (Parasptenoid), 副蝶骨。
1.3 DNA的提取、序列扩增及测定
采用天根生化科技有限公司生产的细胞/组织基因组DNA提取试剂盒(离心柱型)提取纯化基因组DNA。扩增线粒体Cytb的引物为L (5′-GACTT GAAAAACCACCGTTG-3′) 和 H (5′-CTCCGAT CTCCGGATTACAAGAC-3′)[9]。线粒体COⅠ的引物为F (5′-TCAACCAACCACAAAGACATTGGC AC-3′)和R (5′-TAGACTTCTGGGTG GCCAAAG AATCA-3′)。扩增体系为: 1.0 μmol/L的正反引物各2 μL, DNA模版2 μL, 2×TaqMaster Mix Loading Dye 25 μL, ddH2O 17 μL。PCR反应条件为: 95℃预变性2min, 95℃变性30s, 55/51℃退火30s, 72℃延伸30s, 循环35次; 72℃延伸10min, 4℃终止反应。PCR产物经质量分数1.2%的琼脂糖凝胶电泳检测后, 由安徽通用生物系统有限公司进行双向测通。
1.4 基因序列信息及数据分析
从GenBank下载泥鳅、北方泥鳅、黑龙江泥鳅、俄罗斯泥鳅(Misgurnus mohoity)及大鳞副泥鳅的同源序列, 其中COⅠ基因18条, Cytb47条(表1和表2)。外类群的选取参考文献[6], 选取与泥鳅属亲缘关系较近的潘鳅属(Pangio)为外类群。使用BioEdit7.2.5软件进行同源序列拼接、排列及手动矫正。利用MEGA5.0[10]软件对序列进行比对, 去除两端冗余序列。用DnaSP5.10[11]软件确定序列单倍型, 采用最大似然法(ML)和贝叶斯法(BI)构建系统进化树, 用jModelTest2.1.4[12]软件选择最适合的进化模型, 替代模型为GTR+G+I。运用RAxML[13]软件构建ML分子系统树, 用MrBayes3.0软件[14]构建BI分子系统树。
2 结果
2.1 中国泥鳅属(Misgurnus)鱼类检索表
1a.基枕骨末端分叉或愈合不完全; 额骨细、长; 口角须末端未达鳃盖后缘; 鳞片小......................2
1b.基枕骨末端愈合完全; 额骨宽、短; 口角须末端延伸至鳃盖后缘; 鳞片大................................................................................大鳞副泥鳅M.dabryanus
2a.腹鳍起点与背鳍第2—3根分枝鳍条相对;鳞焦小; 上筛骨-中筛骨复合体前端尖锐; 梨骨呈蝶形................................................................................3
2b.腹鳍起点与背鳍起点相对或稍后; 鳞焦大;上筛骨—中筛骨复合体前端钝圆; 梨骨呈梨形................................................黑龙江泥鳅M.mohoity
3a.须长, 口角须末端延伸至或超过眼前缘; 尾柄短, 雄性, 尾柄长为尾柄高的1.3—1.7倍(平均1.6),雌性为1.2—1.7倍(平均1.6).................................................................泥鳅M.anguillicaudatus
