
广西部分地区牛和羊梨形虫流行情况调查及鉴定
2021-03-29 01:46周庆安黄胜斌莫胜兰胡丽萍韩银华何奇松钟华训李剑静郭建刚
动物医学进展 2021年3期
周庆安,黄胜斌,莫胜兰,胡丽萍,韩银华,何奇松,马 琳,钟华训,李剑静,李 军,郭建刚*
(1.广西壮族自治区动物疫病预防控制中心,广西南宁 530001;2.岑溪动物疫病预防控制中心,广西梧州 543200)
梨形虫(Piroplasma)属原生动物界(Protozoan)、顶复门(Apicomplexa)、孢子虫纲(Sporozoea)、梨形虫亚纲(Piroplasmia)、梨形虫目(Piroplasmida)。梨形虫病为牛、羊等动物循环系统的寄生虫病,也是一种动物源性人兽共患病,包括巴贝斯虫病和泰勒虫病。巴贝斯虫病由巴贝斯虫科(Babesiidae)巴贝斯虫属(Babesia)的种类寄生于牛、羊的血液而发生原虫病,常见的种类有双芽巴贝虫(B.bigemina)、牛巴贝斯虫(B.bovis)、卵形巴贝斯虫(B.ovata)、莫氏巴贝斯虫(B.motais)[1]。而泰勒焦虫病由泰勒科(Theileriidae)泰勒虫属(Theileria)的种类寄生于牛、羊的巨噬细胞、淋巴细胞和红细胞而发生原虫病,牛、羊等家畜常见的泰勒虫种类有[1-3]环形泰勒虫(T.annulata)、中华泰勒虫(T.sinensis)、东方泰勒虫(T.orientalis)、小泰勒虫(T.parva)、绵羊泰勒虫(T.ovis)、羊泰勒焦虫(T.hirci)吕氏泰勒虫(T.lunwenshuni)和尤氏泰勒虫(T.uilenbergi)。在一些养殖地区,牛、羊群体发生梨形虫病[4-7],临床症状主要以高热、贫血和消瘦为主,发展为慢性病后,严重影响牛、羊生产,阻碍牛、羊产业健康发展,造成经济损失。
传统的检测方法为血液涂片染色镜检,耗时、费力,受主观因素影响较大。本次运用套式PCR方法,根据巴贝斯虫和泰勒虫18S rRNA 基因序列设计的通用引物。18S rRNA基因是编码真核生物核糖体小亚基的DNA序列,其中有保守序列区域,也有可变序列区域。保守区体现了种之间的亲缘关系,而可变区能区分种间的差异,可用于种属以上的分类分子标志。近年来,牛、羊产业是广西重点扶贫产业,也是部分地区重要畜牧产业中支柱产业,是农民增加收入重要来源之一。梨形虫病是影响牛、羊产业健康发展的重要因素,同时广西对牛、羊的梨形虫病调查较少,因此,本次运用套式PCR方法对牛、羊全血梨形虫开展分子流行病学调查,为广西梨形虫病的防治提供科学依据。
1 材料与方法
1.1 材料
1.1.1 样品 2019年1月至2019年12月根据各市养殖情况在广西南宁市、柳州市、河池市、贺州市、百色市5个地区的牛、羊养殖场进行颈静脉采集全血,置于EDTA真空抗凝管,混均匀后,冷藏保存,带回实验室检测(表1)。

表1 牛、羊全血采集信息表
1.1.2 主要试剂及仪器 TGuide S32磁珠法血液基因组DNA提取试剂盒、2×Taqplus PCR Master mix、大量琼脂糖凝胶DNA回收试剂盒、DH5α感受态细胞、质粒小提试剂盒购于北京天根生化科技有限公司;PMDTM18-T克隆载体、DL 2000 DNA Marker购于Takara公司;TGuide S32核酸抽提仪、Bionetra TONE PCR仪、凝胶成像系统、电泳仪购于北京六一生物科技有限公司。
