
黄海北部真核微藻粒级结构及环境关联
2021-03-30 06:18毕相东宋广军吴金浩刘苏萱辽宁省海洋水产科学研究院辽宁省海洋生物资源与生态学重点实验室辽宁大连60天津农学院水产学院天津市水产生态及养殖重点实验室天津0084大连海洋大学辽宁大连60
中国环境科学 2021年3期
宋 伦,毕相东,付 杰,宋广军,吴金浩,刘 印,刘苏萱(.辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室,辽宁 大连 60;.天津农学院水产学院,天津市水产生态及养殖重点实验室,天津 0084;.大连海洋大学,辽宁 大连 60)
黄海北部是海水贝类主要增养殖区,但近十多年扇贝、牡蛎、杂色蛤等滤食性贝类普遍出现滞长、肉质偏瘦、死亡率升高等现象,限制了海洋渔业可持续健康发展.作为滤食性贝类的饵料基础—海洋微藻,在生理功能、沉降速率和被摄食压力等方面存有差异,致使微藻粒级对生态系统的食物网结构影响较大[1].由于滤食性贝类对饵料微藻具有选择性策略[2],因此微藻粒级结构会影响其营养储备和健康生长,小粒级微藻对质量选择型的贝类影响更大[3].然而微型(2~20μm)和微微型(0.22~2μm)藻类个体微小、形态学鉴定困难,粒级结构研究较为迟缓.
褐潮的暴发引起了各界对小粒径微藻研究的高度关注[4-5],高通量测序分子鉴定技术的发展极大地推动了小粒径微藻多样性的高效检测,进而加速了其对生态系统的影响研究[6-7].然而,有关微藻粒级结构的测算方法亟待优化.
目前微藻粒级测算技术主要有叶绿素 a分级法、电子粒度分析仪法、流式细胞术法、显微测量粒径分析法、分子鉴定法等,其中前三种可对微藻生物量进行粗略区分,无法获取各组分种类信息,显微测量粒径分析法无法观测微微型和部分微型藻类,然而该组分在整个微藻群落中占比较大.研究发现,叶绿素a分级法、电子粒度分析仪法都会低估中、小微藻的生物量,而高估>20μm 小型微藻的贡献[8].目前常用的叶绿素a分级法由于大孔径滤膜截留率较高严重低估了小粒径微藻的贡献,误差最高超过80%[9-10].分子鉴定虽然还存在一些问题,但比传统形态学鉴定方法更为精准、客观、高效,可大大提高微微型藻类的检出率,随着高通量测序技术的发展极大地推动了微型藻类的高效检测研究,为微藻粒级研究提供了新思路.
由于不同真核生物中rDNA的拷贝数差异较大,因此rDNA扩增子测序所获的序列数并不能表征环境样本中真核微藻的丰度[11].但相关研究发现,rDNA序列多态性与细胞中 rDNA的拷贝数成正比[12],而真核微藻细胞体积(生物量)与rDNA的拷贝数成正比[13-15],因此,真核微藻分子鉴定获取的序列数可表征其生物量大小,用于微藻全粒级结构研究可行.宋伦[16]采用高通量测序-分子鉴定分级技术研究发现,2018年春季大连长海县大长山岛邻近海域小粒径微藻(0.22~3μm)粒级组成主要受总氮影响较大,夏季受总氮、总磷影响显著,秋季受溶解硅影响显著,大粒径微藻(>20μm)粒级组成在夏季主要受溶解无机磷影响较大,中粒径微藻(3~20μm)粒级组成受上述环境因子影响不显著.
然而整个黄海北部海域微藻粒级结构在时空上如何演变,哪些环境因子对各粒级微藻影响显著,需要进一步研究和分析.基于此,本文采用高通量测序-分子鉴定分级技术对黄海北部微藻粒级结构进行研究,以期解析滤食性贝类的供饵力基础及环境关联.
