
水稻稻瘟病抗性基因在抗性育种中的研究进展
2021-04-01 09:02:22韩雪琴沈文娟张振海罗成科杨淑琴李培富张银霞
新疆农业科学 2021年3期
韩雪琴,沈文娟,张振海,田 雷,罗成科,杨淑琴,李培富,张银霞
(1.宁夏大学农学院,银川 750021,2.宁夏回族自治区原种场,银川 750200,3.宁夏农林科学院农作物研究所,银川 750105)
0 引 言
【研究意义】水稻(oryzasativa)是全世界第二大粮食作物[1]。病害是影响水稻生产最重要的限制因素,其中稻瘟病作为水稻的三大病害之一,影响着水稻的高产、稳产和优质。稻瘟病是由真菌(magnaportheoryzae)引起的一种极具破坏力的真菌性病害,多样化程度高,具有流行性、突发性、毁灭性的特点。在适宜温度和高湿条件下,水稻产区造成的产量损失在30%~50%[2],在发病严重的时候,造成水稻颗粒无收。常规育种只是通过植株的表型和调查植株的抗性鉴定相结合的方法选育抗病品种,这样的选育不能使多基因聚合,并且选育的品种不具有广谱和持久的抗性,不能有效控制病害的发生。采用化学防治方法来控制稻瘟病,虽然可以发挥一定的作用,但对环境的污染逐渐加重,并且严重影响着水稻的优质特性,不能从根本上控制病害的发生。生产中采用种植含有广谱持久主要抗病基因的抗病品种是控制病害的发生最为经济有效且环保的方法,而广谱持久抗性基因的挖掘和鉴定是前提条件。研究水稻稻瘟病抗性的研究进展,对水稻抗性育种有重要意义。【前人研究进展】已有大约100个抗性基因被鉴定出和500个抗病基因相关的数量性状位点[3],在水稻基因组中被鉴定和定位的基因中,已有37个基因被成功克隆[4]。【本研究切入点】研究以水稻稻瘟病抗性的机理,抗性基因的定位、克隆,以及抗性基因在抗性育种中的应用研究近况进行综述。【拟解决的关键问题】汇总和对比分析国内外的相关文献,分析不同抗性基因在水稻抗稻瘟病中的应用研究进展,综述水稻抗稻瘟病基因的研究现状及前人利用抗稻瘟病基因进行抗性育种的最新研究结果,分析已克隆抗稻瘟病基因在水稻抗性育种应用中存在的问题及前景,为现阶段的抗稻瘟病育种研究提供参考。
1 材料与方法
1.1 材 料
查阅国内外相关文献以及国家水稻数据中心,收集与水稻稻瘟病抗性基因有关的研究报道。
1.2 方 法
整理汇总,从水稻稻瘟病抗性的机理,抗性基因的定位、克隆,以及抗性基因在抗性育种中的应用的研究近况进行综述。
2 结果与分析
2.1 水稻稻瘟病抗性机理的研究与应用
2.1.1 基因对基因假说
