
碎米藓属两种藓类植物孢子萌发与原丝体发育特征研究
2021-03-24 18:30黄士良曹珍张浩牛玉璐王振杰赵建成
广西植物 2021年1期
黄士良 曹珍 张浩 牛玉璐 王振杰 赵建成

摘 要: 为获取其孢子萌发类型与该属植物系统发育、生态选择以及生殖策略选择的相关性,该研究通过室内人工培养的方式,在微米量级下观察并描述了碎米藓属(Fabronia)碎米藓(F. pusilla)和东亚碎米藓(F. matsumurae)两种藓类植物孢子萌发、原丝体发育和配子体发生的过程。结果表明:(1)两种藓类植物孢子均为壁外萌发,均产生由1~15个半圆球形细胞组成的绿丝体(chloronema)短枝;(2)碎米藓在绿丝体顶端分化产生轴丝体细胞,东亚碎米藓未分化产生轴丝体(caulonema);(3)两种藓类植物配子体原始细胞均在绿丝体上分化产生。参照Nishida对藓类植物孢子萌发型划分标准,分析并确定了碎米藓属两种藓类植物孢子萌发型均为蓑藓型(Maromitrium-type),为碎米藓属的系统分类提供了发育学证据。
关键词: 碎米藓, 东亚碎米藓, 孢子萌发, 原丝体发育, 配子体发生, 孢子萌发型
Abstract: The spore germination and protonema development of two Fabronia moss species, F. pusilla and F. matsumurae, were observed and described at the micron level by means of indoor artificial cultivation in order to obtain the correlation between spore germination type and phylogeny, ecological selection and reproductive strategy selection of this genus. The results were as follows: (1) The spores of both species germinated outside the wall and produced chloronema twigs that composed of 1-15 semicircular cells; (2) Caulonema cells of F. pusilla were differentiated and formed from the top of chloronema whereas caulonema cells were undifferentiated in F. matsumurae; (3) Gametophyte initial cells of both species were differentiated and formed from chloronema. On the basis of previous studies, in this experiment, referring to Nishidas classification standard for moss spore germination, spore germination of the two Fabronia Raddi moss species were analyzed and determined to be of Maromitrium-type, providing developmental evidence for the systematic classification of this genus.