3b.须短, 口角须末端不达眼前缘; 尾柄长, 雄性, 尾柄长为尾柄高的2.3—3.4倍(平均2.9), 雌性为2.2—3.5倍(平均2.9).................................................................................................北方泥鳅M.bipartitus
泥鳅属Misgurnus Lacépède, 1803
MesomisgurnusFang, 1935, 6(2): 128—146.
UssuriaNikolskii, 1903: 362.
ParamisgurnusDabry de Thiersant, 1872: 191
模式种:Misgurnus fossilisLinnaeus
体延长; 须3对, 其中吻须2对, 口角须1对; 颏叶发达, 通常呈须状; 体被细鳞, 位于皮下, 头部无鳞。体鳞小, 鳞焦小, 位于基部; 眼间距大于眼径。腹鳍起点与背鳍起点相对或靠后; 基枕骨末端在背大动脉的腹面分叉或愈合。
泥鳅 Misgurnus anguillicaudatus (Cantor, 1842)(图1—3)
检视标本: HU1810435、1708195和1708196,3尾(1尾雄性, 2尾雌性), 体长97.8—108.2 mm, 采自重庆市丰都县名山长江流域; HU1810450、1810452、1810451和1810453, 4尾(2尾雄性, 2尾雌性), 体长66.8—71.4 mm, 采自江西省南昌市赣江; HU1608505、1204021、1204026、1608506和1708244, 5尾(3尾雄性, 2尾雌性), 体长61.4—79.3 mm, 采自广东省东莞市东莞运河。
鉴别特征: 区别与泥鳅属其它物种的特征为:体略粗壮, 颏叶发达, 呈须状, 内颏叶长度约为外颏叶的1/2, 内颏叶长度略小于眼径(图2A); 须较长,口角须末端延伸至或超过眼后缘; 尾柄短, 雄性尾柄长为尾柄高的1.3—1.7(平均1.6)倍, 雌性为1.2—1.7(平均1.6)倍; 鳞焦极小, 鳞焦直径小于基部环片到鳞焦的距离, 具22—24个初生辐射沟(图2B)。
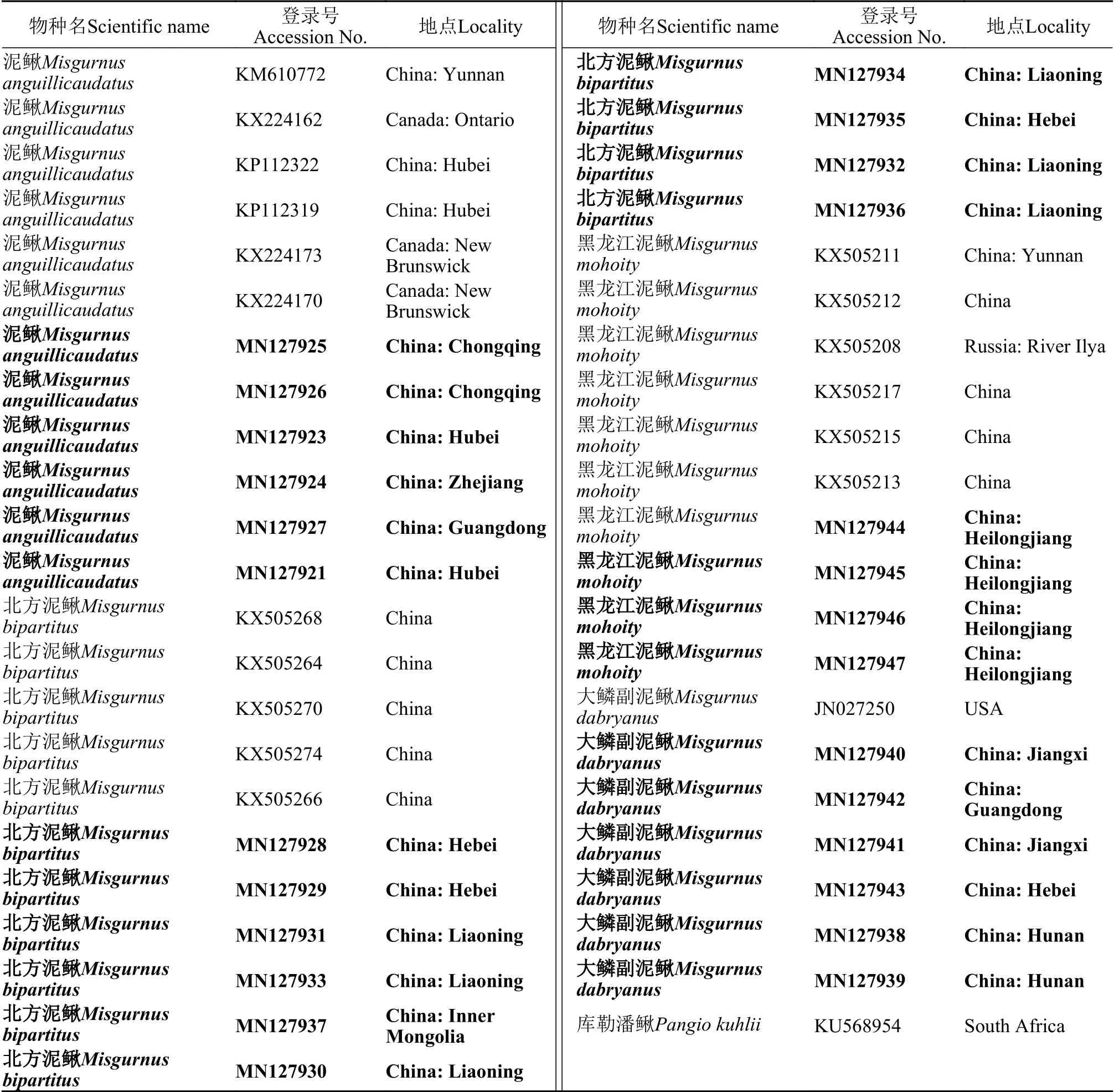
表1 本研究中用到COⅠ的数据来源Tab.1 Source of data for COⅠ used in this study
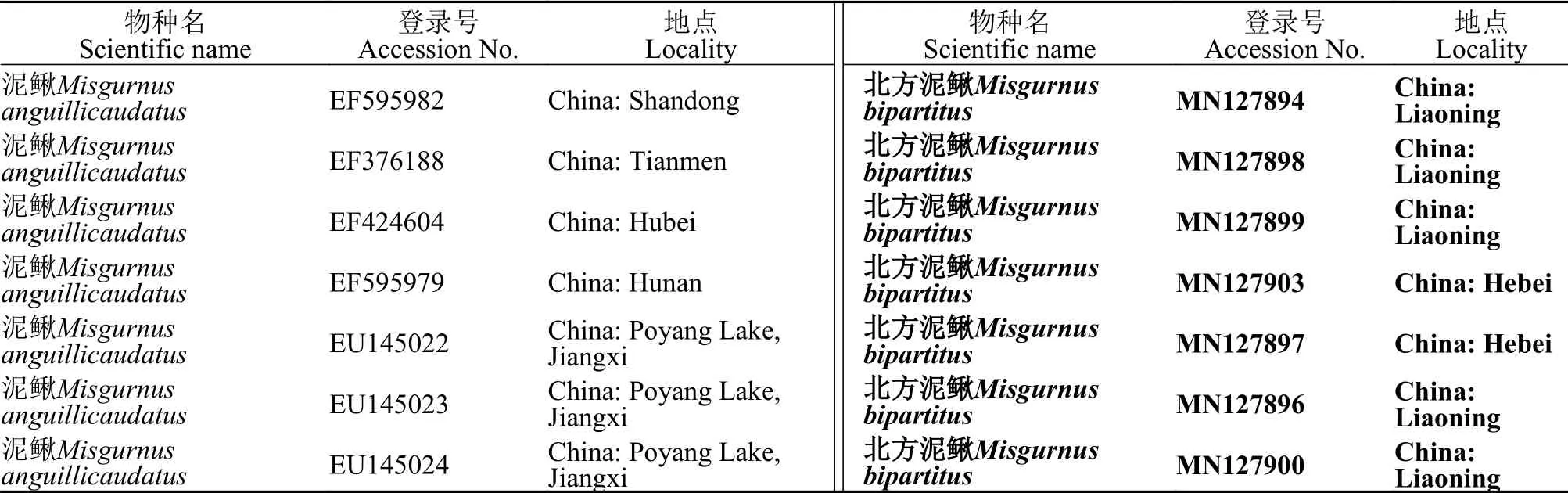
表2 本研究中用到Cyt b的数据来源Tab.2 Source of data for Cyt b used in this study
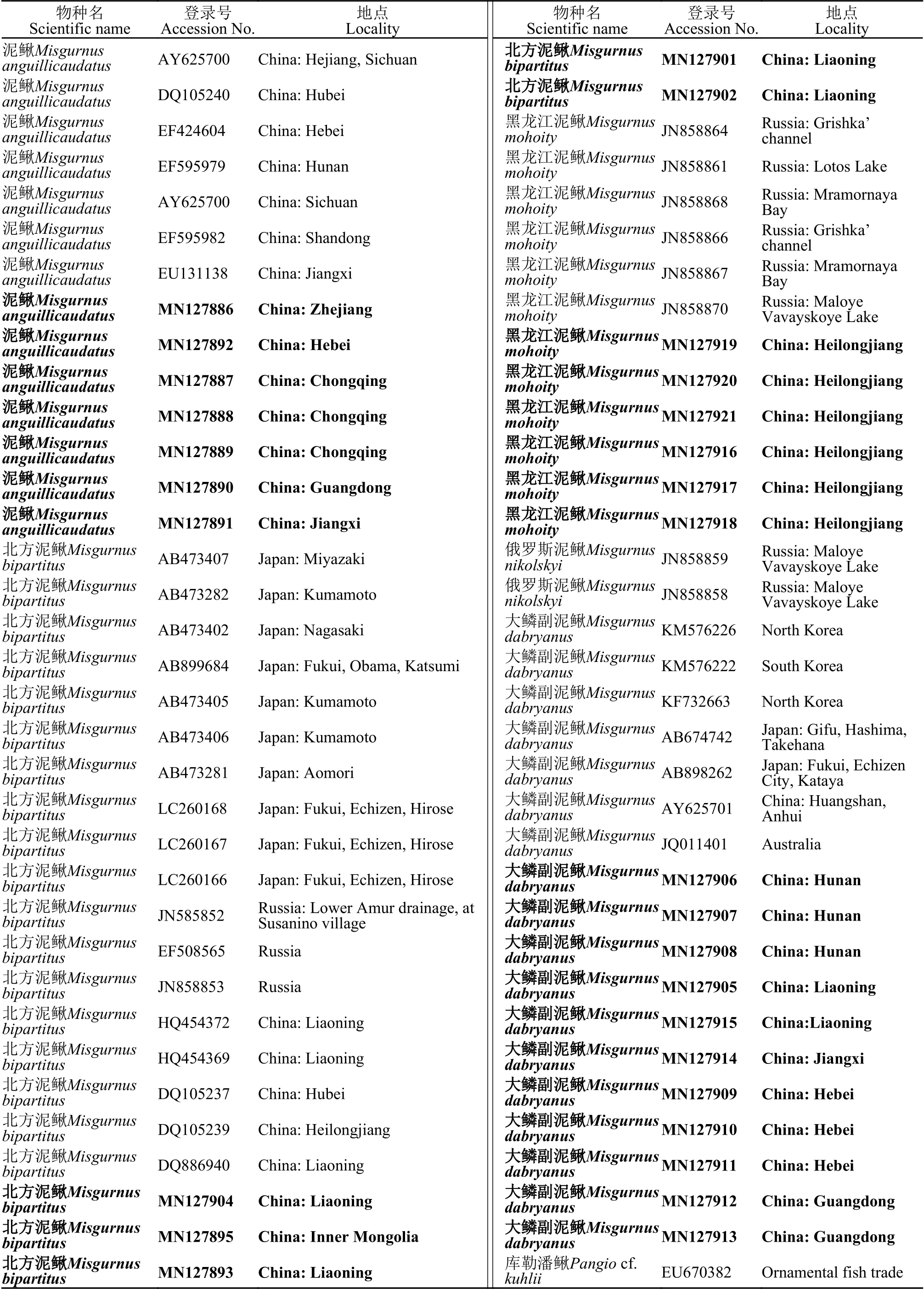
续表2

图1 泥鳅(A.HU1810451 , 66.8 mm; B.HU1810452 ♀,76.5 mm; 采自江西省南昌市;比例尺: A、B=1cm)Fig.1 Misgurnus anguillicaudatus (A.HU1810451 , 66.8 mm;B.HU1810452 ♀, 76.5 mm.China: Nanchang City, Jiangxi Prov.;Scale bars: A, B=1 cm)
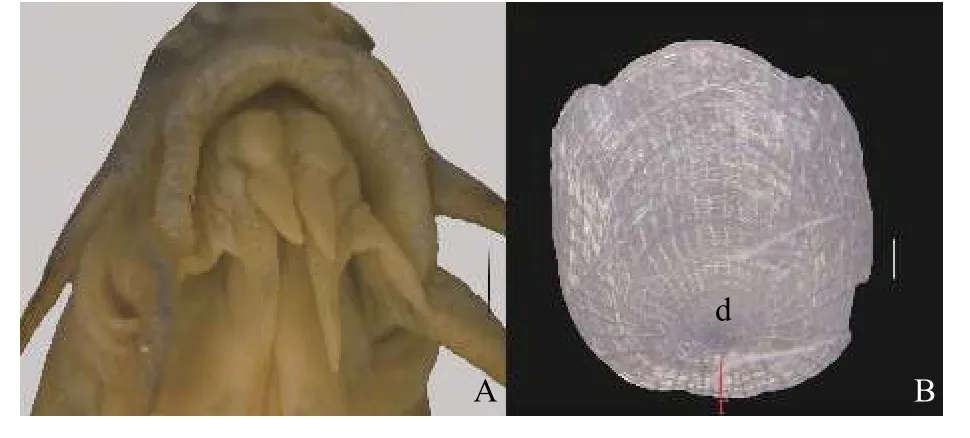