基因对基因假说(gene for gene hypothesis)最早是由Flor在研究亚麻与锈病之间的互作时提出的假说[5]。后来Jia等[6]在水稻抗病基因与稻瘟病菌无毒基因互作中也发现了该假说,揭示了水稻和稻瘟病菌之间特性识别和互作关系。该假说认为,寄主本身具有抗病基因(R)和感病基因(S),病原菌本身具有无毒基因(AVR)和有毒基因(VIR)。当携带有毒基因的病原菌侵入含有感病基因或不含有抗病基因的寄主时,寄主会感病,这种感病反应称为亲和反应。相反,当携带无毒基因的病原菌侵入含有抗病基因的寄主时,寄主会抗病,这种抗病反应称为不亲和反应。这种亲和和不亲和性主要取决于寄主与病原菌之间有无亲和互作关系。例如,Pi-ta与AVR-Pita互作是第一对被鉴定的水稻R蛋白与稻瘟病菌无毒蛋白AVR间的互作对,Pi-ta编码产物的蛋白类型为NBS-LRR,其LRR结构域能与无毒蛋白AVR-Pita在酵母细胞中直接互作[7]。
2.1.2 防卫假说
防卫假说是由Vander和Jones对基因和基因假说的继承和发展[8]。在基因和基因假说中认为,R基因编码的R蛋白和病原菌无毒基因编码的Avr蛋白之间发生相互作用。但在该假说中认为,寄主体内存在一个可以让病原菌和寄主互相识别的蛋白,这种蛋白会先与病原菌的毒性Avr蛋白结合,然后吸引寄主内相对应的R蛋白,R蛋白与Avr蛋白相互识别并形成一种能识别信号的特殊结构,最后诱导防卫基因的表达。在拟南芥的RIN4蛋白参与的丁香假单胞菌抗性反应中就对这种假说有直接的证据,RIN4蛋白会分别与NBS-LRR抗性蛋白RPM1和RPS2形成独特的复合体,启动一系列信号的反应,参与其免疫反应[9]。在正常情况下,1对Pik-h抗病蛋白是处在非激活状态下的,但当病原菌侵入后,寄主植物的防御系统就会被打破,这时的识别效应子Avr-Pik-h的抗性蛋白Pikh-1就会通过改变构象激活抗病蛋白Pikh-2,由Pikh-2传递相关的信号来激发防御反应[10]。
2.1.3 水稻稻瘟病抗性基因
水稻的抗性基因有2类,分别为主效抗性基因和微效抗性基因[11]。抗稻瘟病基因的抗性分为2种,1种是垂直抗性和水平抗性[12],垂直抗性也叫完全抗性,一般由1~2个主效抗性基因控制,但少数的主效基因是由隐性基因或不完全显性基因控制,属于质量性状;带有这类抗性基因的水稻和病原菌之间有不亲和的互作方式,菌株的专化性强,寄主会随着菌株的变异而丧失抗性。例如,地谷的主效抗稻瘟病基因Pid3对我国稻瘟病生理小种Zhong-10-8-14表现出较强的抗性[13]。另一种是水平抗性也叫部分抗性,一般由多个微效抗病基因共同控制水稻对稻瘟病的抗病性,属于数量性状;带有这类抗性基因的水稻和病原菌之间有亲和的互作方式,对不同的稻瘟病生理小种都表现出抗性,使菌株的专化性变弱,使水稻具有持久的抗性。例如,Pb1是在籼稻品种mudan中克隆得到,但该基因只在水稻成熟阶段表现出较有效的抗性,尤其是对水稻的穗颈瘟[14]。
2.2 水稻稻瘟病抗性基因的定位