Key words: Fabronia pusilla, Fabronia matsumurae, spore germination, protonema development, gametophyte development, sporeling-type
藓类植物生活史中的孢子萌发阶段、原丝体发育阶段和配子体发生阶段是单倍体阶段,不同种类的藓类植物在单倍体阶段出现的性状特征存在差异性(Nishida,1978;胡人亮,1987)。目前,国内外对藓类植物原丝体发育、配子体发生的相关研究还比较薄弱。国外Hedwig和Nishida是对藓类植物原丝体发育研究比较早的两位专家,他们通过观察原丝体发育规律,描述了属于13种萌发孢子型的121种藓类植物的孢子萌发与原丝体发育情况,其中有属于26科47属的58种藓类植物的孢子萌发类型是首次报告。Nishida(1978)还研究了不同孢子萌发类型之间的关系,生态适应与孢子萌发类型的关系,孢子萌发类型对苔藓植物各类群分类的作用等,这为藓类植物原丝体的发育研究奠定了基础;Duckett & Ligrone (1994)在研究夭命藓(Ephemerum serratum)原丝体的形态发生过程中,进行了生长调节素干预实验,发现一定浓度的生长素对原丝体发育有正向促进作用,并在后续研究中继续观察了几种苔藓植物原丝体发育特点。之后,Pressel et al. (2005)继续研究夭命藓科(Ephemeraceae)夭命藓属(Ephemerum Hampe.)和细蓑藓属(Micromitrium Austin.)八种藓类植物原丝体发育特征,分析了八种藓类植物原丝体发育特点与生态选择的相关性。Katsumata et al.(2011)研究了细胞色素P450单加氧酶CYP78A亚家族在苔藓原丝体生长和配子体形成中正向促进作用。国内最早对苔藓植物原丝体发育进行系统研究的学者是高谦和张钺,他们研究了真藓亚纲(Bryidae)中红蒴立碗藓(Physcomitrium eurystomum)、角齿藓(Ceratodon purpureus)等九种藓类植物孢子萌发和原丝体发育特点,结果发现藓类原絲体阶段适应性相当广泛(高谦和张钺,1986)。他们还发现刺边葫芦藓(Funaria muehlenbergii)、角齿藓的孢子散落在培养液中也能发育成典型的原丝体系统,发育成熟的原丝体系即悬浮于培养液中,并能在原丝体上形成正常的芽体(高谦和张钺,1986)。2000年以来,苔藓研究学者先后对钟帽藓(Venturiella sinensis)、小石藓(Weissia controversa)、波叶仙鹤藓(Atrichum undulatum)等40多种藓类植物孢子萌发和原丝体发育特征进行了研究,新增了大帽藓型(Encalypta-type)和墙藓型(Tortula-type)2种新的孢子萌发类型(赵建成等,2002;刘保东和丛迎芝,2003;黄士良等,2015)。上述相关的研究探索为中国藓类植物发育学研究提供了重要的基础资料。
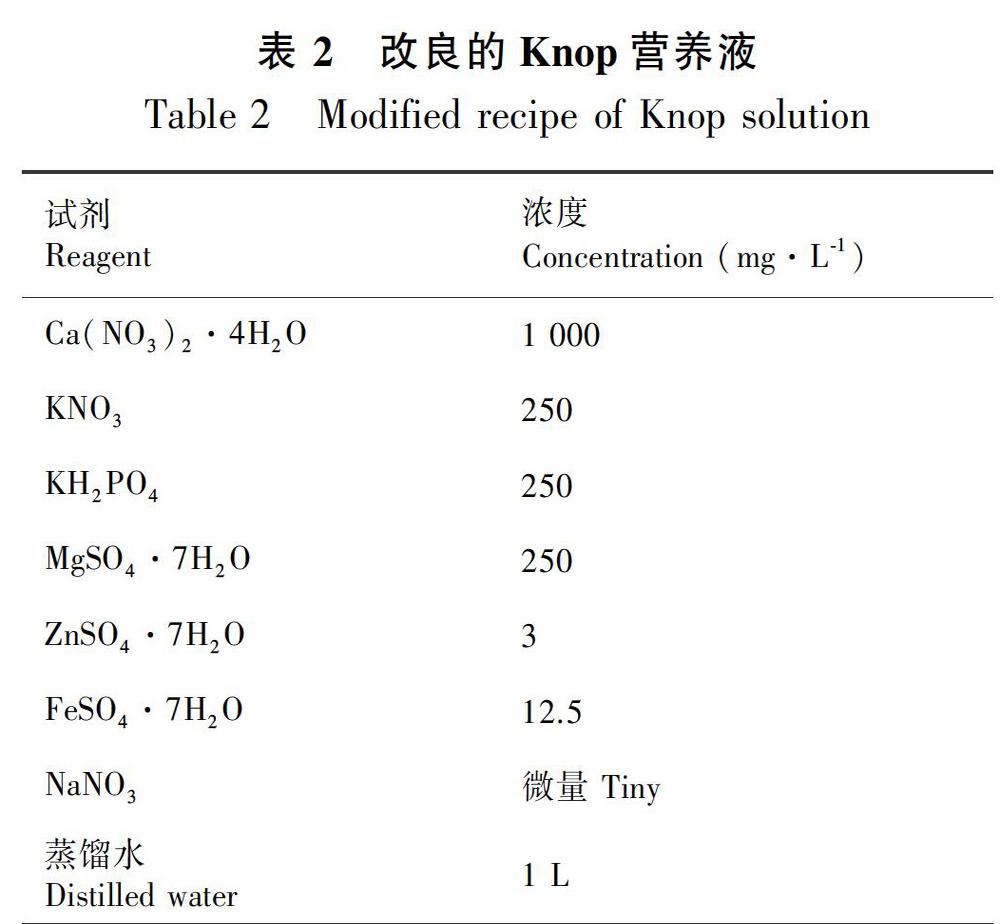
