
鸢尾属分子系统发育学研究进展
2021-03-24 18:30程琳奉树成肖月娥于凤扬朱淑霞王贤荣
广西植物 2021年1期
程琳 奉树成 肖月娥 于凤扬 朱淑霞 王贤荣

摘 要: 鸢尾属植物具有重要的观赏价值和经济价值,研究该属属内系统演化关系对该属种质保存、利用和遗传育种具有重要意义。目前,有关鸢尾属各分类单元的系统学关系長期处于争议状态。因此,该文以经典分类系统主要观点为基础,从不同分类单元的系统关系总结了鸢尾属分子系统发育学研究进展。结果表明:现有分子系统发育学研究结果大多支持鸢尾属为并系,且该属存在多个起源,属下的有髯鸢尾亚属、无髯鸢尾亚属均为多系,朱诺鸢尾亚属可分为5个分支,其中朱诺鸢尾组为多系。有髯鸢尾亚属和无髯鸢尾亚属下多个组的分类地位尚未确立。同时,该研究结果显示无髯鸢尾组下的加利福尼亚鸢尾系和金脉鸢尾亚系可能为复合种类群,从而导致近缘种种间形态特征界限模糊。此外,还确立了经典分类系统中部分存疑种的分类地位,如将射干和野鸢尾并入鸢尾属、将蛇头鸢尾和中甸鸢尾分别归入网脉鸢尾亚属和尼泊尔鸢尾亚属等。该文还对鸢尾属起源分布和进化历史等进行了展望,为今后鸢尾属的进一步研究提供了参考。
关键词: 鸢尾属, 分类, 分子系统发育, 进化, 基因组
Abstract: Iris plants have important ornamental and economic values. The study on the phylogenetic relationship of Iris is of great significance for the conservation, utilization and genetic breeding of Iris. At present, the systematic relations of the taxonomic units of Iris have been long in dispute. Thus, this paper reviewed the progress of molecular phylogeny of Iris from the systematic relations of different taxonomic units based on the traditional taxonomy system. The results were as follows: Iris is paraphyletic with multiple origins, while the Subgen. Iris, Subgen. Limniris are both polyphyletic. Subgen. Scorpiris can be divided into five branches, among which the Sect. Juno is polyphyletic. The Ser. Californicae and Subser. Chrysographes may be composite species groups, which blurred the boundary of morphologic features among sibling species of the two lineages. Some species with unclear taxonomic statuses in classical taxonomy has been confirmed, which showed that Belamcanda chinensis and Iris dichotoma should be incorporated into Iris. I. tuberosa and I. subdichotoma should be classified into Subgen. Hermodactyloides and Subgen. Nepalensis, respectively. We also prospected the related researches focusing on the origin, distribution and evolution history of Iris and put forward some suggestions on the research methods aiming to provide reference for further study.