1 材料与方法
1.1 样品采集及制备
2019年在黄海北部海域垂岸布设12个站位(图1),分别于 2019 年 5 月(春)、8 月(夏)、10 月(秋)采集真核微藻分子鉴定样品.
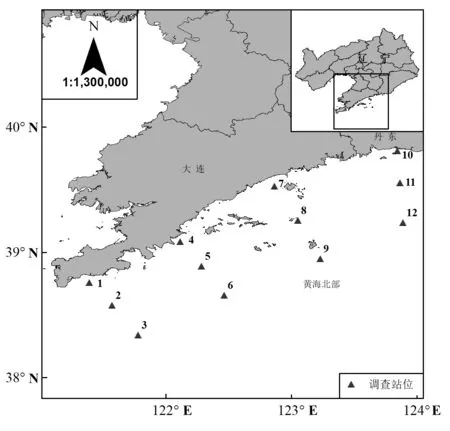
图1 采样站位示意Fig.1 Sampling sites
每个站位采集表层海水1L,现场用0.22μm微孔滤膜收集全部微藻,然后将滤膜转移至 1.5mL无菌离心管中,置于-20℃冷冻保存、运输.同期进行海水的水深(Dep)、pH、盐度(Sal)、溶解氧(DO)、悬浮物(SS)、叶绿素 a(Chla)、化学需氧量(COD)、石油类(Oil)、无机氮(DIN)[铵盐(NH4+)、硝酸盐(NO3-)、亚硝酸盐(NO2-)之和]、无机磷(DIP)、总氮(TN)、总磷(TP)、硅酸盐(SiO32-)、重金属汞(Hg)、铅(Pb)、铜(Cu)、锌(Zn)、镉(Cd)、铬(Cr)、砷(As)等指标的监测.调查、检测、质量控制方法均参照《海洋调查规范》(GB/T 12763-2007)[17]和《海洋监测规范》(GB17378-2007)[18]执行.
1.2 微藻基因组DNA提取
采用 CTAB法提取真核微藻宏基因组,将滤膜剪碎置于1.5mL离心管中,加入500μL CTAB裂解液(2% CTAB;100mmol/L Tris-Cl,pH为8.0;1.4mmol/L NaCl;10mmol/L EDTA)和 1μL β-巯基乙醇,5~10μL蛋白酶K,55℃裂解1~1.5h;短暂离心,取出液体置于新离心管中,用酚氯仿抽提2次后,取上清液,加入两倍体积预冷的无水乙醇,沉淀 2~3h,保留沉淀,使用75%乙醇清洗沉淀,得到浮游生物基因组 DNA,利用1%琼脂糖凝胶电泳和紫外分光光度计检测DNA浓度及纯度,合格后置于-20℃冰箱保存备用.
1.3 18S rDNA V4可变区的PCR扩增
该研究应用的引物为自行开发的真核微藻18S rDNA V4区基因扩增引物——V4(F/R).上游引物为V4-F序列 5'-GCGGTAATTCCAGCTCCAATA-3',下游引物为V4-R序列5'-GATCCCCHWACTTTCG TTCTTGA-3'[19].将引物连接适当的接头送往上海生工生物公司进行合成.PCR反应体系为 50 μL,包括 PCR Buffer 5μL、dNTP Mixture 8μL、上下游引物(10μmol/L)各 2μL、模板 DNA 2μL、Taq DNA 聚合酶2.5U,加适量灭菌水.扩增反应均在PE 9700型PCR仪(美国PE公司)上完成,反应条件:94℃预变性3min;94℃变性 30s,58℃退火 45s,72℃延伸 45s,共33个循环;72℃延伸5min.1%琼脂糖凝胶电泳检测PCR产物,将检测合格的产物交由诺和致源生物信息科技有限公司,使用 NEB Next® Ultra™ DNA Library Prep Kit for Illumina(New England Biolabs)建库试剂盒进行文库的构建,构建好的文库经过Qubit定量(Thermo Scientific)和文库检测,合格后,使用Hiseq2500PE250模式进行上机测序.