碎米藓(Fabronia pusilla)和东亚碎米藓(F. matsumurae)隶属碎米藓科(Fabroniaceae)碎米藓属(Fabronia Raddi),该属在国内分布有11种,生境主要以树附生为主(吴鹏程和贾渝,2004;贾渝和何思,2013)。树附生藓类植物因其生境的特殊性,其生活史中单倍体和二倍体的交替周期也受到生境的影响。目前,国内对碎米藓属植物孢子萌发、原丝体发育和配子体发生的系统性研究还未见报道。本研究所用的两种碎米藓属植物均为树生,通过观察其单倍配子体阶段的发育过程,以期探讨树附生藓类植物在孢子萌发、原丝体发育和配子体发生阶段的发育特征,及其对干旱环境适应产生的独特的性状特点。该研究将对碎米藓属植物分类学研究提供发育学资料,并且对揭示藓类植物对干旱的适应机理有着重要理论意义。
1 材料与方法
1.1 材料
所用材料取自碎米藓属两种植物成熟孢蒴内的孢子(表1),凭证标本存放于河北师范大学植物标本馆(吴鹏程和贾渝,2004;贾渝和何思,2013)。
1.2 方法
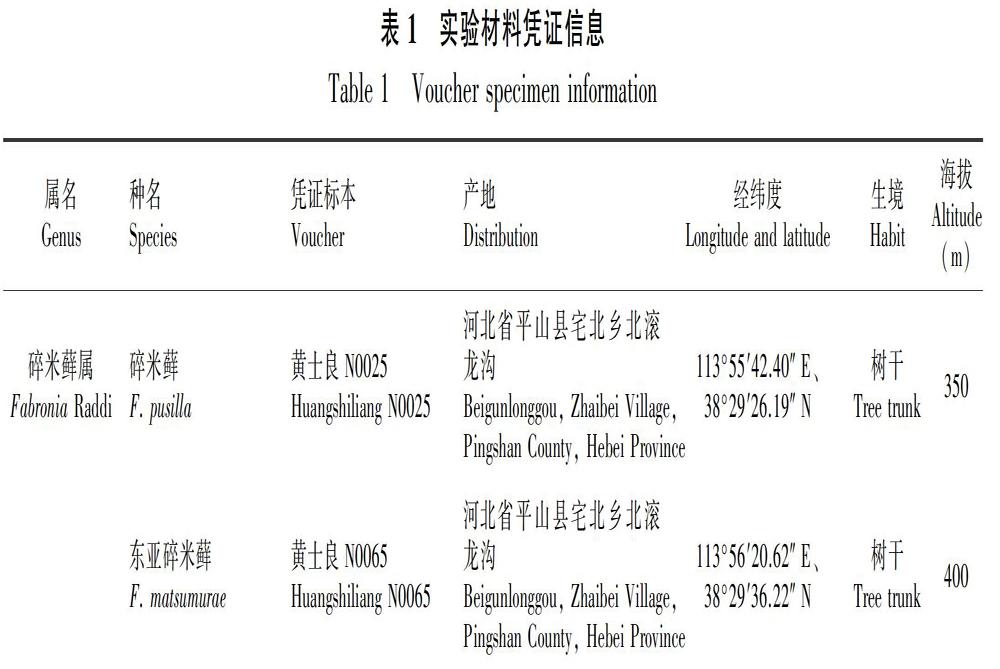
采用含有改良Knop营养液的琼脂培养基(范庆书等,2004;黄士良等,2015),基质pH 7.0,改良Knop营养液配方见表2。
1.3 实验步骤
从两种藓类植物标本上随机选取成熟孢蒴后,按照以下步骤进行操作(范庆书等,2004;赵建成等,2005;黄士良,2009)。
1.3.1 培养基的制备 实验组按表2配方,在一定量的改良Knop营养液中加入适量琼脂,使其浓度为2%,加热熔化后,倒入60 mm的培养皿中,凝固后即可使用。
1.3.2 孢子悬液的制备 分别把两种藓类植物的孢蒴浸入75%的酒精溶液中,20 s后取出,并在无菌蒸馏水中清洗5次;用镊子和解剖针将孢子取出并散入无菌蒸馏水中,制成适当浓度的孢子悬液。
1.3.3 孢子悬液转移 用移液器将稀释的孢子悬液接种于培养基上。
1.3.4 相关参数 培养皿加盖后放在光照培养箱(产品型号: KRG-250A)中培养。温度(23±2)℃,相对湿度大于80%,光照强度24 μmol·m-2·s-1,光照时间12 h·d-1。
1.3.5 镜检要求 自接种培养之日起,每隔24 h镜检孢子发育情况,每次镜检标本量为15~20个孢子的发育情况,详细记录个体的发育状况,并进行显微照相及绘图。
2 结果与分析
显微镜下观察两种碎米藓属植物的孢子均为球形或椭圆形黄绿色单细胞孢子,直径为10~15 μm。通过室内人工培养,它们的生活史中单倍配子体发育分为孢子萌发过程、原丝体发育过程和配子体发生过程三个阶段。
2.1 孢子萌发过程
碎米藓属两种藓类植物的孢子吸水膨胀萌发过程可分为两个时期:(1)孢子吸水膨胀期。室内接种孢子12 h显微镜下观察,两种藓类植物孢子内叶绿体颜色均加深,体积变化开始缓慢增加。24 h观察,两种藓类植物孢子内叶绿体的数目均不断增多,碎米藓孢子直径达到17~20 μm(图版Ⅰ:1);东亚碎米藓孢子吸水后直径达到15~18 μm(图版Ⅰ:2)。2 d时,东亚碎米藓孢子直径达30~40 μm,碎米藓孢子直径变化不明显。(2)破壁萌发期。培养3 d时,两种藓类植物孢子外壁均开始破裂,分化形成绿丝体原始细胞(chloronema initial cell)(图版Ⅰ:3,4)。5 d时,两种藓类植物萌发速度加快,萌发极向均为1~2极。碎米藓萌发率为75%,两极萌发率为28.1%(图版Ⅰ:5);东亚碎米藓萌发率为73.4%(图版Ⅰ:6)。8 d时,碎米藓和东亚碎米藓的两极萌发率分别为68.1%和65.5%,两种藓类植物孢子都开始出现三极萌发生长,孢子分化产生的绿丝体原始细胞生长缓慢,其分裂产生的第一个绿丝体细胞测量长度为20~25 μm,直径为10~15 μm,形状为半圆球形(图版Ⅰ:7,8)。9 d时,两种藓类植物孢子全部萌发,均没有产生初生假根(primary rhizoid)和初生轴丝体(primary caulonema)。 两种藓类植物在孢子萌发阶段的典型特征为产生半圆球形的绿丝体原丝体细胞。