Key words: Iris, classification, molecular phylogeny, evolution, genomes
鸢尾属(Iris L.)隶属于鸢尾科(Iridaceae),全世界约300种,主要分布于北半球温带地区(赵毓棠, 1985)。鸢尾属植物株型优美、花色丰富、花型奇特,具有重要的观赏价值,备受园艺界关注(胡永红和肖月娥, 2012)。同时,部分鸢尾属植物较易发生自然杂交和花色变异,是研究渐渗杂交与进化的代表物种之一(Arnold et al., 1990)。中国鸢尾属种质资源极为丰富,共约有60种,其中21种为特有种,鸢尾(Iris tectroum)、蝴蝶花(I. japonica)、马蔺(I. lactea var. chinesis)和溪荪(I. sanguinea)等种类在中国各地常见栽培。全世界10个鸢尾属植物分布中心的3个分布中心位于中国,即中国西南、西北和东北地区(Waddick, 1992)。
分子系统发育学根据生物大分子包含的遗传信息建立系统发育树,推断生物有机体间的遗传与进化关系,进而解决属间或属下各分类群之间的关系(李德铢, 2012)。鸢尾属植物作为世界重要观赏植物之一,鸢尾属系统发育学研究一直是该属研究热点问题。自Linnaeus于1735年创建鸢尾属以来,植物学家们依据不同的形态学特征如花中脉是否有附属物、根的形态、蒴果形态等,采用不同的分类单元如亚属、组、亚组等,建立鸢尾属不同的分类结果(Tausch, 1823; Spach, 1846; Dykes, 1913; Lawrence, 1953; Rodionenko, 1961; Mathew, 1989)。在这些分类系统中,Mathew(1989)的分类系统使鸢尾属及各亚属间系统关系已较为清晰,然而亚属下组、系以及部分种的分类地位仍存在争议。随着分子生物学技术的发展,人们开始通过分子系统发育学对鸢尾属内的关系进行探讨。但是由于研究方法、选用基因的不同导致研究结果不尽相同,因此有关鸢尾属内各分类单元的系统学关系长期处于争议状态。
鸢尾属分子系统发育学相关研究对该属种质的鉴定、保护、利用和遗传育种具有重要的理论意义和实际价值,也一直是鸢尾属研究的热点问题(Wilson, 2011;Mavrodiev, 2014)。本文以经典形态学分类系统为基础,综述鸢尾属分子系统发育学研究进展,归纳了目前已获得的主要结论和亟待解决的问题,并就该研究领域热点问题、方法和范围等提出了自己的观点或相应的建议,旨在为该领域后续研究提供参考。
1 鸢尾属概况及其系统分类
1.1 鸢尾属概况
在恩格勒分类系统中,鸢尾科隶属于被子植物门单子叶植物纲百合目(Liliflorae)百合亚目(Subordo Liliineae)(赵毓棠, 1985)。而在最新的APG IV系统中,鸢尾科被划到被子植物门单子叶植物纲天门冬目(Asparagales)。全世界鸢尾科植物有80属约1 600种,鸢尾属为该科模式属,也是鸢尾科中最大的属,全世界总计有鸢尾属植物300多种,集中分布于北温带的欧亚大陆和北美,中国至中东(古地中海)为其分化中心,最北分布到冰岛、斯堪的纳维亚半岛,最南分布在美国的佛罗里达。鸢尾属植物为多年生草本,该属典型形态特征为6枚花被片分为明显的内外两轮、柱头花瓣化以及具蜜腺的花被管等(Goldblatt & Manning, 2008)。
1.2 鸢尾属植物系统分类研究
自1735年Linnaeus首次创建鸢尾属以来,鸢尾属分类就一直受到人们的关注。1817年,Roamer和Schuites以外轮花被裂片是否有附属物将鸢尾属划分为3大类群。1823年,Tausch将鸢尾属划分为6个组,该系统成为了后人修订鸢尾属的基础。1846年,Spach修订了Tasuch的系统,将鸢尾属分为15个亚属。之后,陆续有Alefeld(1863)、Klatt(1872)、Bentham & Hooker(1883)依据繁殖器官特征、蒴果特征和根形态分别提出了相应的分类系统。1913年,Dykes将鸢尾属分为4个组,这是该属第一次较为科学的修订(Dykes, 1913)。1953年,Lawrence在Dykes的基础上,在鸢尾属下设置了亚属、组、亚组3个分类单元,为解决复杂的鸢尾属分类关系提供了帮助,鸢尾属植物分类进入成熟时期(Lawrence, 1953)。Rodionenko(1961)以根部是否膨大、花中脉是否有附属物等形态特征为依据将鸢尾属划分为6个亚属,并且将所有球根类鸢尾划分到鸢尾科不同的属,该系统首次将鸢尾属起源与系统发生纳入了该属分类依据。Mathew(1989)在其著作The Iris将鸢尾属划分为6个亚属,亚属下再设组,组下设系,该系统目前被广泛认可。