1.5 测序数据质量控制
通过IlluminaHiSeq 2500测序平台进行PE250模式测序,所得原始数据使用 FLASH软件进行拼接,参照Qiime软件质量控制流程,将拼接后的序列经过截取、过滤得到有效数据.原始序列需经过拼接和质量过滤,为保障数据准确可靠,高质量数据需占到90%以上,为进一步分析物种组成多样性信息,使用 Uparse软件对所有样品的有效序列进行聚类分析,以 97%的一致性将序列聚类成为 OTUs种水平[20],剔除浮游动物OTUs数据后得到微藻注释信息.
1.6 数据分析
利用 Uparse(http://drive5.com/uparse/)对有效数据进行OTUs(Operational Taxonomic Units)聚类和物种分类,采用RDP Classifier方法与SILVA数据库(https://www.arb-silva.de/,Version 108)对OTUs代表序列进行物种注释[21].根据相关研究,不同粒径的真核微藻序列数比例更接近于生物量比例[13-16],将各个站位所获得的优势度超过 0.1%的种类均作为整体优势种参与粒级生物量比例统计,参考相关文献将筛查出的优势种粒径进行筛检分级[13-15,22-25],根据各粒径微藻序列占比统计其粒级结构,同时利用序列数占比和总叶绿素a含量可测算某种粒级微藻的叶绿素a浓度,相关公式:

式中:DBi为某粒级微藻的叶绿素 a浓度,μg/L;NBi为总叶绿素a浓度,μg/L;DC为某粒级微藻的序列数;NC为所有微藻的序列数.
物种生物量优势度(Y)表示微藻群落中某一物种质量所占的优势程度:

式中:nx为第x种微藻种类的OTUs数,N为OTUs总数,fx为第x种微藻种类在各样品中出现的频率.
本文的小、中、大粒径微藻分别代指微微型藻类(0.22~3μm)、微型藻类(3~20μm)及小型藻类(>20μm).