Sasaki通过人工接种的方法,确定了日本水稻品种中有1个显性抗病基因[15]。1965年,Goto Yamasaki对稻瘟病进行了系统性的分类研究。1966年,Kiyosawa等定位到了第一个抗性基因pia[16];到1981年,他再次定位到了8个位点上的14个稻瘟病主效显性基因,分别为pik位点上的pikh、pikm、pikp、piks;pita位点上的pita、pita2;pikz位点上的pikz、pizt以及pia、pish、pib等[17]。1991年,Yu等在12号染色体上定位到了pi-6(t) 基因,还有pi-1(t)、pi-2(t)、pi-4(t) 等基因[18]。1994年,Mew等在Yu的基础上对这3个基因进行了更加精密的定位[19]。同年,Wang等[20]定位到了2个抗性基因,分别为pi-5(t)和pi-7(t) 。而中国对稻瘟病抗性的研究较晚,在1976~1979年才完成了全国稻瘟病生理小种的鉴定,为稻瘟病的研究奠定了基础[21]。1994年,朱立煌等[22]在8号染色体上定位到了pi-zh,这也是第1次在8号染色体上发现抗病基因。之后又在8号染色体上定位到了pi33、pi-11、pi-GD-1基因,在11号、6号、12号染色体上定位到了pi-1、pi-2、pi-4等基因,其中pi-1是第一个定位在11号染色体上的基因。2001年,朱献丰等[23]在12号染色体上定位到了pi6基因。2002年,吴金红等[24]在Mew的基础上对pi-z(t) 进行了精确定位。2003年,Berryer等定位到了位于8号染色体上的pi-33(t) 基因[25]。近年来,随着分子生物学的快速发展,已有大约100个抗性基因被鉴定出和500个抗病基因相关的数量性状位点[3]。表1(http://archive.gramene.org)
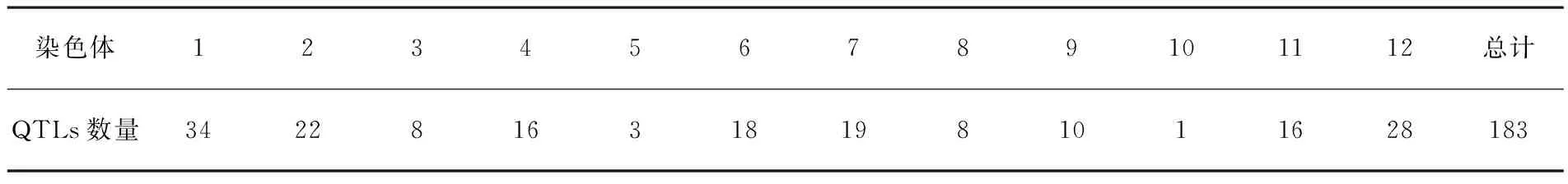
表1 已鉴定的稻瘟病抗性基因Table 1 Rice blast resistance genes identified
2.3 水稻稻瘟病抗性基因的克隆
通过使用图位克隆法,在水稻基因组中被鉴定和定位的基因中,已有37个基因被成功克隆[4]。其中Pita和pib是最早被克隆出来的基因[26,27],而pib是第一个克隆出来的主效抗瘟基因,它位于2号染色体上[27]。Pi9是第一个广谱抗瘟基因被克隆出来[28]。Pi21是第一个被克隆出来的部分抗性基因[29]。Bsr-d1是近期克隆的部分抗性基因,是从广谱持久抗病的材料地谷中发现的[30]。Zhao等[31]鉴定出了一个结构新颖的广谱抗病基因ptr,ptr的广谱抗性不依赖pi-ta,但pi-ta的广谱抗性依赖ptr。这些基因不均匀的分布在各条染色体上,其中11号和6号占多数,分别占35.13%和24.32%。