
长江靖江段沿岸日本鳗鲡丰度的时间格局及生物学研究
2021-04-01 07:56郭弘艺唐文乔倪健夫沈林宏
水生生物学报 2021年2期
郭弘艺 刘 丽 唐文乔 刘 东 张 亚 倪健夫 沈林宏
(1.上海海洋大学海洋动物系统分类与进化上海高校重点实验室, 上海 201306; 2.江苏省靖江市渔政管理站, 靖江 214500)
日本鳗鲡(Anguilla japonica)是一种降海产卵洄游型(Catadromous)鱼类, 其产卵场位于西马里亚纳海脊附近[1—4]。孵化后的柳叶鳗(Leptocephali)随北赤道流(North Equatorial Current)和黑潮暖流(Kuroshio Current)输送至中国、朝鲜和日本的大陆架, 变态为玻璃鳗(Glass eel), 并在河口水域变态为线鳗(Elver)[5]。线鳗溯河至河川或滞留在河口成长为黄鳗(Yellow eel)。数年后黄鳗开始性成熟, 逐步银化为银鳗(Silver eel), 返回至产卵场繁殖, 最终完成生活史[6]。这种复杂的生活史特性, 增加了人工繁殖的难度, 目前养殖所用鳗苗还全部依赖于天然捕捞[7]。由于价格高, 鳗苗捕捞一直处于高强度的“掠夺”模式[8], 加上近年来沿海湿地的减少及环境污染, 已造成资源的严重匮乏[9]。
江苏靖江位于长江近口段, 常年水流平顺, 受潮汐和长江径流的双重影响, 自然形成了流态复杂的水域环境, 不仅是多种鱼类栖息和繁殖的良好场所, 亦是长江鳗苗溯河和成鳗降海洄游的通道[10]。近年来, 我国学者对日本鳗鲡的遗传结构[11,12]、早期生活史特征[13—17]及其资源变动[9,10,18,19]等作了较多研究, 但由于非洄游时期鳗鲡样本的采集十分困难, 因此其在长江水系的资源状况一直未见报道。本文基于2002—2017年在长江靖江段长达十余年的采样调查数据, 对长江近口段日本鳗鲡丰度的时间格局及其年龄结构、全长与体重等的相关性作了初步分析, 以期为这一珍贵物种的保护及管理提供基础数据。
1 材料与方法
1.1 采样地点及方法
采样点位于江苏省靖江市新桥镇的长江沿岸江滩湿地(31°56"N, 120°01"E), 其沿岸上缘为混凝土堤坝, 下缘的堤脚为宽约30 m的大型石块带。石块带下缘至泥质江岸内侧有宽约100 m的密集芦苇(Phragmites australis)带, 夏秋季芦苇高约3 m, 冬天被革除。
垂直于江岸设置当地渔民所用典型“丁”字形定置张网一部, 拦网40 m长, 下网纲埋于江滩, 网片露出江滩高约1.8 m, 在拦网下缘近江心处布置2个笼式网袋。拦网网目为1.86 cm; 网袋网目为0.92 cm。张网位置随季节性的水位涨落而上下移动, 一般控制在平水期水面与拦网的上纲齐平。当网片破损或藻类附着过多时, 更换同一规格的张网。每天由专人在早潮退潮后收集一次渔获物。2002年1月至2003年12月, 每月保存1日、15日的2次样本;2004年1月至2017年12月, 每月均保存1日、11日和21日的3次样本。由于暴雨、洪水等原因, 造成部分样本缺失, 16年共获得519份渔获物。样本现场用10%福尔马林溶液固定后, 带回实验室分析。
1.2 实验方法
从每份渔获物中分离出日本鳗鲡作为研究对象, 用量鱼板逐尾测量全长L, 精确到0.1 cm; 干净纱布吸干鱼体水分后, 用电子天平称量体质量W,精确到0.01 g。采用幂函数拟合全长和体质量的关系, 表达式为:。式中:W表示鱼体质量(g);L表示鱼的全长(cm);a为生长的条件因子;b为幂指数。
观测并详细记录每尾样本头部、躯干部及胸鳍的体色, 依据郭弘艺等[20]对日本鳗鲡体色特征描述和评判标准, 鉴定每尾标本所处的发育期。挖取矢耳石作为年龄鉴定材料, 依据谢正丽等[21]的日本鳗鲡耳石制备、年轮确认和计数方法, 制备耳石磨片并鉴定样本的年龄, 耳石结构和年轮标志见图1。