2.2 原丝体发育过程
11 d时,两种藓类植物原丝体生长速度依然十分缓慢,绿丝体均由1~10个半圆球状细胞组成,形状如念珠状结构,长度为100 μm。15 d时,两种藓类植物均在绿丝体基部第1至第5个细胞上,开始分化产生一级分枝,产生的一级分枝以绿丝体原始细胞为中心,向四周散射生长,生长速度快,绿丝体分枝细胞的形状也为半圆球形(图版Ⅰ:9,10)。18 d时,碎米藓绿丝体基部细胞分枝速度加快,同时在绿丝体顶端细胞处分开始化形成轴丝体。新分化产生的轴丝体细胞由半圆球状细胞快速分化形成长度为600 μm的长圆柱状细胞。轴丝体未分化产生分枝,测量碎米藓的原丝体系统散射生长形成的圆形直径约为1 000~1 500 μm。绿丝体长度基本保持在300~350 μm(图版Ⅰ:11)。東亚碎米藓半圆球状绿丝体细胞生长缓慢,和碎米藓一样,基部绿丝体细胞开始大量分枝,新产生的分枝细胞和原来的绿丝体交叉生长,伸长生长速度缓慢, 新分枝的绿丝体顶部细胞保持半圆球状细胞形状,没有分化产生轴丝体细胞(图版Ⅰ:12)。在原丝体发育后期,碎米藓属两种藓类植物基部绿丝体开始大量分枝,分枝顶端为具有斜壁细胞的轴丝体(图版Ⅰ:13),这种原丝体系统在多维度上高度交叉生长,而没有扩散生长去增加原丝体系统的面积。在原丝体系统高密度生长的同时,其生长速度也在加快(图版Ⅰ:14)。在原丝体发育阶段,两种藓类植物原丝体均产生了由1~15个细胞组成的原丝体短枝,短枝均以绿丝体原始细胞为中心,向四周散射生长。而且两种藓类植物原丝体系统生长半径增长的速度缓慢。
2.3 配子体发生过程
20 d时,两种藓类植物均在绿丝体基部细胞处开始分化产生配子体原始细胞(gametophyte initial cell),细胞直径达100~150 μm,内含较多叶绿体(图版Ⅰ:15,16)。23 d时,两种藓类植物的配子体原始细胞逐渐分化成为桑葚状群体细胞结构,以不对称分裂(asymmertric division)的方式进行快速分化产生分生细胞团(meristem)。碎米藓的桑葚状群体细胞结构基部分化产生丝状假根(rhizoid),假根呈正向地性,假根生长速度很快(图版Ⅰ:17);东亚碎米藓桑葚状群体细胞结构基部未分化产生假根。25 d时,两种藓类植物的幼小配子体(gametophyte)均分化产生出1~3片幼叶(图版Ⅰ:18),配子体的数量开始迅速增加,配子体基部均开始分化产生大量假根。碎米藓的原丝体在配子体发生后,仍然继续分化生长,尤其是绿丝体基部分枝继续增加。东亚碎米藓的绿丝体系统在配子体分化产生之后,依然缓慢生长。30 d时,两种藓类植物的配子体迅速发育,均产生了10~20片幼叶(图版Ⅰ:19,20)。在配子体发生阶段,两种碎米藓属藓类植物均用20 d左右时间在孢子附近的绿丝体上分化产生了配子体原始细胞。两种藓类植物原丝体在配子体分化阶段继续分化生长。
3 讨论与结论
3.1 孢子萌发型与系统发育
3.1.1 两种藓类植物孢子萌发型 孢子萌发型的划分要依据藓类植物稳定的发育性状特征。碎米藓属两种藓类植物孢子均为壁外多极萌发,发育形成半球状细胞组成的绿丝体。碎米藓在绿丝体1,2. 孢子; 3,4. 绿丝体原始细胞; 5,6. 2极萌发的孢子; 7,8. 3极萌发的孢子; 9,10. 绿丝体一级分枝; 11. 绿丝体顶端分化成轴丝体; 12. 半圆球状细胞组成的原丝体系统; 13. 具有斜壁细胞的轴丝体; 14. 高密度生长的原丝体; 15,16. 配子体原始细胞; 17. 假根; 18. 幼小配子体; 19,20. 10~20片幼叶的配子体。比例尺:1-8,15=50 μm; 9-13,16-18=100 μm; 14=200 μm; 19,20=500 μm。1,3,5,7,9,11,13,14,15,17,19. 碎米藓; 2,4,6,8,10,12,16,18,20. 东亚碎米藓。