图2 泥鳅(A、B.HU1810451, , 66.8 mm; 采自江西省南昌市.d.鳞焦直径, r.基部环片到鳞焦的长度.比例尺: A=1 mm; B=0.1 mm)Fig.2 Misgurnus anguillicaudatus (A, B.HU1810451, , 66.8 mm.China: Nanchang City, Jiangxi Prov.; d.length of scale focal;r.length of base scale ridges to scale focal.Scale bars: A=1 mm;B=0.1 mm)
形态描述: 测量数据见表3。背鳍Ⅳ, 6—8; 臀鳍Ⅲ, 5; 胸鳍I, 8—9, 腹鳍I, 5—6。
骨骼特征: 如图3所示, 上筛骨-中筛骨复合体(sec)侧面观前端延伸, 呈尖锐状; 梨骨(pv)腹面观呈蝶形; 额骨(fr)前端细长, 后端宽、短; 由额骨、顶骨(pa)及上枕骨(so)围成的囟门(fon)细、长; 外枕骨(eo)腹后缘未完全覆盖于枕髁(oc)上方; 副蝶骨(ps)近中央的两翼侧突前端略宽大, 与纵轴的夹角为较小的钝角; 基枕骨(bo)末端在背大动脉的腹面分叉。
分布范围: 在我国除西部高原外,各大水系均有分布。
北方泥鳅 Misgurnus bipartitus (Sauvage &Dabry de Thiersant, 1874)(图4—6)
检视标本: HU1608613、1609183、1708191、1708149、1608610、1609178、1708190、1708192和1 7 0 8 1 5 0, 9 尾(2 尾 雄 性, 7 尾 雌 性), 体 长54.2—109.5 mm, 采自辽宁省鞍山市岫岩县大洋河;HU1708178、1708176、1708177、1708181、1609422、1609399、1608601、1608606、1708182、1609403和1609423, 11尾(4尾雄性, 7尾雌性), 体长72.0—128.7 mm, 采自内蒙古赤峰, 西辽河上游阴河;HU1806804、1806836和1806825, 3尾(1尾雄性,2尾雌性), 体长85.3—94.8 mm, 采自河北省易县紫金关拒马河。
鉴别特征: 区别于泥鳅属其他物种的特征: 体纤细, 颏叶不发达, 内颏叶短小, 呈纽扣状; 外颏叶略长(图5A); 须短, 口角须末端不达眼前缘; 尾柄长, 雄性尾柄长为尾柄高的2.3—3.4(平均2.9)倍, 雌性为2.2—3.5(平均2.9)倍; 鳞焦较小, 鳞焦直径约等于基部环片到鳞焦的距离, 具有23—24个初生辐射沟(图5B)。
形态描述: 测量数据见表4。背鳍Ⅳ, 6; 臀鳍Ⅲ, 5; 胸鳍Ⅰ, 8—9, 腹鳍Ⅰ, 5—6。
骨骼特征: 如图6所示, 上筛骨-中筛骨复合体侧面观前端延伸, 呈尖锐状; 梨骨腹面观呈蝶形; 额骨前端细长, 后端宽、短; 囟门细、长; 外枕骨腹后缘完全覆盖于枕髁上方; 副蝶骨的两翼侧突前端窄小, 与纵轴的夹角近似直角; 基枕骨末端在背大动脉的腹面分叉。
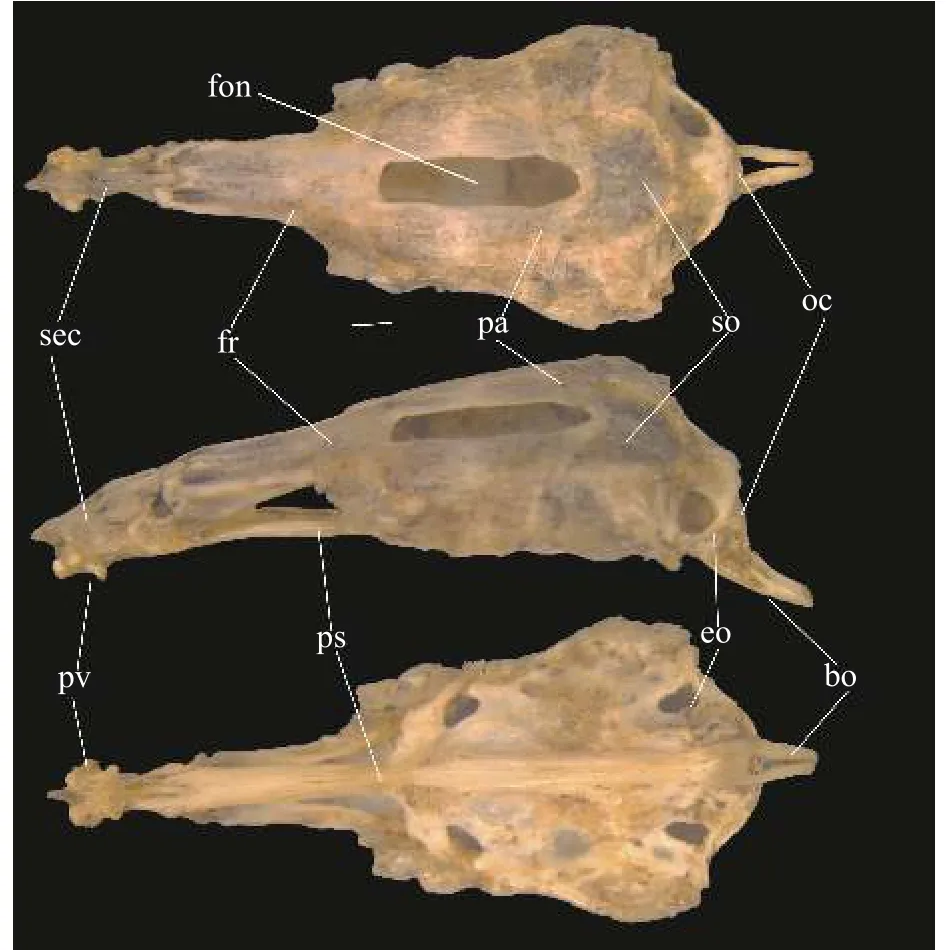
图3 泥鳅脑颅, 从上到下依次为背面观、侧面观、腹面观Fig.3 Cerebral cranium of Misgurnus anguillicaudatus, from top to bottom, the dorsal view, the lateral view and the ventral view
表3 泥鳅的形态测量数据Tab.3 Morphometric data of Misgurnus anguillicaudatus