1.1.3 引物合成 参照文献根据巴贝斯虫和泰勒虫18S rRNA 基因序列信息设计的通用引物[8],用套式PCR检测牛羊血液携梨形虫原虫情况。引物序列由英潍捷基(上海)贸易有限公司合成。
1.2 方法
1.2.1 全血核酸抽提及序列扩增 取200 μL牛、羊全血用于核酸抽提,采用天根TGuide S32磁珠法血液基因组DNA提取试剂盒抽提。
用套式PCR方法扩增目的基因:第1~2轮扩增反应均25 μL体系,包括2×TaqMasterMix 12 μL,ddH2O 9 μL,上、下游特异性引物各0.5μL,DNA模板3 μL,第2轮PCR反应体系模板为第1轮PCR反应产物。PCR扩增程序:95℃ 5 min;94℃ 1 min,55℃ 1 min,72℃ 1 min,34个循环;72℃ 10 min。PCR产物用15 g/L的琼脂糖凝胶在100 V、66 mA条件下电泳,经30 min后,在凝胶成像系统观察是否有特异性亮带。
1.2.2 梨形虫序列测序 应用大量琼脂糖凝胶DNA回收试剂盒回收梨形虫目的基因,并在16℃条件下连接至PMDTM18-T载体后转化至DH5α感受态细胞。挑选可疑阳性菌落扩大培养,再回收重组质粒,经鉴定是阳性后,送至英潍捷基(上海)贸易有限公司进行测序。
1.2.3 序列分析和系统发育树的建立 将本次成功测序的梨形虫序列在NCBI中注册的序列进行同源性对比,分析所得的序列是否为目的基因以及梨形虫病原体种类,并和GenBank上登录的泰勒虫和巴贝斯虫18S rRNA基因序列相似度比较,选东方泰勒虫(KT124538、MH208642)、环形泰勒虫(MF287948、MG599091)、中华泰勒虫(MT271905)、尤氏泰勒虫(JF719835) 、鹿泰勒虫(KT959227)、绵羊泰勒虫(MN493111)、吕氏泰勒虫(LC326007)、牛巴贝斯虫(KY805833)、莫氏巴贝斯虫(AY260179)、卵形巴贝斯虫(MN900525)、双芽巴贝斯虫(MH050358)作为参照序列,选同为梨形虫目的Anthemosomagarnhami(MH093637)、猫胞簇虫(Cytauxzoonfelis,L19080)作为外类群,用MEGA5.2软件中的Neighbor-Joining Tree,经过 1000 次的 Bootstrap计算,构建系统发育树。
2 结果
2.1 梨形虫18S rRNA基因PCR扩增结果
经特异性引物扩增后,将PCR产物进行电泳后,在凝胶成像系统可观察到约438 bp片段亮带,与预期片段大小相符(图1)。

M.DNA标准 DL 2 000;“-”阴性对照;“+”阳性对照;1~7.PCR 产物
2.2 各地区牛、羊规模场梨形虫感染情况
本次检测在南宁、柳州、河池、百色、贺州5个地区均检测到梨形虫病原体,其中7个牛养殖场和10个羊养殖场检测到梨形虫病原体。共检测638份全血,梨形虫平均感染为24.76%,其中牛全血平均感染率为20.19%,羊全血平均感染率为24.19%(表2、表3)。

表2 广西部分地区养殖场携带梨形虫情况
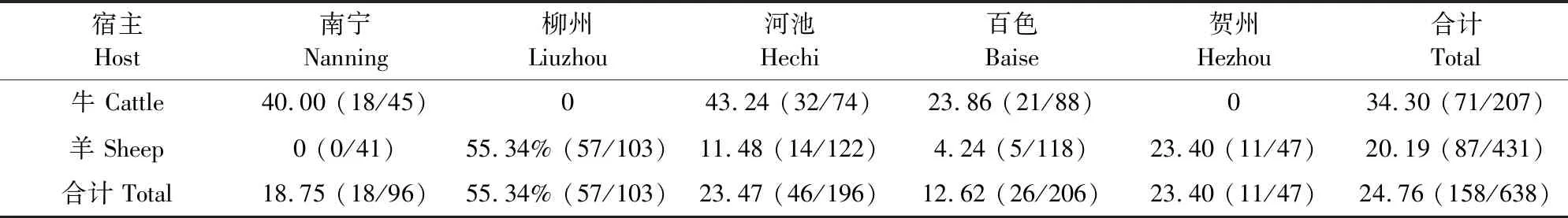
表3 广西部分地区牛、羊感染梨形虫情况
2.3 序列分析结果