而5号、7号、10号染色体上暂时没有克隆出有关基因。已克隆的稻瘟病抗性基因中有33个属于NBS-LRR蛋白类型,其中,pi-b属于典型的NBS-LRR类抗性基因,编码产物为细胞质蛋白;Bsrd-1基因编码的蛋白类型为MYB转录因子;Pi-21基因编码的蛋白类型为富含脯氨酸蛋白;Pid-2基因编码的蛋白类型为类受体激酶蛋白;ptr基因编码的蛋白类型为膜蛋白。其中19个基因具有叶瘟抗性,pi-25基因对叶瘟和穗颈瘟都具有抗性;pi-63和pid-3对整个时期的稻瘟病都具有抗性,占已克隆的稻瘟病抗性基因的5.4%;pb1在水稻成熟阶段具有较有效的抗性,对穗颈瘟具有很好的抗性。表2
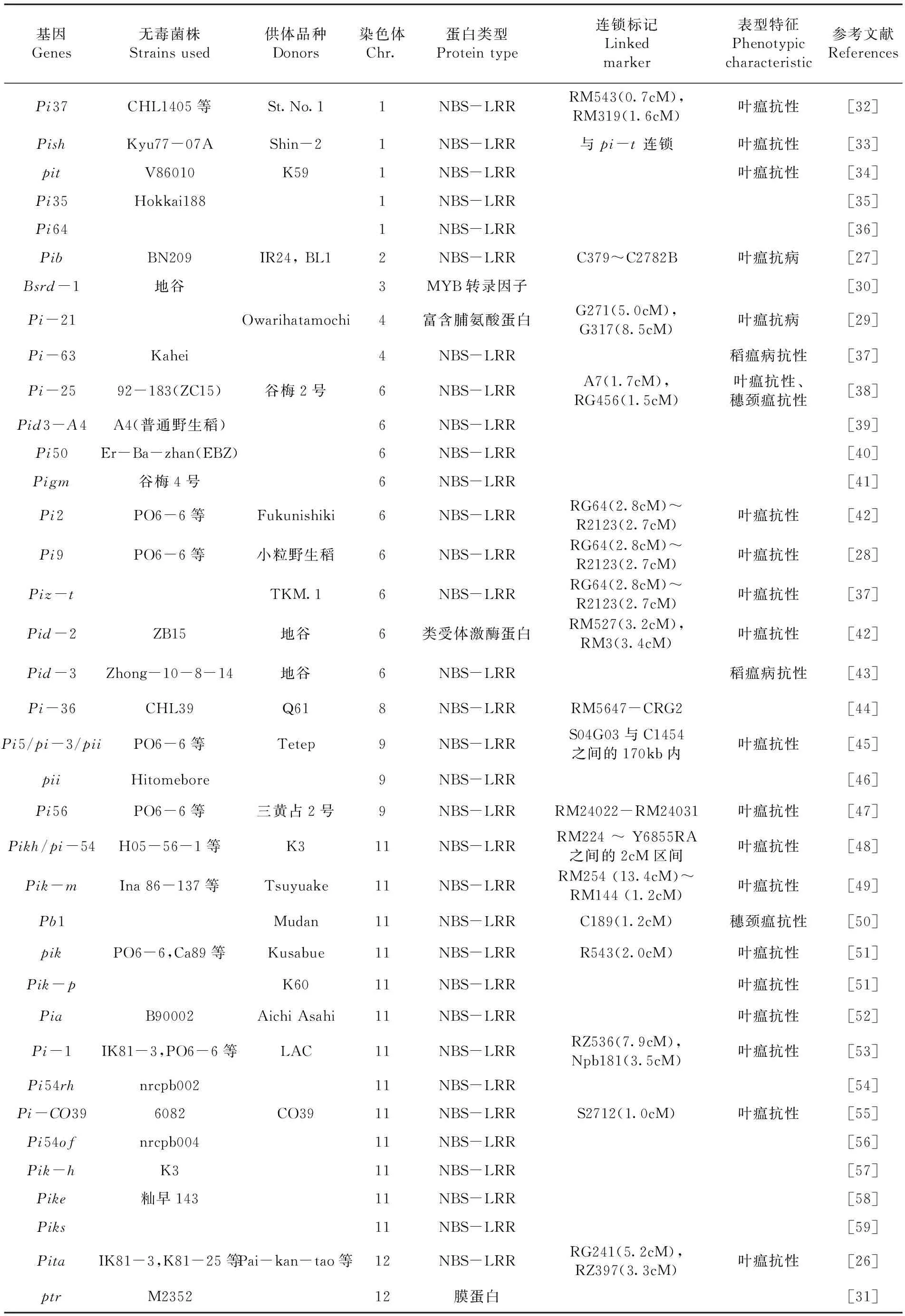
表2 已克隆的稻瘟病抗性基因Table 2 Cloned blast resistance genes in rice
2.4 水稻稻瘟病抗性基因在育种中的应用
2.4.1 常规育种
常规育种的方法是通过有性杂交,利用田间农艺性状选择和抗性鉴定相结合,将抗病基因导入目标株系育成抗病品种。常规育种从选择亲本开始,利用单交、复交、回交的手段,将抗稻瘟病基因重组到一个新的品种中去。例如,可以选用品质优、产量高但不抗病的品种和抗病但不具有此优点的品种作为亲本,来选育新的抗病优质高产品种。在抗病育种中,导入的抗病基因的遗传传递能力要强、抗谱性要广,导入的抗源材料亲和力要强,这样才能容易有效的把抗病基因导入新的品种中。目前,常规育种仍然是水稻抗稻瘟病育种中比较有效的方法。但此方法是通过田间抗性鉴定为依据,田间不同的抗性基因会出现交叉相掩盖现象,只能选择1个或2个抗性基因,不利于多基因聚合的选择,很难实现选育的新品种具有广谱抗性和持久抗性。
2.4.2 分子标记辅助选择育种
分子标记辅助选择(molecular marker-assisted selection, MAS)是将分子标记应用于作物育种中的一种选择辅助手段。该方法是利用与目的基因紧密连锁或基因本身的分子标记来选择基因型,在分子水平上确定材料的基因型,同时还可以对全基因组进行筛选,来减少连锁累赘。此方法不受环境和稻瘟病病菌生理小种的影响,可以有效的提高育种的效率和准确性,还可以在短时间内选育出具有持久抗性的水稻品种。近年来,研究者利用此方法开展了稻瘟病抗性育种,并且取得了实质性的进展。