图4 北方泥鳅(A.HU1708176 ♂, 96.5 mm; B.HU1608601 ♀,72.0 mm; 采自内蒙古自治区赤峰.比例尺: A、B=1 cm)Fig.4 Misgurnus bipartitus (A.HU1708176 ♂, 96.5 mm; B.HU1608601 ♀, 72.0 mm.China: Chifeng City, Inner Mongolia.Scale bars: A, B=1 cm)
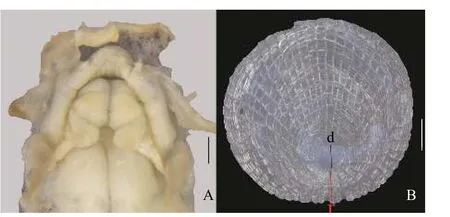
图5 北方泥鳅(A、B.HU1708176 ♂, 96.5 mm; 采自内蒙古自治区赤峰.比例尺: A= 1 mm, B= 0.1 mm)Fig.5 Misgurnus bipartitus (A, B.HU1708176 ♂, 96.5 mm.China: Chifeng City, Inner Mongolia.Scale bars: A=1 mm, B=0.1 mm)
分布范围: 分布于海河以北的大部分水系。
黑龙江泥鳅 Misgurnus mohoity (Dybowski,1869)(图7—9)
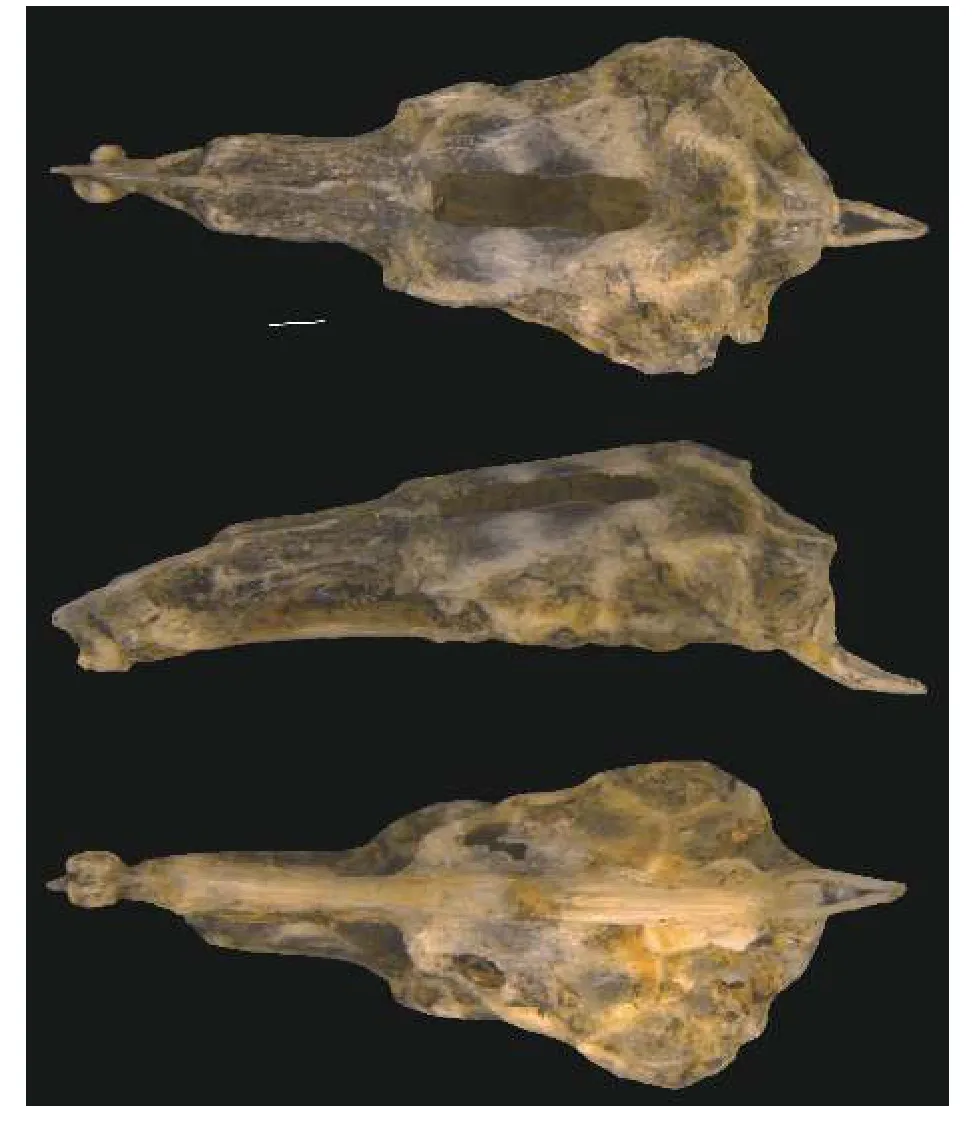
图6 北方泥鳅脑颅 (从上到下依次为背面观、侧面观、腹面观.比例尺=1 mm)Fig.6 Cerebral cranium of Misgurnus bipartitus (from top to bottom, the dorsal view, the lateral view and the ventral view.Scale bars=1 mm)
检视标本: HU1609381、1708194、1708147、1690385、1609382、1608457、1608388、1608463、1609387、1708160、1708102、1804029、1608385、1708148、1708163、1708159和1609386, 17尾(7尾雄性, 10尾雌性), 体长84.6—152.6 mm, 采自黑龙江塔河县的呼玛河; HU1609387、1608453、1708163、1608454、1608462、1608452、1608455、1804032、1804028、1804031、1608456、1608387和1708161,13尾(4尾雄性, 9尾雌性),体长68.9—143.8 mm, 采自黑龙江宝清县的饶力河。
鉴别特征: 区别于泥鳅属其他物种的特征: 体中等, 颏叶略发达, 呈须状, 内颏叶长度大于眼径(图8A); 腹鳍起点与背鳍起点相对或略靠后; 体鳞,鳞焦较大, 鳞焦直径大于基部环片至鳞焦的距离,具有28—29个初生辐射沟(图8B); 梨骨呈梨形; 基枕骨末端在背大动脉的腹面愈合不完全。
形态描述: 测量数据见表5。背鳍Ⅳ, 6; 臀鳍Ⅲ, 5; 胸鳍Ⅲ, 8—10; 腹鳍I, 5—6。
骨骼特征: 如图9所示, 上筛骨-中筛骨复合体侧面观前端短, 未延伸, 呈钝圆; 梨骨腹面观呈梨形;额骨前端细长, 后端宽、短; 囟门细、长; 外枕骨腹后未覆盖于枕髁前上方; 副蝶骨的两翼侧突前端略宽大, 与纵轴的夹角为较大的钝角; 基枕骨末端在背大动脉的腹面愈合不完全。
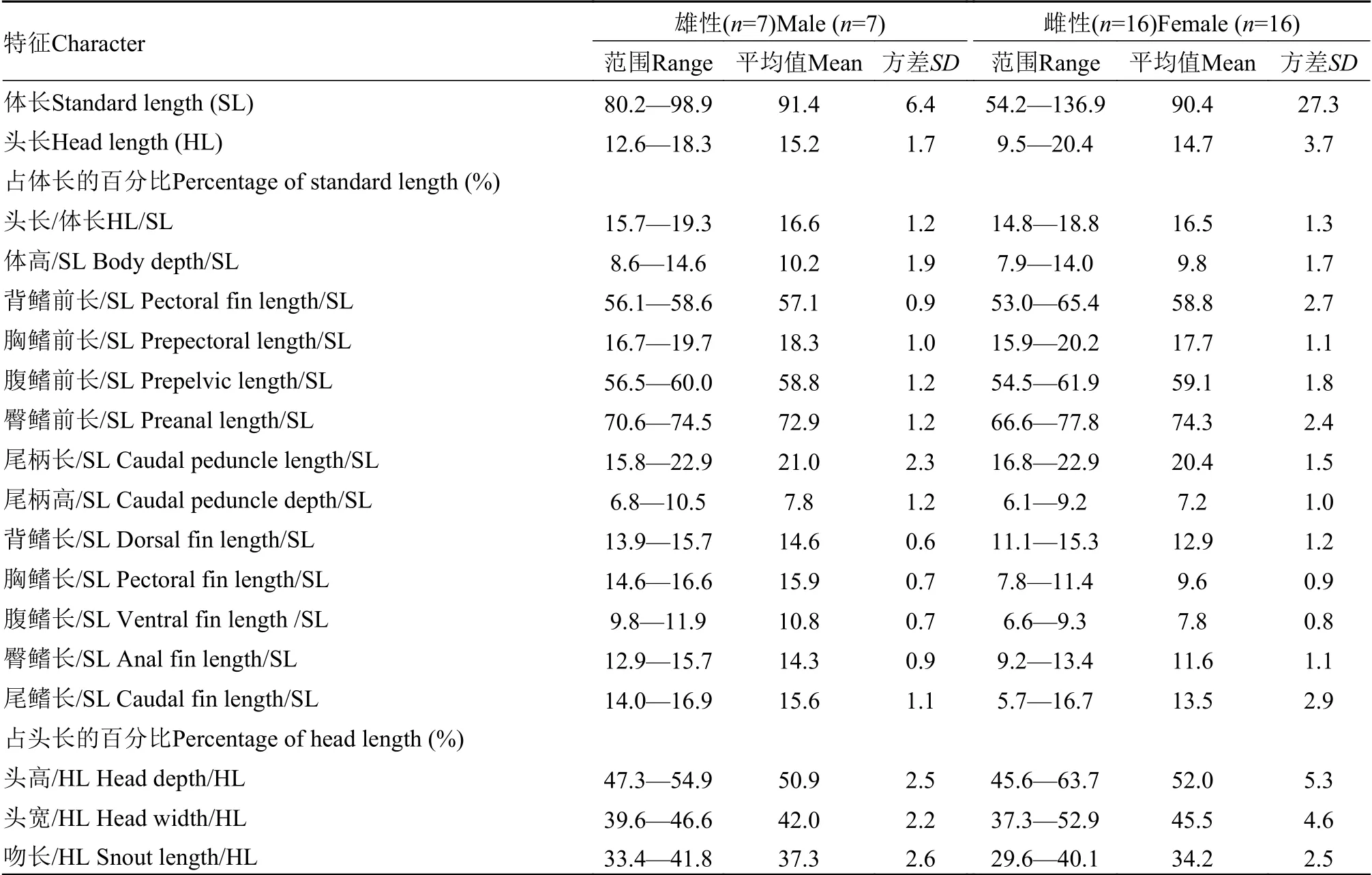
表4 北方泥鳅的形态测量数据Tab.4 Morphometric data of Misgurnus bipartitus
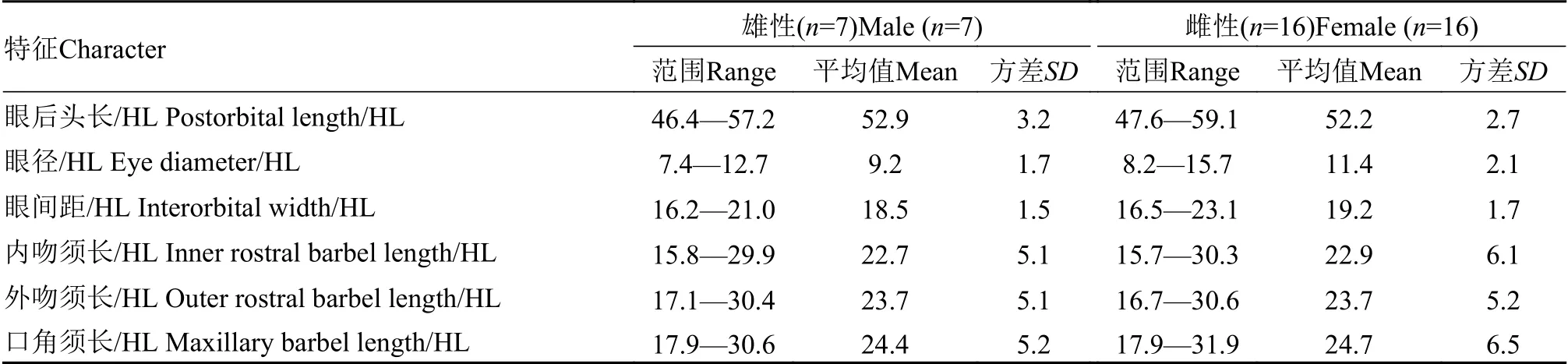
续表4