我国对于鸢尾系统分类的研究起步较晚。刘瑛(1936)发表的《中国之鸢尾》是我国最早关于鸢尾属植物的文献,记载了分布于我国的35种鸢尾。1980年,赵毓棠依据Rodionenko(1961)的分类系统,将我国的60种、13变种和5变型鸢尾属植物归入6个亚属,这成为我国延用至今的主要分类系统。Flora of China(Zhao et al., 2000)鸢尾科卷册对《中国植物志》中部分种进行合并,并增加了部分新发表的种类,总计描述了鸢尾属植物58种,其中21种为我国特有。
2 鸢尾属分子系统发育学研究
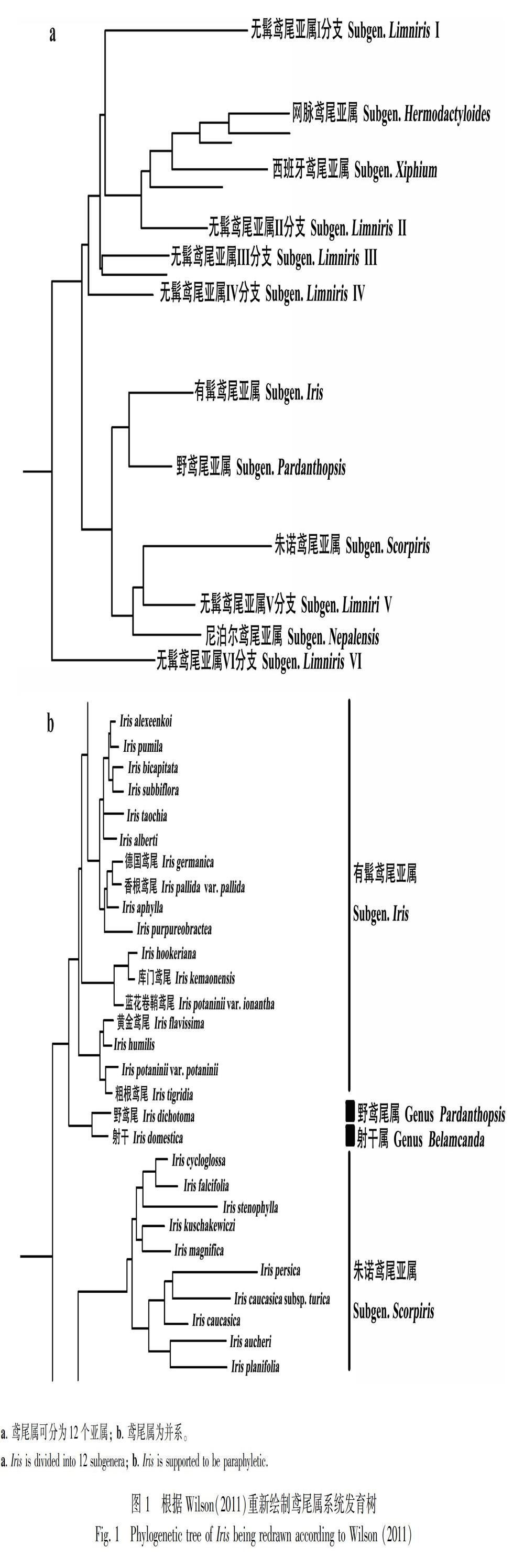
传统系统学以形态学、解剖学、孢粉学等作为分类依据,而分子系统发育学依据蛋白质或DNA序列推断生命有机体的系统演化关系(刘慧杰和谢磊, 2016)。20世纪末期,随着分子生物学的发展以及科学技术的更新,分子标记成为了创建系统发育分类系统的重要方法,并使鸢尾属分类进入后成熟发展时期(卢建红等, 2014)。Young(1998)采用2个叶绿体基因和3个限制性位点的对比,首次探讨了5种鸢尾属植物系统发育关系。Makarevitch et al.(2003)以分布于西伯利亚地区的22种鸢尾属植物为研究对象,利用叶绿体基因非编码间隔区序列和随机扩增多态性DNA分子标记分别构建系统发育树,并首次在基因库上提交了鸢尾属植物叶绿体基因序列,重建了西伯利亚地区鸢尾属分类系统。Wilson(2011)根据叶绿体基因序列建立的系统发育树,将全属初步分为12亚属,将传统分类系统中的无髯鸢尾亚属再细分为6个亚属(图1:a)。Movrodiev(2014)和Crespo et al.(2015)運用更长序列和更多样本建立了极广义鸢尾属系统发育树,分别建议将鸢尾属细分为23个属和25个属,将传统分类系统中某些种和亚属的地位提升为属的地位,如建立蝴蝶花属、 马蔺
属和溪荪属等,但该系统划分过细,多个属种数过少,造成应用不便,因此并未受到广泛认可。相比之下,Wilson(2011)提出的分类系统与传统分类系统的契合度更高,因此更被学术界认可。
目前,鸢尾属的分类地位已明确,Wilson(2004, 2011)研究结果表明鸢尾属为并系结构,该属有多个起源(图1:b)。但有关鸢尾属下部分亚属、组、系和种的分类地位仍存在争议。鸢尾属分子系统学研究现在主要集中在对无髯鸢尾亚属、有髯鸢尾亚属、朱诺鸢尾亚属3个亚属和一些重要存疑种上。
2.1 属内亚属分类地位
2.1.1 无髯鸢尾亚属(Subgen. Limniris)
2.1.1.1 亚属分类地位 目前,分子系统学研究结果一致支持无髯鸢尾亚属为多系,有多个起源。Tillie et al.(2000)通过叶绿体分子系统学研究结果表明无髯鸢尾亚属为单系,但支持率不高。Wilson(2009)采用叶绿体基因重新探讨了无髯鸢尾亚属的系统发育关系,结果表明无髯鸢尾亚属为多系,但其中有一个核心的物种群为单系,这个核心群包括Ser. Californicae、Ser. Sibiricae、Ser. Laevigatae、Ser. Tripetalae、Ser. Prismaticae、Ser. Ensatae、Ser. Ruthenicae、Ser. Hexagonae、Ser. Chinenses和Iris songarica,其中Ser. Sibiricae、 Ser. Tripetalae和Ser. Chinenses为多系,Ser. Laevigatae为并系。王殊(2018)对中国产鸢尾属部分种进行系统发育分析,发现无附属物组种间亲缘关系较复杂。蒋喻林等(2018)基于叶绿体基因对鸢尾属36个种进行发育树构建,结果表明无髯鸢尾亚属为多系,并可能有多个起源。
2.1.1.2 属下组分类地位 Rodionenko(1964)将无髯鸢尾亚属分为无髯鸢尾组(Sect. Limniris)和紫苞鸢尾组(Sect. Ioniris),赵毓棠(1985)在其基础上又新增单苞鸢尾组(Sect. Ophioris)。而Mathew(1989)将无髯鸢尾亚属分为无髯鸢尾组和冠饰鸢尾组(Sect. Lophiris),前者包括Rodionenko(1964)和赵毓棠(1985)系统中的无髯鸢尾组、紫苞鸢尾组和单苞鸢尾组种类,后者为Rodionenko(1964)系统中的冠饰鸢尾亚属(Subgen. Crossiris)。目前,由于针对性研究偏少,冠饰鸢尾组分类地位尚未确立,紫苞鸢尾组和单苞鸢尾组是否并入无髯鸢尾组也无定论。