检出牛、羊全血梨形虫病原体中,随机抽取部分阳性样品进行序列测序,在NCBI 中Blast相似性达97%以上,双芽巴贝斯虫3株、牛巴贝斯虫1株、环形泰勒虫4株、东方泰勒虫23株、吕氏泰勒虫21株。基于18S rRNA 基因序列构建系统进化树中得出泰勒虫属和巴贝斯虫属的种类分布在两个大分支,相应的种类分别与该病原相似度较高的参考序列在同一小分支(图2)。

图2 基于18S rRNA基因梨形虫不同种类的进化树
3 讨论
本次检测的638份全血覆盖南宁、柳州、河池、百色、贺州等地区32个养殖场,在5个地区中均检测出梨形虫,在20个养殖场中检测出梨形虫,平均感染率达到24.76%,涉及双芽巴贝斯虫、牛巴贝斯虫、环形泰勒虫、东方泰勒虫、吕氏泰勒虫5个种类,与云南省腾冲市牛羊梨形虫平均感染率为27.31%,涉及双芽巴贝斯虫、牛巴贝斯虫、吕氏泰勒虫、环形泰勒虫、东方泰勒虫等13个种类略相似[9];与四川省阿坝州牦牛和藏绵羊梨形虫平均感染率为49%,种类为吕氏泰勒虫和中华泰勒虫,策勒县和田羊和策勒黑羊梨形虫平均感染率为69.2% ,均为绵羊泰勒虫略有不同[10-11]。广西牛、羊梨形虫感染率较一些地区较低,感染种类也略有不同,但是以上调查结果表明广西仍然部分地区是梨形虫感染风险较高地区。梨形虫是动物源性人兽共患病,动物宿主包括家畜和野生动物,可以通过蜱虫传播媒介传播[12-13],如硬蜱既是梨形虫的传播媒介又是贮藏宿主,可在在动物宿主和硬蜱完成其生活史[14]。近年有巴贝斯虫感染人的事件,巴贝斯虫病又引起人们的重视和关注,蜱与巴贝斯虫关系密切,因此,蜱虫传播媒介也是以后要监测的对象。
牛主要以感染东方泰勒虫为主,在采样过程未发现携带有东方泰勒虫的牛表现临床症状。感染牛的泰勒虫分为良性和恶性泰勒虫2种,东方泰勒焦虫属于良性泰勒虫,致病性较低,不表现临床症状,感染的家畜处于带虫免疫状态[15]。有研究分析编码梨形表面蛋白(MPSP)的P32基因,可鉴定出11种基因型[16-18],其中感染2型(Ikeda)和少数1型(Chitose)可表现临床症状[19-23]。而羊以吕氏泰勒虫为主,与我国部分地区羊泰勒虫病中某些地区样品的结果相似[15],该种与原有种类的传播媒介、培养特性及分子对比后命名为吕氏泰勒虫(T.lunwenshuni),是羊泰勒虫致病力较强之一[2,3,24]。东方泰勒虫、吕氏泰勒虫分别是广西5个地区感染牛、羊的优势种,也具有致病力,因此,在日常监测和防治牛、羊寄生虫病中应重点关注。
系统发育树分析发现,检测到的3种不同泰勒虫,它们的18S rRNA基因序列分别分布在不同分支上,南宁、百色的环形泰勒虫、东方泰勒虫和河池的东方泰勒虫在泰勒虫属的大分支中形成独立大分支,而柳州和贺州的吕氏泰勒虫也在泰勒虫属的大分支中形成一个独立分支,分支里又有3个小分支,说明不同吕氏泰勒虫之间有差异,与文献[25]调查结果相似。
猜你喜欢
浙江社会科学(2022年4期)2023-01-09
当代水产(2022年1期)2022-04-26
现代畜牧科技(2021年11期)2021-12-21
现代畜牧科技(2021年8期)2021-10-13
科教新报(2021年11期)2021-05-12
成都医学院学报(2021年6期)2021-03-26
民间故事选刊·下(2020年3期)2020-04-24
作文评点报·中考版(2020年9期)2020-04-10
发明与创新·大科技(2018年10期)2018-03-18
渔业致富指南(2016年12期)2016-11-11