对上述公式计算、数据分析、分布饼图绘制均通过WPS Office、SPSS 19.0、ARGIS10.2软件完成.
2 结果
2.1 真核微藻各粒级优势种演替规律
黄海北部各粒级微藻第一优势种及优势度见表1,主要真核微藻粒级名录见表2(各站位所获得的序列数优势度超过0.1%的种类).参与统计的真核微藻总生物量占总检出真核微藻生物量的 97%以上,可代表整个真核微藻群落用于粒级结构分析.

表1 各粒级微藻第一优势种及优势度Table 1 The first dominant species and dominant degree of microalgae in each grain size
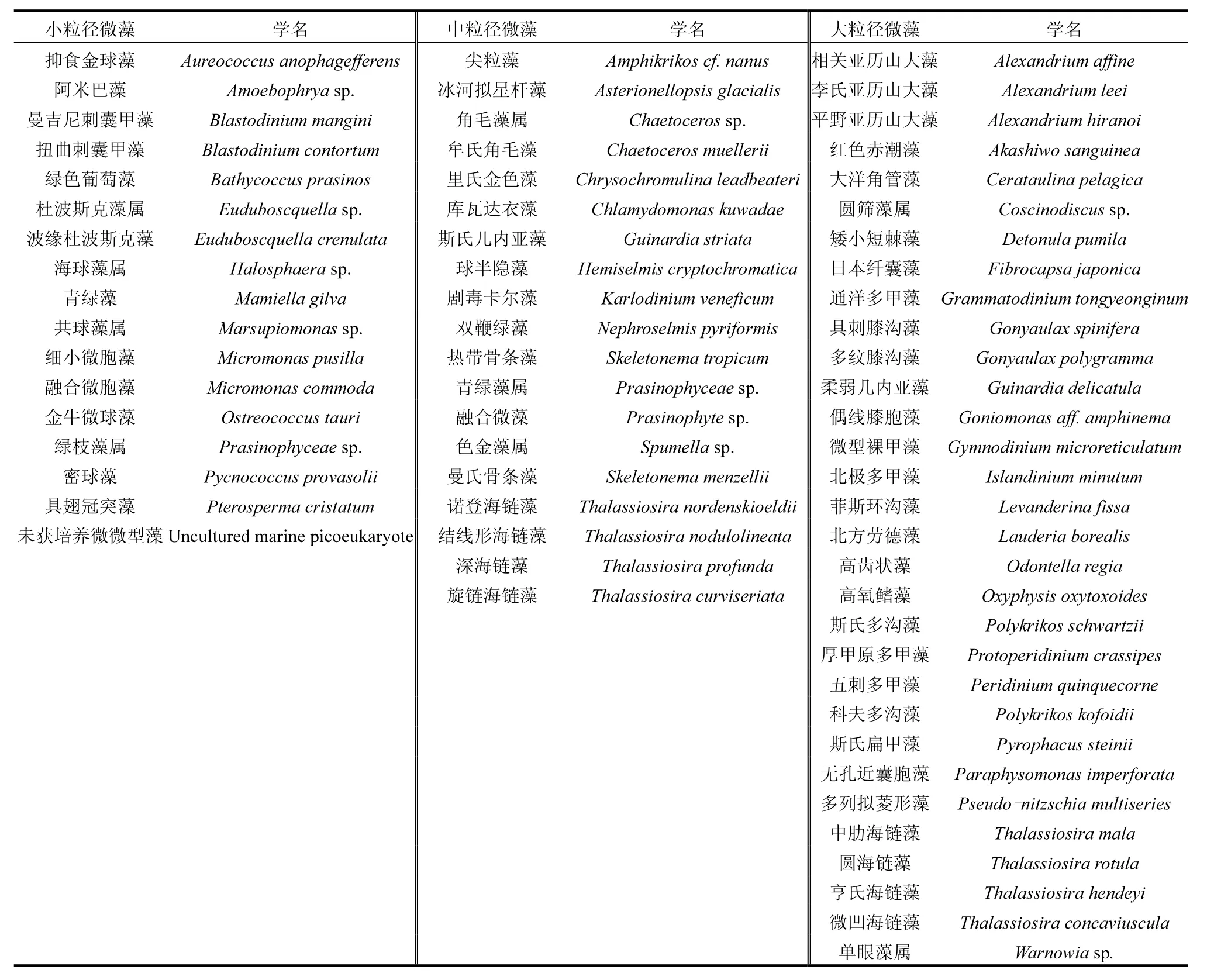
表2 主要微藻粒级名录Table 2 The granulation of dominant species of microalgae
其中,小粒级微藻春季细小微胞藻、融合微胞藻、曼吉尼刺囊甲藻在小粒级微藻群落中生物量优势度较高,分别为 0.183、0.159、0.111;夏季细小微胞藻、金牛微球藻、青绿藻生物量优势度较高,分别为0.290、0.229、0.121;秋季金牛微球藻、曼吉尼刺囊甲藻、细小微胞藻生物量优势度较高,分别为0.488、0.156、0.120.综合来看细小微胞藻和金牛微球藻在小粒级微藻群落中占有绝对优势,春夏秋季演替规律由细小微胞藻占优势转为金牛微球藻控制生态位.由于小粒级微藻粒径较小,细胞中 rDNA的拷贝数较少,推测其数量优势度在整个微藻群落中占有绝对优势.
中粒级微藻春季剧毒卡尔藻和三叶原甲藻在中粒级微藻群落中生物量占有绝对优势,优势度分别为0.429、0.141;夏季剧毒卡尔藻、虫黄甲藻、三叶原甲藻生物量优势度较高,分别为0.450、0.132、0.095;秋季剧毒卡尔藻和三叶原甲藻生物量优势度较高,分别为0.619、0.106.综合来看各季节剧毒卡尔藻在中粒级微藻群落中一直占有绝对优势.