Wilson(2006)研究发现冠饰鸢尾组下的Iris wattii和I. cristata亲缘关系较远,Wilson(2009)进而发现I. tenuis和I. cristata二者并非单系。Rodionenko(1964)的紫苞鸢尾组含紫苞鸢尾(I. ruthenica)和单花鸢尾(I. uniflora)2种。王玲(2005)基于叶表皮微形态研究发现紫苞鸢尾、单花鸢尾与其他无髯鸢尾亲缘关系较远,支持Rodionenko(1964)的观点。现有分子系统学研究结果也支持将紫苞鸢尾和单花鸢尾作为独立种处理,且二者与其他无髯鸢尾植物亲缘关系较远(Wilson, 2009;仲轶, 2010;郑洋, 2016)。赵毓棠(1985)依据中国特有种单苞鸢尾(I. anguifuga) 生活史和苞片数量的特殊性将其独立为组,目前有关该种系统学研究未见开展。
2.1.1.3 组下分类地位 目前,有关无髯鸢尾组(Sect. Limniris)下系统发育研究集中在加利福尼亚鸢尾系(Ser. Californicae)和西伯利亚鸢尾系(Ser. Sibiricae)2个分类单元上。Foster(1937)认为加利福利亚鸢尾系至少包含3个分类群,Lenz(1958)则提出此系可按花被管长度分为2个亚群。Wilson(1998)根据叶绿体基因数据,认为加利福尼亚鸢尾系为单系。不同于Foster(1937)和Lenz(1958)结论的是,Wilson(2003)采用核糖体DNA的ITS序列进一步研究,发现加利福尼亚鸢尾系可分为4个支系,且Iris tenax和I. hartwegii 2个复合种并非先前认为的单系。Tillie et al.(2000)和Wilson(2011)基于叶绿体基因开展的分子系统学研究结果支持Mathew(1989)的观点,即将西伯利亚鸢尾系分为金脉鸢尾亚系(Subser. Chrysographes)和西伯利亚鸢尾亚系(Subser. Sibiriae)2个亚系。但仲轶(2010)和蒋喻林等(2017)基于多基因组研究和杂交实验发现西伯利亚鸢尾系物种及其近缘种的分类地位较混乱。
而有关无髯鸢尾组下琴瓣鸢尾系分类地位的争议已有了定论。Rodionenko(1964)将外轮花被片形似提琴状的一类鸢尾独立为1个亚属——琴瓣鸢尾亚属(Subgen. Xyridion),而Mathew(1989)将这些种类处理为无髯鸢尾亚属下无髯鸢尾组琴瓣鸢尾系(Ser. Spuriae)。Makarevitch(2003)、仲轶(2010)和王殊(2018)基于叶绿体基因数据的分子系统学研究结果均支持Mathew(1989)分类观点。此外,无髯鸢尾组下马蔺和白花马蔺(Iris lactea)分类地位仍未确定。赵毓棠(1985)将马蔺处理为白花马蔺的变种,该观点获得了余小芳(2009)基于形态学和ITS序列系统发育学研究结果的支持,但蒋喻林等(2017)基于叶绿体基因数据的分子系统学研究结果表明马蔺和白花马蔺亲缘关系较远,这可能是由于使用不同的分子标记而造成,需进一步探讨。
冠饰鸢尾组不同种间断分布于东亚与北美,这些间断分布种之间的演化关系尚未开展研究,有关该组下形态学上相似性高的扇形鸢尾(I. wattii)、扁竹兰(I. confusa)和蝴蝶花的分类地仍需进一步研究(余小芳, 2009;蒋喻林等, 2017)。
2.1.2 有髯鸢尾亚属(Subgen. Iris)
2.1.2.1 亚属分类地位 Tillie(2000)和Wilson(2004, 2011)分别采用叶绿体基因片段对有髯鸢尾亚属进行系统学研究,结果均支持Mathew(1989)的观点,即将有髯鸢尾亚属分成6个组(Sect. Iris、Sect. Psammiris、Sect. Oncocyclus、Sect. Regelia、Sect. Hexapogon、Sect. Pseudoregelia),但Sect. Psammiris、 Sect. Pseudoregelia和Sect. Regelia 3個组并非形态学分类所认定的单系。
2.1.2.2 属下组分类地位 目前,有关有髯鸢尾亚属下系统学相关研究集中在假种皮鸢尾组(Sect. Oncocyclus)。假种皮鸢尾组因其种子表面覆盖有白色附属物而得名,关于该组分类地位一直存在争议。Siemssen(1846)、Baker(1877)和Dykes(1913)分别将假种皮鸢尾处理为属、亚属和组。而在广为接受的Mathew(1989)分类系统中,假种皮鸢尾组隶属于有髯鸢尾亚属。Wilson(2016)基于1个低拷贝核基因和6个叶绿体基因数据的结果基本支持Mathew(1989)的观点,假种皮鸢尾组与Sect. Hexapogon为姐妹群,但Sect. Hexapogon部分种可移入假种皮鸢尾组。假种皮鸢尾组为单系,该组很有可能起源于高加索地区,而地中海东部地区是假种皮鸢尾多样性分布中心之一。