分子标记辅助育种的前提是利用分子标记对稻瘟病基因型进行检测,对品种的抗性进行针对性的改良。最初是通过回交的方法,将供体中的目的基因渗入到受体中,但随着目的基因的渗入,与目的基因连锁的其它基因也会渗入到受体中,形成“连锁累赘”。分子标记辅助选择必须应用与抗稻瘟病基因紧密连锁的标记,可以在目的基因附近发生重组个体,比较理想的是标记离目的基因5cM,这样选择到的目的基因植株的正确率在99.75%以上[60]。李仕贵等[61]应用与稻瘟病抗性基因pi-d(t) 紧密连锁的微卫星标记RM262对含有该抗病基因的品种地谷与感病品种江南香糯8987的F2群体进行MAS选择,结果表明,应用该标记的抗性重合和杂合带型选择抗性植株的准确率在98%以上。文绍山等[62]利用与抗稻瘟病基因pi-9(t) 紧密连锁的分子标记pB8对育种后代进行检测,筛选到携带有pi-9(t) 基因的恢复系新株系。
分子标记辅助选择在基因聚合育种方面也有着广泛的应用,在基因聚合育种中,应用目标性状紧密连锁的分子标记进行辅助选择,即将多个不同抗性基因聚合到1个新的品种中,这样可以打破回交育种中只能改良个别性状的局限性,可以使品种的多个性状得到改良,提高育种的效率,缩短育种周期,得到更有价值的育种材料,并且可以有效的克服病原菌的变异。Hittalmani等利用MAS选择将分别含有抗稻瘟病基因pi-l、pi-5、pi-ta聚合到同一品系BL24中,得到改良单株,其抗性比含有单个抗病基因的单株要强[63]。董巍等[64]通过MAS选择的方法将稻瘟病抗病基因pi-1、pi-2回交聚合到培矮64S中,筛选到10株对稻瘟病抗病明显增强的改良株系。
2.4.3 基因工程育种
基因工程育种是水稻稻瘟病抗性育种中最直接的方法之一,它是以抗性基因克隆为基础,将克隆后的抗性基因经过剪切和拼接加工后,借助载体导入受体细胞中,使抗性基因融合与受体细胞中,受体细胞表达抗性,受体材料就会对稻瘟病具有抗性[65]。基因工程育种也可以将水稻基因库中不具有的各种抗性相关基因转入水稻中,拓宽抗病基因的来源,培育出新的抗病材料,为抗病育种提供新途径[66]。潘素君等[67]通过农杆菌介导法将广谱抗稻瘟病基因Pi-9转入5个籼稻品种,接种鉴定表明,转基因植株具有很强的稻瘟病抗性。以抗稻瘟病的粳稻中花9号转溶菌酶基因材料 D2-1-2 与籼型杂交稻两优培9的恢复系 9311 及不育系培矮64S、汕优 63 的恢复系明恢 63 分别杂交和多代回交,进行外源溶菌酶基因的转育。结果表明,转育后代抗性与轮回亲本相比、测交组合抗性与对应杂交稻组合相比都有了较大提高。随着转育回交世代的增加,抗性增强得越明显[68]。但此方法对实验室的要求和对操作者的技术要求比较高,成本也较高,在实际生产中较难普及。
3 讨 论
3.1 目前水稻稻瘟病抗性育种的研究大多数还停留在传统的育种研究。这种育种方法不仅耗时长还费人费力。传统育种出的新品种存在单一性,但病原菌变化多样,环境条件变化多端,新品种不能有长期的抗性效果,要不断的培育新的抗病新品种;在常规杂交育种中,对于多个抗性基因聚合时,由于不同的抗性基因间存在抗谱的交叉重叠,在后代中出现大量的假阳性现象,降低了选择的目标性和准确性。分子标记辅助选择育种、基因工程育种可以有效的提高抗性基因的准确性,但这些方法成本高、不利于规模化检测,应该加强与抗性基因紧密连锁或分离的操作简便、成本低、方便规模化检测的分子标记。