大粒级微藻春季柔弱几内亚藻、北极多甲藻、矮小短棘藻、中肋海链藻、微型裸甲藻在大粒级微藻群落中生物量优势度较高,分别为0.330、0.181、0.120、0.119、0.105;夏季平野亚历山大藻和红色赤潮藻生物量优势度较高,分别为0.574、0.103;秋季多纹膝沟藻生物量占有绝对优势,优势度为0.882.大粒级微藻优势种在各季节演替较大,分别由不同种类占据主要生态位.
综合整个真核微藻种群优势分析,春季由中粒径的剧毒卡尔藻占有绝对优势,占比 21.3%,其他微藻占比均小于 7%;夏季由大粒径的平野亚历山大藻占有绝对优势,占比23.6%,其他微藻占比均小于11%;秋季由大粒径的多纹膝沟藻占有绝对优势,占比高达56.6%,其他微藻占比均小于 11%.由此可见,虽然整个微藻群落由大粒级微藻控制生态位,但都是有毒的甲藻,对黄海北部增养殖的贝类贝毒累积风险较高.
2.2 真核微藻粒级结构特征
真核微藻粒级结构分析结果显示,春季小粒级微藻组成分布在 27%~54%,平均(39±6)%,中粒级微藻组成分布在25%~67%,平均51±8%,大粒级微藻组成分布在 6%~22%,平均(11±2)%(图 2a).
夏季小粒级微藻组成分布在 8%~72%,平均(40±18)%,中粒级微藻组成分布在 2%~64%,平均(24±14)%,大粒级微藻组成分布在 8%~84%,平均(36±26)%,各站位分布差异比较大(图2b).
秋季小粒级微藻组成分布在 1%~89%,平均(26±19)%,中粒级微藻组成分布在 1%~34%,平均(13±7)%,大粒级微藻组成分布在 9%~99%,平均(62±18)%,各站位分布差异比较大(图2c).

图2 春、夏、秋季真核微藻粒级组分Fig.2 Size fractions of eukaryotic microalgae in spring,summer and autumn
整体而言,黄海北部春季真核微藻以中、小粒级为主,各站位粒级结构相对比较均衡,整体生物量较高(图3a),小、中、大粒级微藻比例为39:51:11;夏季以小、大粒级为主,各站位粒级结构波动较大,整体生物量较低,小粒级微藻主要分布在近岸海域,大粒级微藻主要分布在离岸海域(图 3b),小、中、大粒级微藻比例为 40:24:36;秋季除鸭绿江口附近海域以小粒级微藻为主外,其他区域主要以大粒级为主,各站位粒级结构相对比较均衡,整体生物量有所回升(图 3c),小、中、大粒级微藻比例为26:13:62.


图3 春、夏、秋季真核微藻粒级生物量分布Fig.3 Distribution of size biomass of eukaryotic microalgae in spring、summer and autumn
2.3 真核微藻粒级结构与环境因素关联
春、夏、秋季黄海北部真核微藻粒级结构与环境因子的相关系数见表 3.其中,春季与小粒级微藻生物量显著正相关的环境因素有悬浮物和活性硅酸盐,显著负相关的有水深、重金属汞、盐度、砷,其中与水深、重金属汞相关极显著;与中粒级微藻生物量显著正相关的环境因素有悬浮物、活性硅酸盐、化学需氧量、硝酸盐、无机氮,其中与悬浮物、活性硅酸盐相关极显著,显著负相关的有水深、盐度、重金属汞、镉,其中与前三项因子相关极显著;与大粒级微藻生物量显著负相关的环境因素有水深、重金属汞,达到极显著水平,无显著正相关环境因子.