图7 黑龙江泥鳅(A.HU1708103 116.4 mm; B.HU1608387♀, 143.8 mm; 采自黑龙江省宝清县和塔河县; 比例尺: A、B=1 cm)Fig.7 Misgurnus mohoity (A.HU1708103 , 116.4 mm; B.HU1608387 ♀, 143.8 mm.China: Baoqing and Tahe County,Heilongjiang Prov.; Scale bars: A, B=1 cm)

图8 黑龙江泥鳅(A、B.HU1608457 , 132.4 mm; 采自黑龙江省宝清县和塔河县.比例尺: A=1mm, B=0.1 mm)Fig.8 Misgurnus mohoity (A, B.HU1608457, 132.4 mm.China: Baoqing and Tahe County, Heilongjiang Prov.; Scale bars:A=1 mm, B=0.1 mm)
分布范围: 分布于黑龙江流域。
大鳞副泥鳅 Misgurnus dabryanus (Dabry de Thiersant, 1872)(图10—12)
检视标本: HU1810438、1708239、1708238、1810443、1810444、1810445、1810446、1810449、1708240、1810433、1810439、1810447、1800448和1608421, 14尾(9尾雄性, 5尾雌性), 体长54.8—111.4 mm, 采自雄安新区白洋淀(38°87′N, 116°05′E);HU1608424、1608425和1608423, 3尾(1尾雄性,2尾雌性), 体长79.9—80.4 mm, 采自湖南省岳阳市平江县湘江; HU1608425、1608423、1608448和1608435, 4尾(1尾雄性, 3尾雌性), 体长79.9—102.1 mm,采自湖北省咸宁市通城陆水; HU1810436、1810437、1810434、1608605、1708151和1609189, 6尾(1尾雄性, 5尾雌性), 体长42.4—107.0 mm, 采自辽宁省鞍山市岫岩县大洋河。
鉴别特征: 体粗壮, 颏叶发达, 呈长须状, 内颏叶长度大于眼径(图11A); 须长, 口角须末端延伸至鳃盖后缘; 吻短, 雄性吻长为头长的26.4%—55.8%(平均34.0%), 雌性为29.8%—41.7%(平均34.6%); 尾柄长, 雄性尾柄长为尾柄高的1.3—1.8(平均1.6)倍,雌性为1.2—2.0(平均1.5)倍; 皮质棱发达; 体鳞大,鳞焦小, 鳞焦直径小于基部环片至鳞焦距离的1/2,具有28—30个初生辐射沟(图11B); 基枕骨末端在背大动脉的腹面愈合完全。
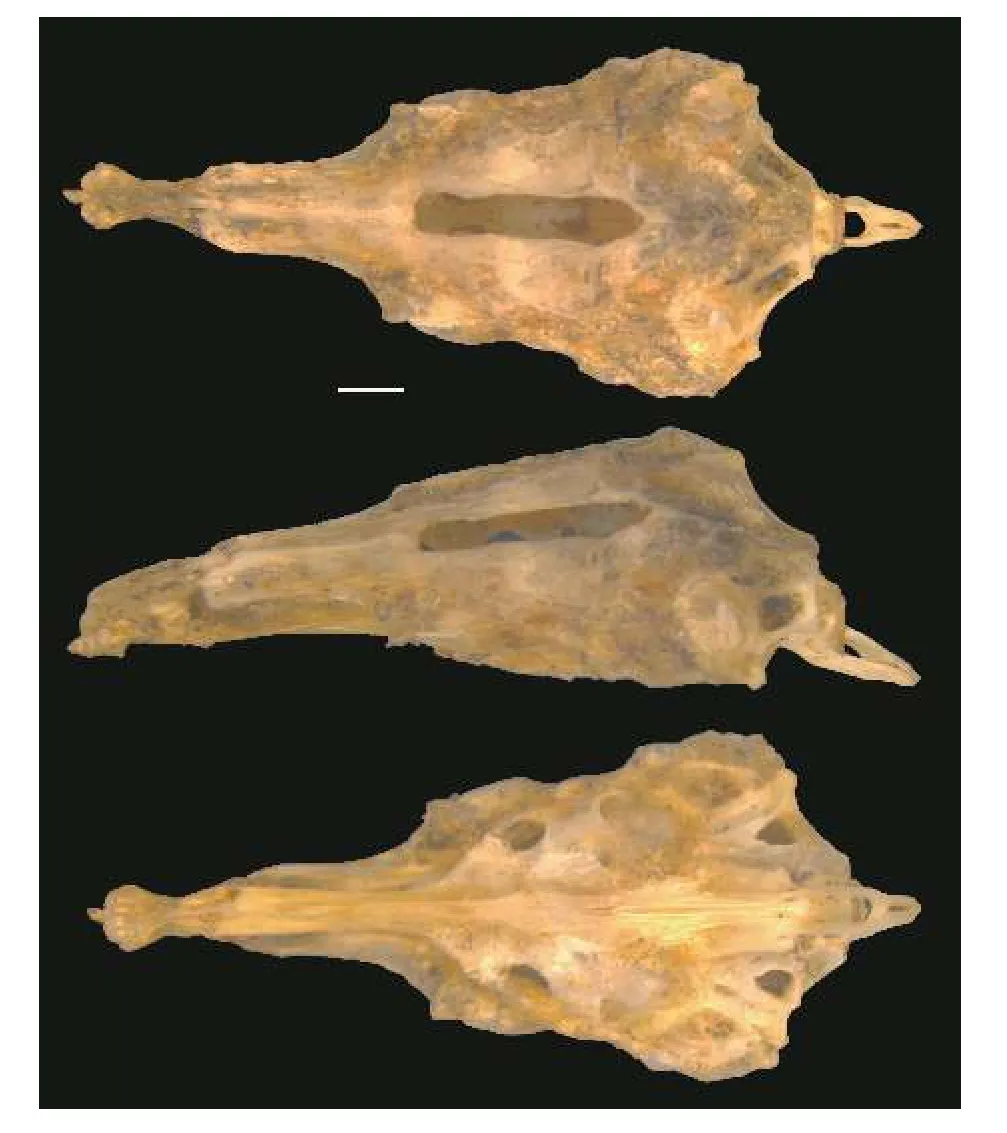
图9 黑龙江泥鳅脑颅 (从上到下依次为背面观、侧面观、腹面观;比例尺=1 mm)Fig.9 Cerebral cranium of Misgurnus mohoity (from top to bottom, the dorsal view, the lateral view and the ventral view;Scale bars=1 mm)
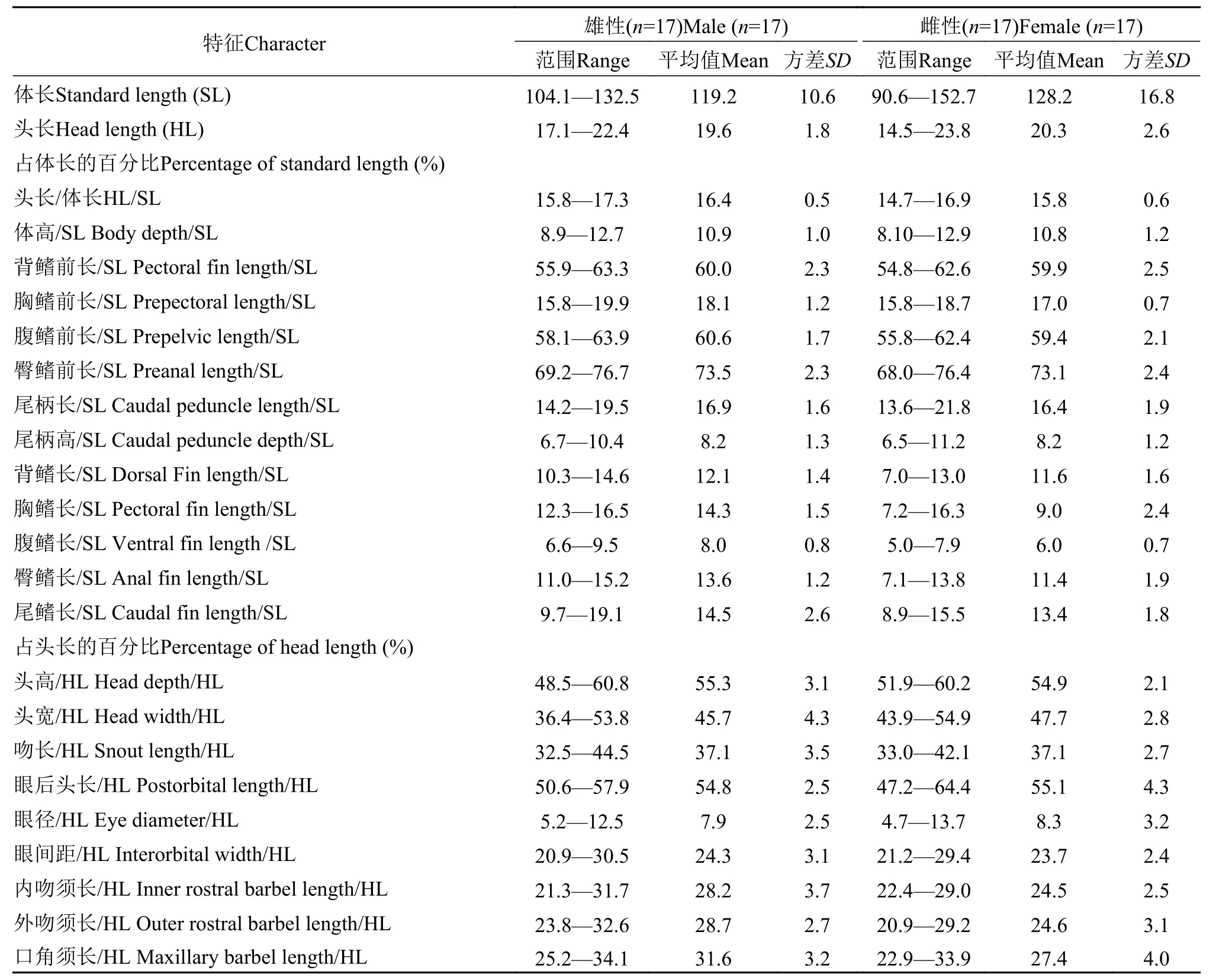
表5 黑龙江泥鳅的形态测量数据Tab.5 Morphometric data of Misgurnus mohoity