有髯鸢尾亚属中部分种分类地位仍不明确。比如,长白鸢尾(Iris mandshurica)因外轮花被片上具有须毛状附属物而被列入有髯鸢尾亚属,但其花粉形态和种苗形态等又与无附属物亚属植物相似,因此该种分类地位一直受到关注。王玲(2005)的UPGMA聚类树发现长白鸢尾和无附属物鸢尾亚属的黄菖蒲(I. pseudacorus)聚为同一支系,推断长白鸢尾可能是无附属物鸢尾亚属与有髯鸢尾亚属之间的过渡类型。但王殊(2018)基于叶绿体基因构建的系统发育树显示长白鸢尾处于有髯鸢尾亚属,支持现有分类地位。赵毓棠(1985)将大锐果鸢尾(I. goniocarpa var. grossa)和细锐果鸢尾(I. goniocarpa var. tenella)作为锐果鸢尾(I. goniocarpa)的变种处理,Flora of China将细锐果鸢尾并入锐果鸢尾,将大锐果鸢尾作为独立种。蒋喻林等(2017)认为锐果鸢尾和细锐果鸢尾的亲缘关系较大锐果鸢尾更近。而笔者在野外调查中发现锐果鸢尾、细锐果鸢尾、大锐果鸢尾在生境、植株高度、花大小和花色上存在明显差别。目前,有关这些种类的研究仍偏少,下一步可以有针对性地确立其分类地位。
2.1.3 朱诺鸢尾亚属(Subgen. Scorpiris)分类地位 Dykes(1913)和Lawrence(1953)分别将朱诺鸢尾作为组(Sect. Juno)和亚属(Subgen. Scorpiris)处理,而Rodionenko(1961)将朱诺鸢尾单独成属。但Mathew(1989)同意Lawrence的观点,即将朱诺鸢尾种类作鸢尾属内亚属处理,目前这一观点已获得了分子系统学研究结果一致支持(Ikinci et al., 2011;Wilson, 2004, 2009;Guo & Wilson, 2013)。Guo & Wilson(2013)采用叶绿体基因对外轮花被上具鸡冠状附属物的鸢尾种类进行了研究,结果显示尼泊尔鸢尾亚属(Subgen. Nepalensis)与冠饰鸢尾组聚成一支,之后再与朱诺鸢尾亚属构成姐妹群。Guo & Wilson(2013)研究发现朱诺鸢尾亚属内部可分为4分支,支持Ikinci et al.(2011)关于朱诺鸢尾组(Sect. Juno)为多系的观点。
2.2 部分种分类地位
2.2.1 野鸢尾(Iris dichtotoma)和射干(Belamcanda chinecsis) 射干与野鸢尾具有相似的形态与生境:高度分支,花较小,单朵花花期短,未受精的花会连同子房一起脱落;分布于干旱砂质草地、山坡石隙等向阳干燥处。野鸢尾和射干较易发生杂交(Yu et al., 2017)。传统的形态学分类中,野鸢尾被归入鸢尾属下的亚属(Subgen. Pardanthopsis)(Rodionenko, 1964)、组(Dykes, 1913)、亚组(Lawrence, 1953)中,或与射干合并为一个独立的属(Mathew, 1989)。目前,多数分子系统发育学研究结果显示尽管野鸢尾与鸢尾属下种亲缘关系较远,但仍支持将野鸢尾归入鸢尾属(Zhuravle et al., 1998;Tillie et al., 2000;仲軼, 2010;郑洋, 2016)。而Wilson(2004)基于叶绿体基因数据、王玲(2005)基于核基因(ITS序列)数据和王殊(2018)基于叶绿体数据均支持将射干归入鸢尾属。
2.2.2 蛇头鸢尾(Iris tuberosa) 蛇头鸢尾因具有肉质的指状地下器官以及单室的子房而有别于鸢尾属植物。Dykes(1913)、Lawrence(1953)、Rodionenko(1964)和 Mathew(1989)将蛇头鸢尾处理为单独的属。但是,Tillie et al.(2000)和Wilson(2011)基于叶绿体基因数据建议将该种归入鸢尾属网脉鸢尾亚属(Subgen. Hermodactyloides)。
2.2.3 中甸鸢尾(I. subdichotoma) 中甸鸢尾和野鸢尾形态较为相似,区别在于中甸鸢尾叶直立或稍弯曲,而野鸢尾叶弯曲,且中甸鸢尾花茎分枝较野鸢尾少,因此《中国植物志》将中甸鸢尾划分到野鸢尾亚属。但沈云光等(2004)通过形态学研究发现中甸鸢尾具有须毛状附属物、根肉质等特征,建议将中甸鸢尾归入尼泊尔鸢尾亚属,这一观点已获得仲轶(2010)和王殊(2018)等基于分子系统学研究结果的支持。
3 问题与展望