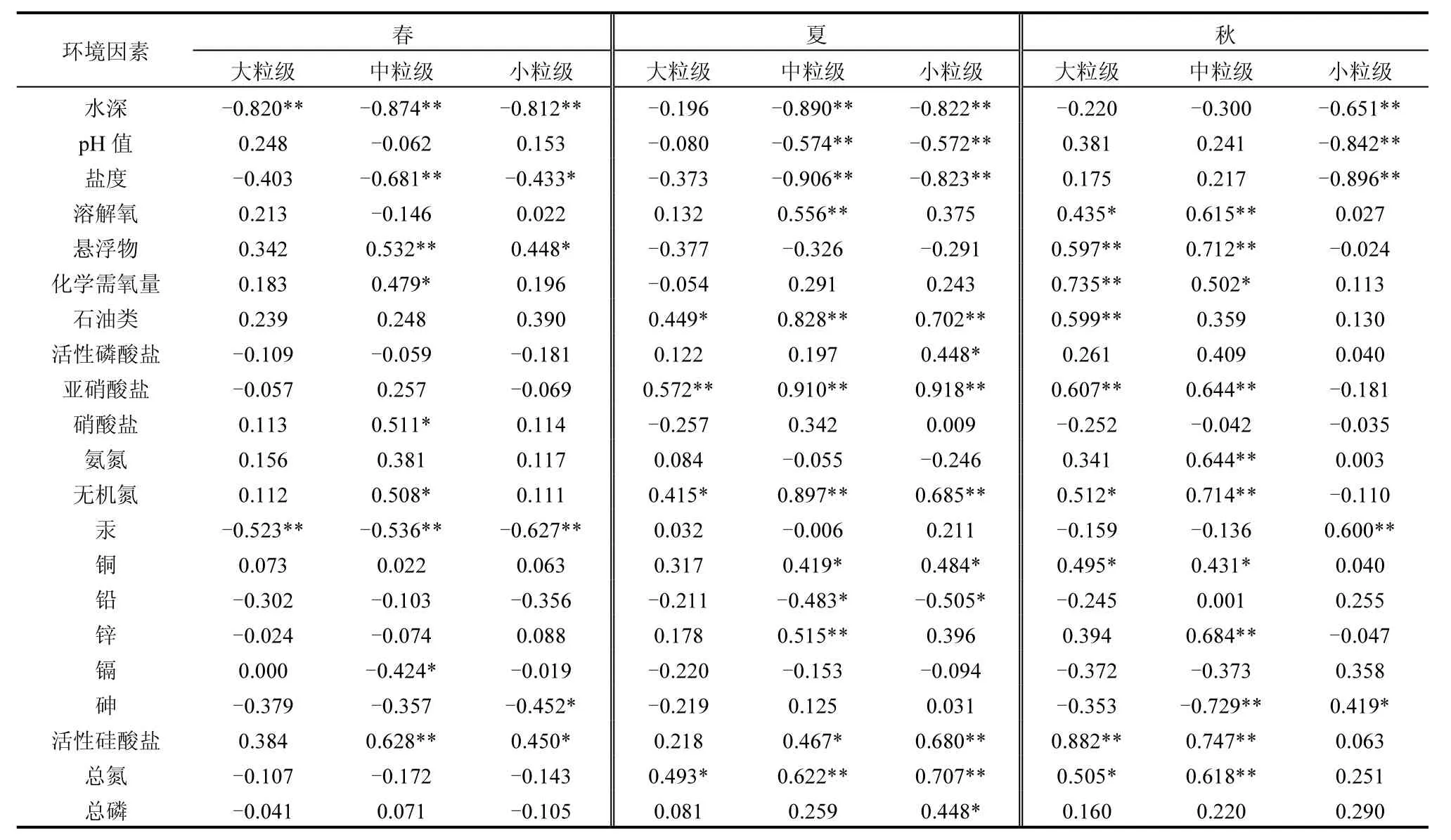
表3 微藻粒级组成与环境因子相关性Table 3 Correlation between microalgae particle size structure and environmental factors
夏季与小粒级微藻生物量显著正相关的环境因素有石油类、亚硝酸盐、无机氮、活性硅酸盐、总氮、总磷、活性磷酸盐、重金属铜,其中与石油类、亚硝酸盐、无机氮、活性硅酸盐、总氮相关极显著,显著负相关的有水深、pH值、盐度、重金属铅,其中与水深、pH值、盐度相关极显著;与中粒级微藻生物量显著正相关的环境因素有溶解氧、石油类、亚硝酸盐、无机氮、总氮、活性硅酸盐和重金属锌、铜,其中与溶解氧、石油类、亚硝酸盐、无机氮、总氮相关极显著,显著负相关的有水深、pH值、盐度和重金属铅,其中与前三项因子相关极显著;与大粒级微藻生物量显著正相关的环境因素有亚硝酸盐、无机氮、总氮、石油类,其中与亚硝酸盐相关极显著,无显著负相关环境因子.