图10 大鳞副泥鳅(A.HU1810444 , 86.2 mm; B.HU1810448♀, 62.6 mm; 采自河北省安新县白洋淀.比例尺: A、B=1 cm)Fig.10 Misgurnus dabryanus (A.HU1810444 , 86.2 mm;B.HU1810448 ♀, 62.6 mm.China: Baiyangdian, Hebei Prov.;Scale bars: A, B=1 cm)
形态描述: 测量数据见表6。背鳍Ⅳ, 6—8; 臀鳍Ⅲ, 5; 胸鳍Ⅰ, 9—10, 腹鳍Ⅰ, 5—6。
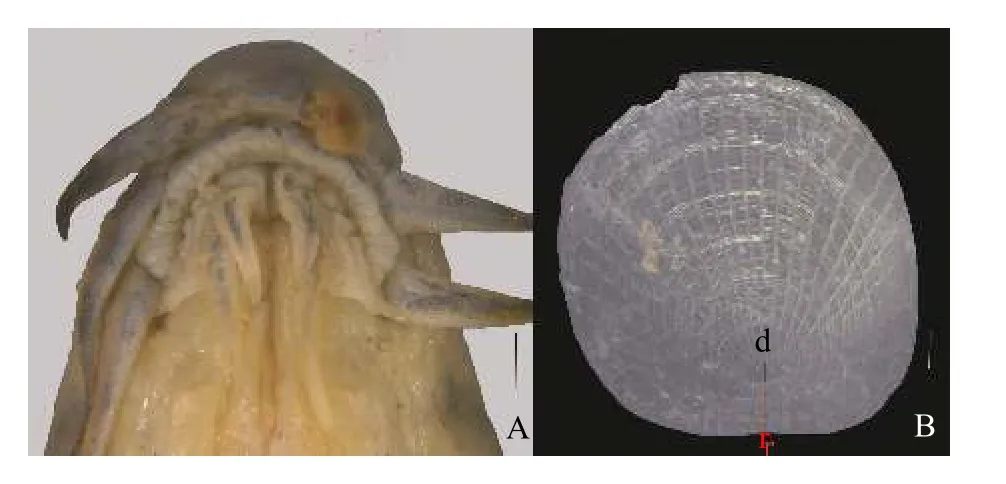
图11 大鳞副泥鳅(A、B.HU1810444 , 86.2 mm; 采自河北省安新县白洋淀.比例尺: A=1 mm, B=0.1 mm)Fig.11 Misgurnus dabryanus (A, B.HU1810444 , 86.2 mm.China: Baiyangdian, Hebei Prov.; Scale bars: A=1 mm, B=0.1 mm)
骨骼特征: 如图12所示, 上筛骨-中筛骨复合体侧面观前端延伸, 呈尖锐状; 梨骨腹面观呈梨形; 额骨宽、短, 前端窄于后端; 囟门宽、短, 呈长方形;外枕骨腹后缘未覆盖于枕髁前上方; 副蝶骨的两翼侧突前端宽大, 与纵轴的夹角近似直角; 基枕骨末端在背大动脉的腹面末端愈合完全。
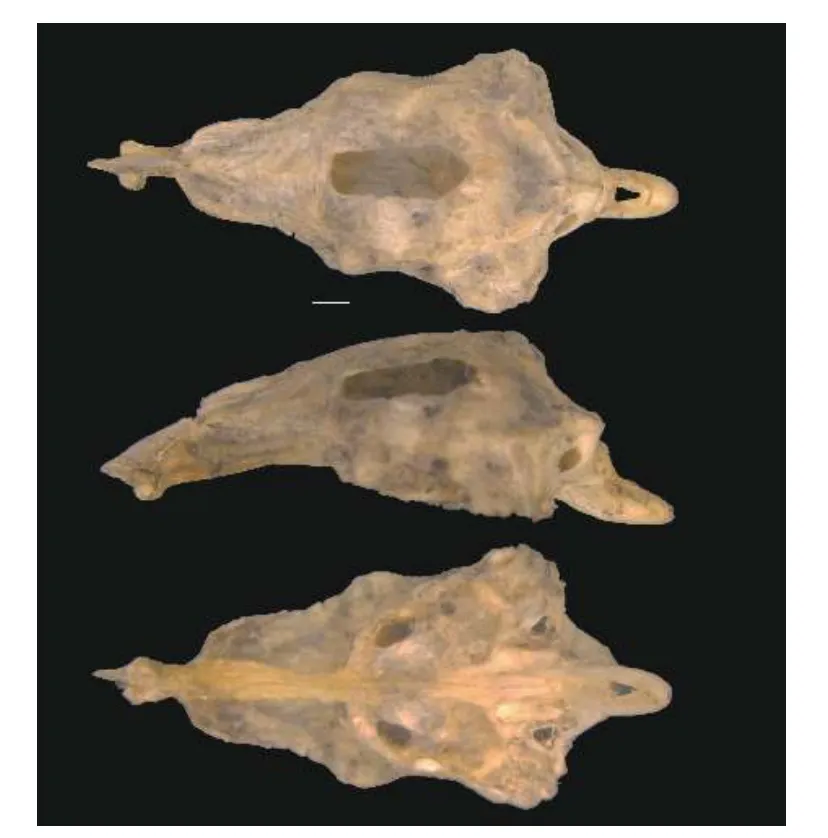
图12 大鳞副泥鳅脑颅(从上到下依次为背面观、侧面观、腹面观;比例尺=1 mm)Fig.12 Cerebral cranium of Misgurnus dabryanus (from top to bottom, the dorsal view, the lateral view and the ventral view;Scale bars=1 mm)
分布范围: 在我国除西部高原外,各大水系均有分布。
2.2 基于COⅠ与Cyt b基因的系统进化分析
将GenBank下载的18条COⅠ序列与实验所得数据一并分析, 共88条, 单倍型44个, 单倍型多样性Hd=0.9711。COⅠ基因序列长度为632 bp,其中, 保守位点(446)个, 占序列全长的70.6%, 变异位点(V)186个, 占29.4%, 简约信息位点(Pi)157个, 占24.8%。序列的平均碱基组成为T=32.3, C=26.6,A=23.5, G=17.7, 碱基组成表现出明显的T偏倚; 转换位点50个, 颠换位点19个, 转换与颠换的比值为(Ts/Tv)2.59。基于Kimura双参数进化模型计算的遗传距离显示, 泥鳅属和副泥鳅属属间遗传距离为14.4%—18.6%, 泥鳅属种间遗传距离10.4%—19.6%;在4个物种中, 泥鳅与北方泥鳅遗传距离最小, 为10.4%; 泥鳅与黑龙江泥鳅遗传距离最大, 为19.6%(表7)。基于COⅠ基因构建的贝叶斯树(BI)和最大似然树(ML)拓扑结构一致(图13), 泥鳅、北方泥鳅、黑龙江泥鳅及大鳞副泥鳅的个体均以高置信度聚为一枝, 其中泥鳅与北方泥鳅构成姐妹群, 再与黑龙江泥鳅聚在一起, 大鳞副泥鳅位于树的根部。
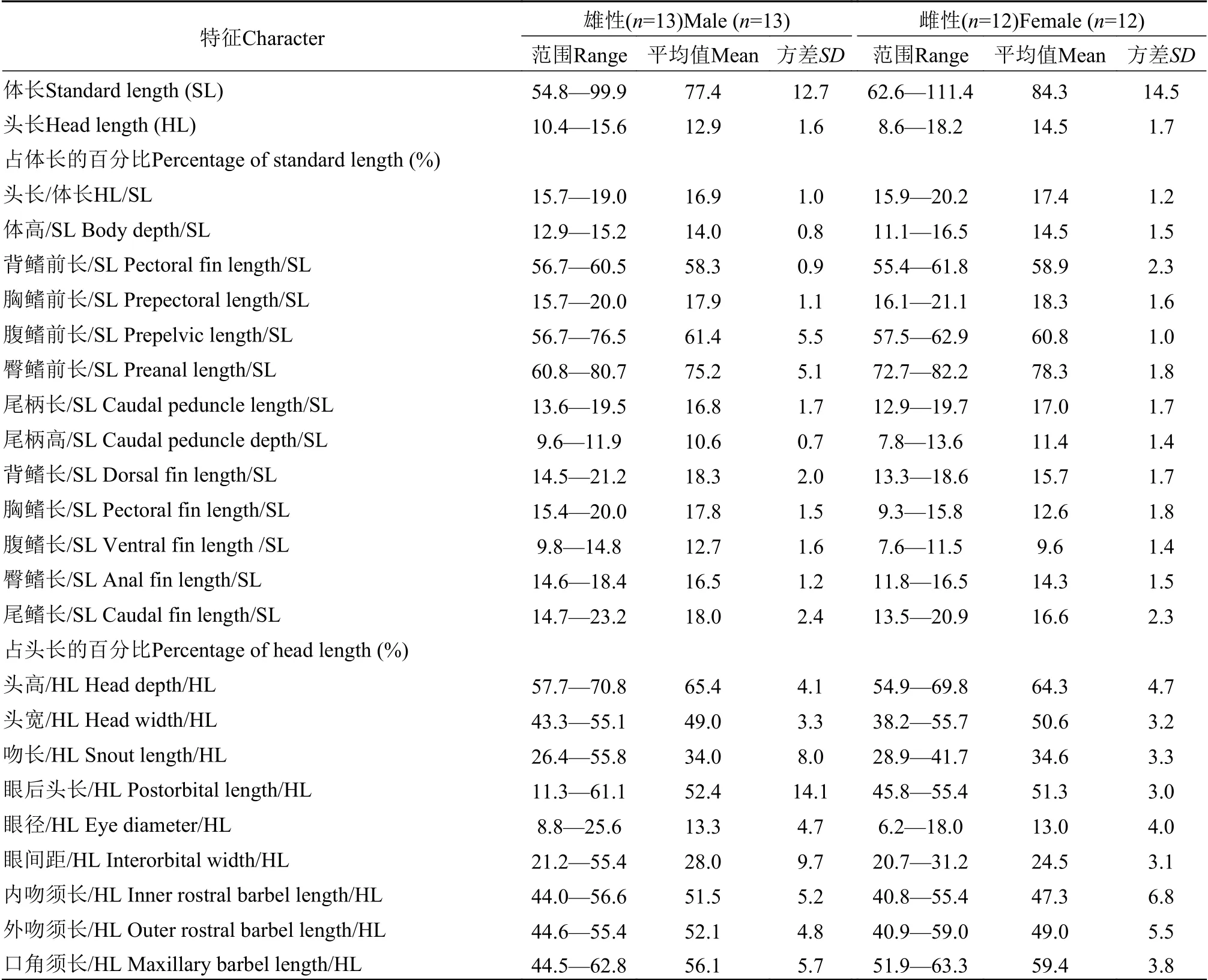
表6 大鳞副泥鳅的形态测量数据Tab.6 Morphometric data of Misgurnus dabryanus
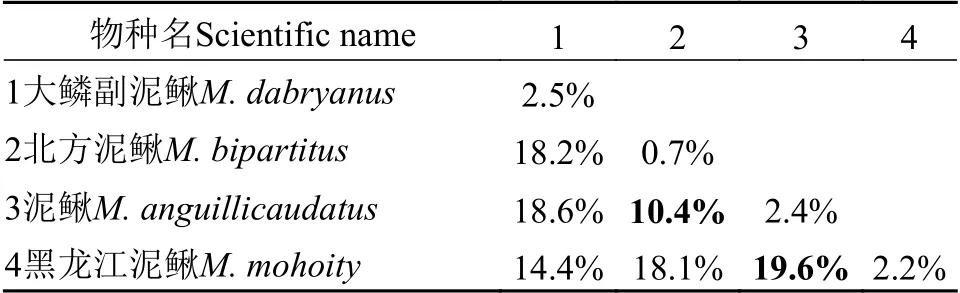
表7 基于COⅠ基因泥鳅属种间遗传距离与种内遗传距离Tab.7 Genetic distance between species of Misgurnus based on the COⅠ gene