图1 日本鳗鲡矢耳石横截面的结构和年轮Fig.1 Structure and annuli character on transversal section of the sagittal otolith of Anguilla japonica collected at the Jingjiang section of the Yangtze River
1.3 数据分析
采用Kruskal-Wallis检验对样本年龄、全长、体质量在时间尺度上的差异显著性进行检验。采用互相关函数(Cross-correlation function)对靖江段沿岸日本鳗鲡丰度与长江口鳗苗数量的时间序列进行互相关分析(Cross-correlation analysis), 长江口鳗苗为单船年捕捞量(尾/年), 时间分辨率为年, 数据来源于长江三峡工程生态与环境监测公报。数据处理和分析、图形绘制用SPSS Statistics 20和GraphPad Prism 8.0软件进行。
由于丰度较低, 0值较多, 本研究以日本鳗鲡的月出现率作为响应变量建立GAM模型, 分析各环境因子包括输沙量(万吨/月)、径流量(m3/s)、水温(℃)、潮差(m)、气象因子气压(hPa)、降水量(mm)、浑浊度(NTU)、化学需氧量CODMn(mg/L)和氨氮(mg/L)等的影响。GAM表达式为:

式中, 函数g()为联结函数, IO为日本鳗鲡的月出现率,为常数截距项。模型中的输沙量和径流量选取距离靖江段最近的长江干流大通水文站的数据, 数据来源于水利部长江水利委员会发布的长江泥沙公报。表层水温和潮汐数据(水位基准为吴淞高程)来源于靖江市水利局, 潮差为2次涨落潮潮差的平均值。气压、降水量来源于靖江市气象局;浑浊度、化学需氧量CODMn和氨氮来源于靖江市生态环保局。环境因子的时间尺度与日本鳗鲡出现率对应, 为2002年1月至2017年12月, 时间分辨率为月。
利用池信息量准则(AIC, Akaike Information Criterion)检验逐步加入因子后模型的拟合度,AIC值越小, 模型拟合度就越好[22,23]。根据F检验评估因子的显著性[22]。最终GAM模型采用样条平滑法,分布函数族为二项式分布, 模型构建和检验采用R 3.1.1软件中的mgcv包实现[22]。
2 结果
2.1 丰度和出现率的时间尺度变化
分析显示, 在519次渔获物中, 日本鳗鲡出现了113次, 总体出现率为21.8%。共采集到日本鳗鲡137尾、8181.61 g, 仅占渔获物数量和重量的0.08%和0.69%。
年际分析显示, 在长江靖江段沿岸, 每年均采集到日本鳗鲡个体, 年均丰度为(9±4.2)尾。其中,2010年之前的年均出现率为28.2%, 2002年曾高达50.0%; 但此后7年的出现率大幅降低, 年平均仅为15.1%, 总体呈波动式递减的趋势(图2)。
月际比较显示, 日本鳗鲡在5—11月的出现率较高, 12月至次年3月的出现率较低(图3)。此外,2010年以前每年平均有6.0月可采集到样本, 2010年以后仅4.1月可采集到样本, 同时出现率呈明显的季节性后移, 1—4月未曾采集到样本。
2.2 生物学特征