秋季与小粒级微藻生物量显著正相关的环境因素有重金属汞、砷,其中与重金属汞相关极显著,显著负相关的有水深、pH值、盐度;与中粒级微藻生物量显著正相关的环境因素有溶解氧、悬浮物、亚硝酸盐、氨氮、无机氮、总氮、活性硅酸盐、化学需氧量和重金属锌、铜,其中除了化学需氧量和重金属铜,与其他因子相关极显著,与砷极显著负相关;与大粒级微藻生物量显著正相关的环境因素有悬浮物、化学需氧量、石油类、亚硝酸盐、活性硅酸盐、无机氮、总氮、溶解氧和重金属铜,其中与悬浮物、化学需氧量、石油类、亚硝酸盐、活性硅酸盐相关极显著,无显著负相关环境因子.
总体来看,水深、pH值、盐度、无机氮、总氮、活性硅酸盐及部分重金属与真核微藻粒级结构演变显著相关,直接影响不同粒级微藻的时空变动.
3 讨论
3.1 真核微藻粒级结构特征分析
本海域相关研究主要集中在长海县海域,2018年大长山岛临近海域小粒径真核微藻生物量组成春季为86%,夏季为52%,秋季为20%[16].本研究2019年长海县海域小粒径真核微藻生物量组成春季为38%,夏季为 72%,秋季为 39%.出现的差异主要是春季优势种发生演替,2018年春季优势种为抑食金球藻,生物量占41%,2019年春季优势种为细小微胞藻,生物量占18%,而抑食金球藻生物量仅占9%.夏、秋季优势种 2个年度均为细小微胞藻和金牛微球藻.吴文广利用叶绿素a分级法发现2015年该海域秋季小粒径(<2μm)微藻生物量占 2%,中粒径(2~20μm)占 65%,大粒径(>20μm)占 33%[3].由于叶绿素 a 分级法测算微藻粒级结构误差较大,可能会严重低估小粒径微藻的生物量组成[9,16],但也不排除调查时优势种演替及环境因子不同而造成的差异.
当然,利用测序所得的序列数比例表征生物量占比也存在偏差,但该方法与叶绿素 a分级法相比更接近实际,如在褐潮暴发微微型藻类占优势的区域叶绿素a分级法却显示微型藻类组成较高[24-26].