3.1 国产鸢尾重要类群系统发育地位亟需确立
系统学的基本任务是分类和命名,但另一重要功能是探讨物种演化历史的系统发育关系。通过演化研究可以了解植物与传粉者的关系、生物地理、环境适应、物种形成和演化速率等各种生物学现象。美国植物学家Carol Wilson有关鸢尾属系统发育的研究在深度和广度上都极具代表性。而国内研究多集中于属下具体种类分类地位的研究,忽略了有关我国鸢尾属起源与演化方面的探讨,建议在后续加强此方面研究,以此加大对我国鸢尾属种质资源的开发和可持续利用。
国产鸢尾中,有髯鸢尾亚属和无髯鸢尾亚属两大类群系统发生关系亟待解决。有髯鸢尾是由有髯鸢尾亚属有髯鸢尾组(Sect. Iris)的种类杂交获得的一个园艺类群,拥有数万个观赏品种,是鸢尾属中最为重要的观赏类群之一(胡永红和肖月娥, 2012)。赵毓棠(1985)分类系统认为中国原产有髯鸢尾亚属植物均隶属于果实侧裂组(Sect. Hexapogon),该组种类并未参与杂交选育。因此,未来应加强对中国产有髯鸢尾亚属系统发育研究,明确种间分类地位与亲缘关系,挖掘其优异基因。而《中国植物志》记载的60种国产鸢尾属植物中,31种隶属于无髯鸢尾亚属(约占总种数的51.7%),其中玉蝉花(Iris ensata)、燕子花(I. laevigata)和溪荪(I. sanguinea)等是鸢尾著名园艺类群的重要亲本(肖月娥和胡永红, 2012)。但是,赵毓棠(1985)分类系统对无附属物组多数种分类过于笼统,种间亲缘关系不清。而Mathew(1989)对无髯鸢尾亚属下组和系的分类更为细致,并得到了多数系统分类学研究结果的支持。因此,未来可参考Mathew(1989)分类系统,结合现代分子系统学最新分类结果,在国产无髯鸢尾亚属下设组、系,进一步将各分类单元细分,理清种间亲缘关系,以此更好地对该亚属进行开发利用。
3.2 部分复合种系统发育关系亟待理清
植物复合种是指由于杂交或其他手段使基因产生交流,形成在形态学上难以分辨的连续类群(刘晓贤, 2011)。鸢尾属部分近缘种之间可发生自然杂交而形成复合种(Wilson, 1998)。复合种的存在尽管会导致种间形态特征界限模糊,但是能为鸢尾这一重要园艺植物带来丰富的花色、花型变异,并且研究植物复合种对探讨与地理分布格局的遗传多样性变化及物种形成等具有重要意义。比如,目前有不少关于金脉鸢尾(I. chrysographes)、西南鸢尾(I. bulleyana)和西藏鸢尾(I. clarkei)分子系统发育学研究,但这些研究并未成功理清三者之间的系统发生关系。而笔者在野外考察中发现这3种染色体数目相同(2n = 40)的鸢尾属植物集中分布于中国西南地区,在自然生境中存在大量种间杂交过渡类型,推测倍性相同的三者可能是通过自然杂交形成复合种。未来,可先通过系统发育分析检验这3种在系统发育树上是否构成一个稳定的单系类群,然后再采用亲缘地理学研究方法对复合种的物种界定、种间遗传分化程度和进化历史等开展研究。
3.3 研究方法和研究范围亟待更新和扩大
目前,对于鸢尾属系统发育的研究多数建立在单个基因或几个基因片段构建的系统发育树上。当前使用的分子标记方法主要为叶绿体基因标记和少量的核基因标记。仲轶(2010)认为单一的叶绿体基因和核基因长度受限,信息位点较少,因此得到的鸢尾属植物的系统发育关系并不明确。因为不同的基因有不同的特点,如核基因的变异速率快,叶绿体基因信息量大、进化速率适中且单亲遗传,因此结合多基因开展系统发育学研究会得到更全面的拓扑结构(刘丽, 2017)。而全基因组包含着更为全面的进化信息,这可以为寻找物种进化标记物提供更多数据资料。当前全基因组测序技术已成为趋势(张树波和赖剑煌, 2010),建议在鸢尾属系统发育研究中推广多个基因标记相结合的方式以及全基因组测序技术,为鸢尾属研究获得更多的信息。
由于鸢尾属植物分布范围广泛、生境多样,较难收集到世界范围内鸢尾属植物,因此现有大部分学者将研究聚集在某个地区或聚焦于鸢尾属下某一分类单元,而取样范围受限和样本受限必然会无法获得全面的鸢尾属系统发育相关信息。因此未来应调查、梳理更大范围内鸢尾属植物种质资源,进一步扩大取样范围和数量,以此更好地探讨大区域范围内物种间的亲缘关系及演化关系。
参考文献:
ARNOLD ML, BOBBY BD, ZIMMER EA, 1990. Natural hybridization between Iris fulva and Iris hexagona: Pattern of ribosomal DNA variation [J]. Evolution, 44(6): 1512-1521.