图2 2002—2017年长江靖江段沿岸日本鳗鲡丰度和出现率的年际变化Fig.2 The abundance and incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
采集到的137尾个体全长在17.0—65.0 cm, 平均为(31.1±9.8) cm; 体质量在5.03—499.50 g, 平均(57.64±91.94) g; 丰满度在0.80—2.38, 平均为1.23±0.30(n=137)。据全长和体质量数据,拟合所得关系式为:W=0.0002415×L3.483(r2=0.9537,n=137,图4)。经耳石磨片鉴定年龄的126尾个体, 年龄在1—6龄, 平均(1.8±1.0)龄。
时间尺度分析显示, 个体全长(P=0.387>0.05)和体质量(P=0.560>0.05)在年际间均无显著差异,但在月间存在显著差异(P<0.05)。由图5可见, 全长大于45 cm、体质量大于200 g的样本仅出现在9—11月的成鳗降海洄游季节。年龄的月分布图亦显示, 1龄和2龄个体分别占总样本数的48.4%和40.5%, 3龄及以上样本仅占11.1%, 且仅出现在9—12月(图6)。
依据性腺形态, 仅有15尾个体可鉴定性别(8尾雄性、7尾雌性), 其他89.1%的样本均为未成熟个体。依据体表的色素发育程度判别, 样本中有黄鳗期130尾、银化前期(Pre-silver stage)7尾, 分别占个体总数的94.9%和5.1%, 且银化前期仅出现在10—11月。
2.3 丰度与环境因子的关系
广义可加模型(GAM)分析显示, 径流量、泥沙量、潮差、气压、降水量和化学需氧量CODMn对日本鳗鲡的出现率影响均不显著(P>0.05), 而水温及氨氮和浑浊度对其出现率却有显著影响(P<0.05)。从偏差变化及AIC值可以看出, 模型中环境因子对日本鳗鲡出现率的总偏差解释率为34.0%。其中以水温影响最大, 偏差解释率为25.8%, 水质指标氨氮和浑浊度偏差解释率分别仅为4.0%和4.2%(表1)。
由图7可见, 在10—25℃水温区间内, 日本鳗鲡的出现率呈增加趋势, 水温至25℃左右, 出现率达到最大。而氨氮和浑浊度与出现率均呈负相关关系,即随着水体中氨氮浓度和浑浊度的增大, 日本鳗鲡的出现率呈下降趋势。当水体中氨氮高于0.35 mg/L、浑浊度大于130 NTU时, 日本鳗鲡几乎就不出现。
3 讨论
3.1 长江沿岸日本鳗鲡群体的组成
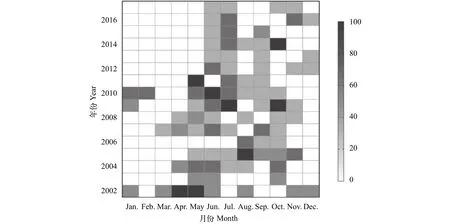
图3 2002—2017年长江靖江段沿岸日本鳗鲡出现率的月际变化Fig.3 Monthly incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
长江靖江段处于长江下游与河口段的交汇地带, 受长江淡水径流及海洋咸水潮汐的交互作用,形成了独特的生态环境, 为多种鱼类提供了育幼和索饵的良好场所[24]。其中的沿岸水域由于具有独特的自然资源特征和生态系统过程, 具有很高的生态服务价值, 在维持河口鱼类多样性和渔业资源上具有独特的作用[25]。本文采集到的137尾日本鳗鲡平均全长为(31.1±9.8) cm, 体质量为(57.64±91.94) g,平均年龄仅(1.8±1.0) 龄。由此可见, 线鳗溯河成功、已在淡水育肥1—2龄的黄鳗是日本鳗鲡长江近口段沿岸群体的主要成分, 也预示着长江沿岸是日本鳗鲡幼鱼良好的栖息和索饵场所之一。已有研究表明, 在日本鳗鲡生殖洄游过程中消化道会逐步萎缩, 也不再摄食, 洄游所需的能量消耗来源于洄游前的积累[26,27]。但本研究显示, 10—11月间有部分已启动生殖洄游的银化前期个体出现本研究水域, 表明日本鳗鲡在降海生殖洄游的初期也需要沿岸生境的依托。
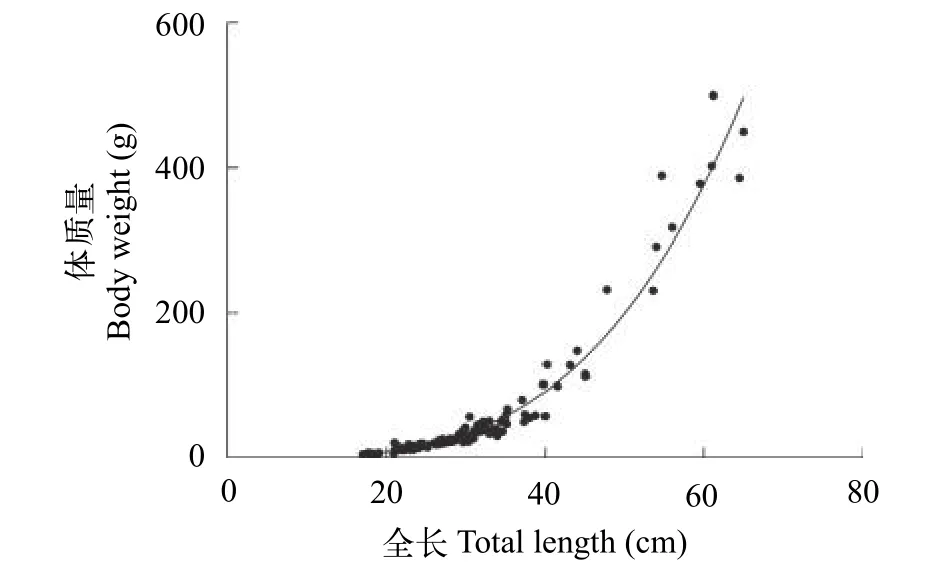
图4 2002—2017年长江靖江段沿岸日本鳗鲡全长体质量关系Fig.4 The relationship between body weight and total length of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017