3.2 有些抗病品种在当地表现为抗病,但在其它地方抗病性却表现较差,这是由于可能某些抗病基因受到环境条件和病原菌变化的影响,导致其抗病性不能顺利表达有关。这就需要育种工作者对稻瘟病病原菌与水稻寄主之间的相互关系进行深入的研究,以便解决水稻的广谱持久抗性问题。
3.3 在水稻抗稻瘟病育种中,一般携带抗病基因的品种,其农艺性状表现较差,会出现分蘖少、产量低、晚熟等症状,这是由于受基因连锁等因素的影响,所以打破这种不良连锁将推动水稻抗病害育种产生质的突破。
3.4 虽然已经有很多抗稻瘟病基因被定位、克隆,并且在育种中表现出来了一定的抗性,但对大多数基因的抗性、广谱性和持久性以及生态适应区还不清楚,有些抗病基因还未标记到。因此,还需要对抗性资源进行系统的研究,继而通过分子标记遗传改良水稻的稻瘟病抗性,加速水稻抗病分子育种进程。
抗稻瘟病基因在水稻育种应用上,抗性基因可操作性强、效果明显,但这些基因在生产中的分布及利用情况尚不清楚。比如,Piz和Pik是水稻中目前报道含有广谱高抗稻瘟病最多的2个位点,它们分别包含pi2、pi9、pizt和pi1、pikh、pikm等复位基因[69-71]。另外,pita和pib对我国很多稻区也表现出很高水平的抗性[72-73]。但这些位点或基因的分布及利用上还缺乏完善的研究,限制了它们在育种中的应用。
4 结 论
已有大约100个抗性基因被鉴定出和500个抗病基因相关的数量性状位点;在水稻基因组中被鉴定和定位的基因中,已有37个基因被成功克隆;在不同的育种方法中,基因工程育种是水稻稻瘟病抗性育种中最直接的方法。水稻稻瘟病抗性基因的鉴定是抗性育种的基础,抗性基因的充分利用有利于快速有效的选育出新的具有广谱和持久抗性的抗病品种。
猜你喜欢
今日农业(2022年4期)2022-06-01 06:12:46
作物学报(2022年6期)2022-04-08 01:26:44
空间科学学报(2021年1期)2021-05-22 01:36:34
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
浙江农业学报(2017年1期)2017-05-17 06:13:50
河南农业(2016年6期)2016-11-26 07:06:46
新农业(2016年20期)2016-08-16 11:56:22
中国果菜(2016年9期)2016-03-01 01:28:44
现代农业(2016年5期)2016-02-28 18:42:37
中国蔬菜(2015年9期)2015-12-21 13:04:40