CRESPO MB, MARTINEZ-AZORIN M, MAVRODIEV EV, 2015. Can a rainbow consist of a single colour? A new comprehensive generic arrangement of the ‘Iris sensu latissimo clade (Iridaceae), congruent with morphology and molecular data [J]. Phytotaxa, 232 (1): 1-78.
DYKES WR, 1913. The genus Iris [M]. New York: Dover Publications: 124-178.
FOSTER CA, 1937. Cyto-taxonomic survey of the North American species of Iris [J]. Harvard Univ Herb, 22(119): 1-82.
GUO JY, WILSON CA, 2013. Molecular phylogeny of crested Iris based on five plastid markers (Iridaceae) [J]. Syst Bot, 38(4): 987-995.
GOLDBLATT P, MANNING JC, 2008. The Iris family: natural history and classification [M]. Portland, Oregon: Timber Press: 200-204.
HU YH, XIAO YE, 2012. Wetland Irises: Appreciation, cultivation and application of cultivars [M]. Beijing: Science Press: 1-209. [胡永紅, 肖月娥, 2012. 湿生鸢尾: 品种赏析、栽培及应用 [M]. 北京: 科学出版社: 1-209.]
IKINCI N, HALL T, DOLORES LLEDO M, et al., 2011. Molecular phylogenetics of the juno irises, Iris Subgenus Scorpiris (Iridaceae), based on six plastid markers [J]. Bot J Linn Soc, 167: 281-300.
JIANG YL, HUANG Z, LIAO JQ, et al., 2018. Phylogenetic analysis of Iris L. from China on chloroplast TRNL-F sequences [J]. Biologia, 73(5): 459-466.
JIANG YL, LIU YJ, FENG YM, et al., 2017. Phylogenetic relationships among Iris species based on rbcL sequence [J]. Bull Bot Res, 37(3): 351-359. [蒋喻林, 刘宇婧, 冯艺玫, 等, 2017. 基于rbcL序列试论鸢尾属部分物种间的系统发育关系 [J]. 植物研究, 37(3): 351-359.]
LAWRENCE GHM, 1953. A reclassification of the genus Iris [J]. Gentes Herb: 346-371.
LENZ LW, 1958. A revision of the Pacific Coast Irises [J]. Aliso, 4: 1-72.
LI DZ, 2012. Plant systematic [M]. Beijing: Higher Education Press:1-38. [李德铢, 2012. 植物系统学 [M]. 北京: 高等教育出版社: 1-38.]
LIU HJ, XIE L, 2016. Advances in molecular phylogeny of Ranunculaceae [J]. Acta Bot Boreal-Occident Sin, 36(9): 1916-1924. [刘慧杰, 谢磊, 2016. 毛茛科分子系统发育研究进展 [J]. 西北植物学报, 36(9): 1916-1924.]
LIU L, 2017. Phylogeography of the endangered species Ulmus lamellosa (Ulmaceae) [D]. Linfen: Shanxi Normal University : 1-59. [刘丽, 2017. 濒危植物脱皮榆的谱系地理学研究 [D]. 临汾: 山西师范大学: 1-59.]
LIU Y, 1936. Iris of China [J]. Chin J Bot: 929-956. [刘瑛, 1936. 中国之鸢尾 [J]. 中国植物学杂志: 929-956.]
LIU XX, 2011. Studies on the phylogeny and phylogeography of Smilax china complex based on nrITS and cpDNA sequence variation [D]. Hangzhou: Zhejiang University: 1-75. [刘晓贤, 2011. 基于nrITS及cpDNA序列变异的菝葜复合种(Smilax china complex)系统发育及谱系地理学研究 [D]. 杭州: 浙江大学: 1-75.]
LU JH, PENG FC, ZHANG KL, 2014. Research and development of Iris classification system [J]. Jiangxi For Sic Technol, 42(5): 38-41. [卢建红, 彭方程, 章开灵, 2014. 鸢尾属植物分类系统的研究与发展 [J]. 江西林业科技, 42(5): 38-41.]
MAKAREVITCH I, GOLOVNIAN K, SCHERBIKET S, et al., 2003. Phylogenetic relationships of the Siberian Iris species inferred from noncoding chloroplast DNA sequences [J]. Int J Plant Sci: 229-237.
MATHEW B, 1989. The Iris [M]. London: Batsford: 89-156.
MAVRODIEV EV, MATINE-AZORIN M, DRANISHNKOV P, et al., 2014. At least 23 genera Instead of one: The case of Iris L. s.l. (Iridaceae) [J]. PLoS ONE, 9(8): 1-8.
RODIONENKO GI, 1987. The genus Iris. (English translation) [M]. London: The British Iris Society: 132-143.
SHEN YG, WANG ZL, GUAN KY, et al., 2004. Phylogenetic relationship of several species from Iris L. based on rbcL sequences [J]. Acta Bot Yunnan, 26 (5) : 487-492. [沈云光, 王仲朗, 管開云, 等, 2004. 中甸鸢尾的分类位置及其一新变型 [J]. 云南植物研究, 26 (5) : 487-492.]