图5 2002—2017年长江靖江段沿岸日本鳗鲡全长和体质量月际小提琴变化图Fig.5 The monthly violin plot of total length (a) and body weight(b) of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017

图6 2002—2017年长江靖江段沿岸日本鳗鲡年龄月际小提琴变化图Fig.6 The month violin plot of age of Anguilla japonica collected at the Jingjiang section of the Yangtze River during 2002—2017
全长-体质量关系式中的特征参数b反映鱼类在不同生活史阶段、季节和环境中的生长情况[28]。本研究发现长江沿岸生境鳗鲡群体的b值达3.483,高于福建萩芦河的黄鳗群体(b=3.268)[29], 也高于同一水域敞水生境的银鳗群体(b=3.246)[21]及珠海口(b=3.051)[30]和日本滨名湖(b=3.400)的银鳗群体[31](表2)。可见, 长江更适合于日本鳗鲡的生长和肥育。
3.2 长江日本鳗鲡资源动态
大量研究显示, 由于围湖造田、江湖阻隔、水环境污染及过度捕捞等人类活动, 造成了世界各地的日本鳗鲡自然种群均处于枯竭状态[9,19,32]。本研究连续16年的监测结果显示, 日本鳗鲡在靖江段沿岸鱼类渔获物的出现率为21.8%, 仅占渔获物数量与重量的0.08%和0.69%, 资源相当匮乏。这一结果与帅方敏等[9]对珠江水系的调查结果相似。虽然长江三峡工程生态与环境监测公报显示, 2002—2017年长江口鳗苗单船捕捞量平均高达19161尾/年(图8),资源量并不贫乏[32], 但由于日本鳗鲡的生活史很复杂, 真正能溯河至长江并且能发育至黄体鳗的数量极有限[10]。互相关分析亦证实, 靖江段沿岸当年日本鳗鲡丰度与前1—2年进入长江口的鳗苗数量并没有显著的相关性(图9)。
近年来, 随着长江禁渔期的实施和水产种质资源保护区的建立, 长江靖江段沿岸的刀鲚、胭脂鱼和乌鳢等的幼鱼资源都有增长的趋势[34]。但本研究分析却显示, 日本鳗鲡的丰度和出现率均呈逐年波动式下降的趋势。有研究指出, 大坝修建的阻隔、河流宽度与水深是造成鳗鲡空间分布结构差异的最主要因子[9]。但本文的GAM分析结果显示,水温、氨氮和浑浊度是造成日本鳗鲡的出现率差异的主要影响因子。由于靖江沿岸水域近年岸线围垦、堤岸硬化和码头建设的加剧, 河岸植被大多消失, 加之污染物的超标排放, 渔业资源维持的生态服务功能也正受到威胁[35]。值得注意的是, 环境因子对日本鳗鲡出现率的解释率仅为34%, 其他影响其资源波动的因子仍有很多, 有待于更深入研究。此外, 本研究仅分析了靖江段沿岸的一个调查点, 难以有效评估整个长江水系的资源状况和时间分布特征。

表1 长江靖江段日本鳗鲡出现率与环境因子的GAM检验Tab.1 Significance of explanatory variables effects on incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River as determined by generalized additive model

图7 广义可加模型(GAM)所揭示的显著影响长江靖江段日本鳗鲡出现率的环境因子Fig.7 Generalized additive model (GAM)-derived effects on incidence of occurrence of Anguilla japonica collected at the Jingjiang section of the Yangtze River

表2 不同水域的日本鳗鲡生长参数比较Tab.2 Comparison of growth parameters of Japanese eels in different waters

图8 2002—2017年长江口日本鳗鲡鳗苗单船年捕捞量Fig.8 Annual catch of Anguilla japonica glass eels collected on the Yangtze River estuary during 2002—2017

图9 长江靖江段沿岸日本鳗鲡丰度与长江口日本鳗鲡鳗苗单船年捕捞量间的互相关分析Fig.9 Cross-correlation analysis of Anguilla japonica abundance collected at the Jingjiang section of the Yangtze River and annual catch of Anguilla japonica glass eels collected on the Yangtze River estuary during 2002—2017
猜你喜欢
化工管理(2022年14期)2022-12-02
江苏安全生产(2022年3期)2022-04-19
中老年保健(2021年11期)2021-08-22
华人时刊(2021年21期)2021-03-09
青年歌声(2020年11期)2020-11-24
哈尔滨工业大学学报(2020年6期)2020-06-11
收藏界(2019年4期)2019-10-14
天津科技(2019年9期)2019-09-28
天津诗人(2017年2期)2017-11-29
天津诗人(2017年2期)2017-11-29