Cytb序列共94条, 单倍型80个, 单倍型多样性Hd=0.9945.Cytb基因序列长度为1113 bp,其中, 保守位点(C)697个, 占序列全长的62.6%, 变异位点(V)416个, 占37.4%, 简约信息位点(Pi)439个, 占39.4%, 序列的平均碱基组成为T=31.2, C=26.8,A=27.0, G=14.9, 碱基组成表现出明显的T偏倚; 转换位点92个, 颠换位点32个, 转换与颠换的比值为(Ts/Tv)2.7。基于Kimura双参数进化模型计算的遗传距离显示, 泥鳅属和副泥鳅属属间遗传距离为15.4%—19.5%, 泥鳅属种间遗传距离为 11.9%—19.3%; 4个物种间, 泥鳅与北方泥鳅的遗传距离最小, 为11.9%; 泥鳅与大鳞副泥鳅的遗传距离最大,为19.5%(表8)。基于Cytb基因以及基因联合构建的贝叶斯树(BI)和最大似然树(ML)拓扑结构一致(图14), 与COⅠ基因进化树不同。泥鳅、北方泥鳅和大鳞副泥鳅亲缘关系近, 黑龙江泥鳅位于树的基部。
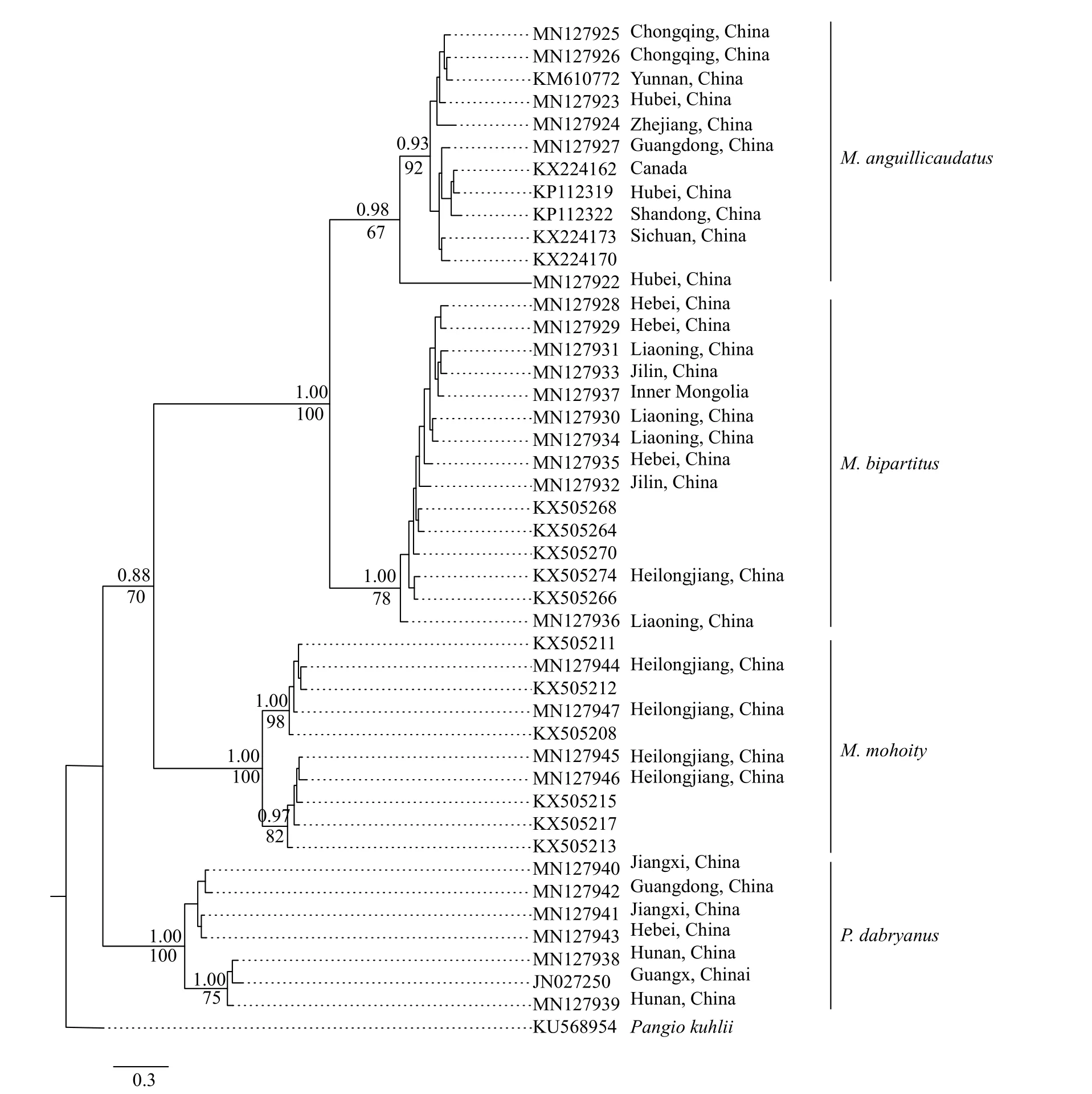
图13 基于线粒体COⅠ基因构建的BI树和ML树的合意树Fig.13 Bayesian tree for Misgurnus inferred from mitochondrial COⅠgene sequences
联合Cytb和COⅠ基因计算泥鳅属和副泥鳅属属间遗传距离为15.2%—17.5%, 泥鳅属种间遗传距离为10.7%—18.4%。基因联合基因构建的BI树和ML树与Cytb基因树拓扑结构一致。
3 讨论
3.1 泥鳅属与副泥鳅属的分化及属的有效性
Mary等[15]认为属的主要特性是形态学上的独特性, 它与其他相似的单元之间存在着一个确定的间断。陈景星[2]认为在骨骼特征及外部形态特征上, 副泥鳅属区别于泥鳅属鱼类的特征为基枕骨末端在背大动脉腹下愈合, 须长, 鳞大, 尾柄皮质棱发达等。本文研究显示, 基枕骨末端在背大动脉腹下结构特征从分叉到愈合是一个逐渐演化的过程, 如北方泥鳅和泥鳅基枕骨末端分叉, 黑龙江泥鳅为不完全愈合, 大鳞副泥鳅为完全愈合。同样, 须的长度也是逐渐延长的过程, 北方泥鳅最短, 其次是黑龙江泥鳅、泥鳅和大鳞副泥鳅。虽然Nalbant[16]认为鳞片大小和形状, 鳞焦大小和位置, 及辐射沟特征等在鳅科不同属中存在差异, 但在泥鳅属和副泥鳅属4个物种中, 鳞片差异属于种间差异, 如泥鳅和大鳞副泥鳅鳞焦小, 北方泥鳅和黑龙江泥鳅鳞焦大。在鳅科鱼类中, 第二性征通常是属的主要鉴别特征, 如鳅属(Cobitis)在雄性胸鳍第二根鳍条基部具一骨质突起(Lamina circularis); 双须鳅(Bibarba)在雄性第二和第三根鳍条上具骨质突起; 萨瓦纳鳅属(Sabanejewia)在繁殖季节, 体侧垂直膨胀(Vertical swellings)。泥鳅属和副泥鳅属第二性征相同,雄性胸鳍第二根鳍条变长变宽, 在繁殖季节, 雄性身体两侧水平膨胀(Horizontal swellings)。综上所述, 基于形态特征副泥鳅属的有效性并没有得到很好的支持。
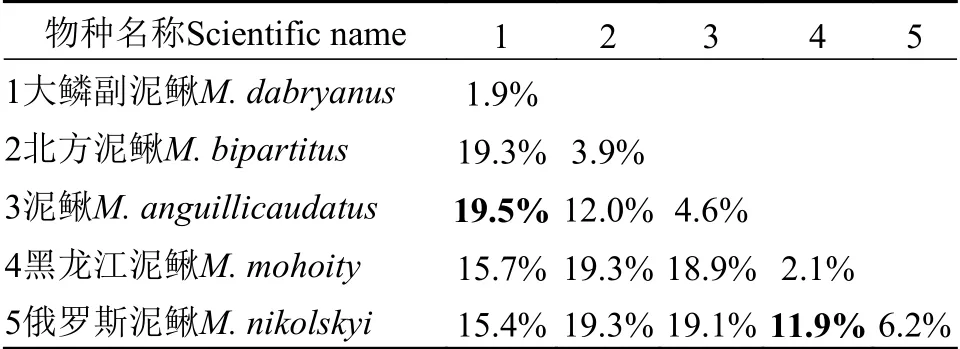
表8 基于Cyt b基因泥鳅属种间遗传距离与种内遗传距离Tab.8 Genetic distance between species of Misgurnus based on the Cyt b gene
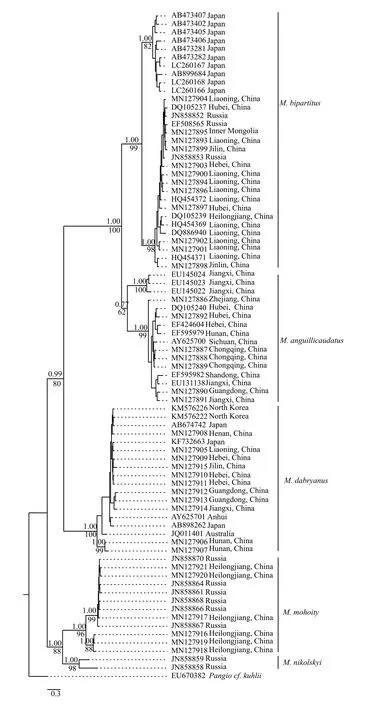