SIEMSSEN CH, 1846. Ueber eine neue Gattung der Irideen [M]. Berlin: Botanische Zeitung: 705-710.
TILLIE NNMJD, CHASE MW, HALL T, 2000. Molecular studies in the genus Iris L.: A preliminary study [J]. Ann Di Bot, 58(7): 180-186.
WADDICK, 1992. Iris of China [M]. New York: Timber Press: 11-42.
WANG L, 2005. Studies on developmental biology and phylogenetic evolution of some Iris species [D]. Harbin: Northeast Forestry University: 1-90. [王玲, 2005. 鸢尾属部分种发育生物学与系统演化的研究 [D]. 哈尔滨: 东北林业大学: 1-90.]
WANG S, 2018. Phylogenetic studies on some species of Iris L. from China [D]. Changchun: Northeast Normal University: 1-50. [王殊, 2018. 中国产鸢尾属(Iris L.)部分物种的系统发生学研究 [D]. 长春: 东北师范大学: 1-50.]
WILSON CA, 1998. A cladistic analysis of Iris series Californicae based on morphological data [J]. Syst Bot, 23(1): 73-88.
WILSON CA, 2003. Phylogenetic relationships in Iris Series Californicae based on ITS sequences of nuclear ribosomal DNA [J]. Syst Bot, 28(1): 39-46.
WILSON CA, 2006. Patterns in evolution in characters that define Iris subgenera and sections [J]. Aliso, 22(1): 425-433.
WILSON CA, 2009. Phylogenetic relationships among the recognized series in Iris Section Limniris [J]. Syst Bot, 34(2): 277-284.
WILSON CA, PADIERNOS J, SAPIR Y, 2016. The royal irises (Iris subg. Iris sect. Oncocyclus): plastid and low-copy nuclear data contribute to an understanding of their phylogenetic relationships [J]. Taxon, 65(1): 35-46.
WILSON CA, 2004. Phylogeny of Iris based on chloroplast matK gene and trnK intron sequence data [J]. Mol Phylogenet Evol, 33: 402-412.
WILSON CA, 2011. Subgeneric classification in Iris re-examined using chloroplast sequence data [J]. Taxon, 60(1): 27-35.
YOUNG ND, 1998. Pacific coast Iris species delimitation using three species definitions: biological, phylogenetic and genealogical [J]. Evolution, 63: 99-120.
YU FY, XU WJ, XIAO EY, et al., 2017. Identifying apomixis in matroclinal progeny from an interspecific crossing between Iris domestica and three different colors of Iris dichotoma [J]. Euphytica, 213(12): 1-12.
YU XF, 2009. Systematics and seed dormancy and germination characteristics of Iris in Sichuan Province [D]. Yaan: Sichuan Agricultural University: 1-113. [余小芳, 2009. 四川鳶尾属植物的系统学及种子休眠与萌发特性研究 [D]. 雅安: 四川农业大学: 1-113.]
ZHAO YT, 1980. Studies on the genus Iris form China [J]. Acta Phytotax Sin, 18: 153-161. [赵毓棠, 1980. 国产鸢尾属的研究 [J]. 植物分类学报, 18: 153-161.]
ZHAO YT, 1985. Flora Reipublicae Popularis Sinicae, Vol. 16(1) [M]. Beijing: Science Press: 134-172. [赵毓棠, 1985. 中国植物志. 第16卷第1分册. [M]. 北京: 科学出版社: 134-172.]
ZHAO YT, NOLTIE HJ, MATHEW B, 2000. Iridaceae [M]//Flora of China. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press: 297-313.
ZHENG Y, 2016. Classification and evaluation of wild Iris in Northeast China [D]. Shenyang: Shenyang Agricultural University. [ 郑洋, 2016. 东北地区野生鸢尾属植物的分类与评价研究 [D]. 沈阳: 沈阳农业大学 ]
ZHONG Y, 2010. Systematic taxonomy of some Iris species based on multi-genome [D]. Haerbin: Northeast Forestry University: 1-168. [仲轶, 2010. 基于多基因组的鸢尾属部分种的系统分类研究 [D]. 哈尔滨: 东北林业大学: 1-168.]
ZHURAVLE YN, KOZYRENKO MM, ARTYUKOVAET EV, et al.,1998. Fingerprinting genomes of the far eastern species of the genus Iris L. by RAPD-PCR [J]. Russ J Genet, 34(3): 285-288.
ZHANG SB, LAI JH, 2010. Bioinformatics approach for molecular evolution analysi [J]. Computer Sci, 37(8): 47-52. [张树波, 赖剑煌, 2010. 分子系统发育分析的生物信息学方法 [J]. 计算机科学, 37(8): 47-52.](责任编辑 何永艳)
猜你喜欢
中国典型病例大全(2022年7期)2022-04-22
科学导报(2021年3期)2021-02-22
小天使·一年级语数英综合(2017年11期)2017-12-05
初中生世界·七年级(2017年9期)2017-10-13
少儿科学周刊·儿童版(2017年3期)2017-06-29
出版广角(2016年14期)2016-12-13
少儿科学周刊·少年版(2015年3期)2015-07-07