3.2 敏感微藻分布与贝类养殖风险分析
滤食性贝类主要摄食饵料微藻,而微藻营养(粒径大小)及毒性直接影响贝类生长与品质安全.本研究在黄海北部海域共发现 16种潜在褐潮微藻(<3 μm 小粒径微藻),其中金牛微球藻和抑食金球藻曾在渤海海域引发褐潮[19],并且这 2种褐潮微藻均会影响贝类摄食[25-26],滤食性贝类增养殖风险较高[27].长海县是我国虾夷扇贝主产区,近些年出现的大规模死亡现象可能与该海域饵料微藻粒级结构发生改变有关[16],关于微藻粒级小型化机制有待于进一步研究.另外,本研究海域虽然大粒径真核微藻在个别站位优势度较高,尤其在秋季占绝对优势,但主要优势种均为有毒甲藻,如亚历山大藻含有麻痹性贝毒,多纹膝沟藻和红色赤潮藻含有溶血毒素,具刺膝沟藻含有虾夷扇贝毒素,以及中粒径优势种剧毒卡尔藻,严重影响着增养殖贝类的食用安全[28].
3.3 微藻粒级结构的环境影响因素
相关研究发现,大长山岛海域与微藻粒级结构演替相关的环境因子主要为总氮、总磷、活性硅酸盐等[16],本研究也发现这几种因子与微藻粒级结构演替显著相关.还有研究发现无机氮是影响秋季獐子岛海域微藻生长和群落结构的主要因子,而硅酸盐可能成为影响大粒径微藻(>20μm)生长的重要因子[3],与本研究结论也基本一致.虽然各研究时期粒级结构不尽相同,但与环境因子关联结果总体一致.
由于调查海域站位间距较大,水深、pH值、盐度等有所差异,对中、小粒级微藻分布影响显著,但都是负向影响.已有研究表明,一般自养微藻都喜好无机营养盐[28-29],氮、磷、硅都是微藻生长必须元素[30],本研究海域无机氮、总氮、活性硅酸盐对微藻粒级结构也都是正向影响,部分重金属作为微量元素,适量范围内可刺激微藻生长,但超量则起到抑制作用,比如本研究中重金属汞春季抑制微藻生长,到了秋季则促进小粒级微藻生长,适量铜、锌可促进微藻生长,过量铅、镉、砷可抑制微藻生长,适量石油类也可促进微藻快速生长[31-36].
本研究与微藻粒级结构演替极显著相关的环境因子对各站位各季节微藻群落结构影响较大,各环境因子之间也相互耦合关联,水深较浅区域,营养盐和重金属含量相对较高,反之亦然.本研究海域水深较浅海域,小粒级微藻含量较高,除受环境因子的上行效应影响之外,贝类及浮游动物大量摄食的下行效应也会影响微藻的粒级结构[2].黄海北部海域主要增养殖对象为杂色蛤、扇贝、牡蛎等滤食性贝类,对天然饵料微藻需求量较大.虽然鸭绿江、大洋河等每年携带陆源氮磷等营养盐入海,但随着入海河流污染整治,氮磷消减力度加大,海域无机营养盐已呈下降趋势,对微藻生物量及粒级结构影响较大.
4 结论
4.1 黄海北部真核微藻春季以中(51%)、小粒级(39%)为主,夏季以小(40%)、大粒级(36%)为主,秋季以大粒级(62%)为主.
4.2 小粒径微藻中金牛微球藻和抑食金球藻曾在渤海引发褐潮,潜在威胁贝类养殖业.另外有毒甲藻在该海域中占有绝对优势,中粒径的剧毒卡尔藻在春夏秋季优势明显,优势度在 0.4以上,贝毒累积风险较高.
4.3 水深、pH值、盐度、无机氮、总氮、活性硅酸盐及部分重金属等环境因素对黄海北部真核微藻粒级结构影响较大.
猜你喜欢
矿冶工程(2022年6期)2023-01-12
古今农业(2022年1期)2022-05-05
现代畜牧科技(2021年8期)2021-10-13
科学技术创新(2021年22期)2021-08-16
昆明医科大学学报(2021年5期)2021-07-22
水泥工程(2020年3期)2020-10-28
生命科学研究(2018年1期)2018-05-29
环境保护与循环经济(2017年3期)2017-09-26
食品工业科技(2014年23期)2014-03-11
中国医学科学院学报(2013年6期)2013-03-11