图14 基于线粒体基因Cyt b构建的贝叶斯和ML的合一系统发育树Fig.14 Bayesian tree for Misgurnus inferred from mitochondrial Cyt b gene sequences
基于分子标记分析副泥鳅属的有效性也没有得到很好的支持。Šlechtová等[17]利用Cytb和Rag1作为分子标记否定了副泥鳅属的有效性。Perdices等[6]利用Cytb和Rag1作为分子标记支持这一结论, 认为其为泥鳅属的次定同属异名。本研究基于线粒体Cytb和COⅠ基因的分析中, 副泥鳅属的有效性也没有得到很好的支持。在遗传距离上, 副泥鳅属与泥鳅属的遗传距离与泥鳅属种间距离相似; 在分子进化树中, 副泥鳅属的进化关系并不稳定, 在COⅠ基因树中, 副泥鳅属位于树的根部; 在Cytb基因树中, 副泥鳅属嵌套分布于泥鳅属中。因此,本研究认同将副泥鳅属作为泥鳅属同属异名的观点。
3.2 黑龙江泥鳅与北方泥鳅的鉴定及物种有效性
本研究基于形态和分子的研究均支持北方泥鳅的有效性。虽然北方泥鳅被多数学者否认其有效性, 将其认为是黑龙江泥鳅的次定同物异名[6]。然而, 在形态特征上, 北方泥鳅与黑龙江泥鳅的形态差异最为直观, 如陈景星[2]、朱松泉[3]和李雅娟等[4]均指出北方泥鳅体细长、尾柄细长、背鳍前躯短、须短和尾柄皮质棱不发达等。本研究认为北方泥鳅属区别于黑龙江泥鳅还有以下特征: 上筛骨-中筛骨复合体前端延伸呈尖锐状; 梨骨呈蝶形;外枕骨腹后缘延长, 覆盖于枕髁上方; 基枕骨末端在背大动脉腹面分叉; 鳞焦小。易少奎[18]推测黑龙江泥鳅与北方泥鳅是泥鳅属物种进化过程中变化较为迅速的表型。在分子水平上, 北方泥鳅与黑龙江泥鳅的遗传距离介于18.1%—18.9%, 属于泥鳅属种级分化水平; 在系统进化上, 北方泥鳅与黑龙江泥鳅处于两个不同的分支, 且北方泥鳅与泥鳅具有较近的亲缘关系, 与黑龙江泥鳅亲缘关系较远。本文的分子结果与易少奎[18]的结论一致, 支持北方泥鳅为有效种。
基于Cytb的系统发育关系表明, 分布于日本的泥鳅与北方泥鳅聚为一支, 其遗传距离为7.0%, 属于种内分化水平, 分布于日本的泥鳅应为北方泥鳅。我国的黑龙江泥鳅与分布于俄罗斯部分俄罗斯泥鳅(JN858864、JN858861、JN858868、JN 858866、JN858867和JN858870)以高置信度(BI=1.00, ML=96%)聚为一支, 其遗传距离为11.9%,属于种间分化水平。Perdices等[6]曾认为俄罗斯泥鳅应为两个物种, 本研究支持其观点, Perdices所分析的材料包含2个物种: 俄罗斯泥鳅与黑龙江泥鳅。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:07:02
西北植物学报(2022年4期)2022-05-24 07:52:26
中国矫形外科杂志(2021年21期)2021-11-19 09:53:28
世界科学技术-中医药现代化(2021年5期)2021-11-05 06:56:18
科学大众·小诺贝尔(2021年9期)2021-01-22 12:41:56
新农民(2020年15期)2020-06-22 07:38:58
脊柱外科杂志(2020年3期)2020-02-11 21:42:01
现代园艺(2018年2期)2018-03-15 08:00:10
天津诗人(2017年3期)2017-11-14 17:26:10
创伤外科杂志(2016年12期)2016-03-24 02:18:10