1,2. Spore; 3,4. Chloronema initial cell; 5,6. Spore appears two pole germination; 7,8. Spore appears third pole germination; 9,10. Chloronema first-order branches; 11. Chloronema apical cells to produce caulonema; 12. Protonema system composed of semicircular globular cells; 13. Caulonema with oblique wall cells; 14. High density growth of protonema; 15,16. Gametophyte initial cell; 17. Rhizoid; 18. Young gametophyte; 19,20. Gametophyte producing 10-20 young leaves. Scale:1-8,15=50 μm; 9-13,16-18=100 μm; 14=200 μm; 19,20=500 μm。1,3,5,7,9,11,13,14,15,17,19. F. pusilla; 2,4,6,8,10,12,16,18,20. F. matsumurae.
頂端分化产生轴丝体,轴丝体细胞直径随着生长发育逐渐变小。两种藓类植物原丝体均产生1~15个细胞组成的短枝,短枝均以绿丝体原始细胞为中心,向四周散射生长,配子体原始细胞均在绿丝体基部细胞上产生。两种藓类植物整个原丝体系统只存在有绿丝体、轴丝体和假根三种形态。该研究参照Nishida(1978)对藓类植物孢子萌发型划分标准,确定了两种碎米藓属植物原丝体发育类型均为蓑藓型(Maromitrium-type)(Nishida,1978;胡人亮,1987)。Nishida(1978)研究认为蓑藓型藓类植物多为孢外萌发,原丝体发育过程中出现由圆柱状或半球形细胞组成的原丝体,原丝体系统只含有绿丝体和次生假根或绿丝体、轴丝体俱存。因此,根据碎米藓属两种藓类植物原丝体发育特点,确定其孢子萌发型为蓑藓型。
3.1.2 两种藓类植物孢子萌发型与其系统发育地位 藓类植物的孢子萌发型与其系统发育地位有着密切关系。多数苔藓分类学家都认为原丝体性状的演化顺序为片状原丝体—块状原丝体—半圆球状原丝体—丝状原丝体。其中,片状原丝体特征是相对原始的,丝状原丝体是原丝体发育形态中的最高级的性状特征(Nishida,1978;Duckett & Ligrone,1994)。在两种碎米藓属植物中,碎米藓原丝体发育过程出现了半圆球状原丝体和丝状轴丝体,东亚碎米藓仅出现了半圆球状原丝体,因此,碎米藓原丝体系统比东亚碎米藓发达。
根据Nishida(1978)对藓类植物原丝体发育特征研究,结合两种碎米藓属植物原丝体发育性状,本研究认为碎米藓较为进化。究其原因,可能因东亚碎米藓在水分相对湿润的南方和干旱少雨的北方树生环境中都有分布,其对外部环境的适应性较强。在实验室优越的培养条件下,两种藓类植物在原丝体发育阶段依然以半圆球状原丝体性状占主体,且原丝体生长发育速度缓慢,说明了发育性状演化的稳定性以及藓类植物的孢子萌发型不受生境条件影响(Nishida,1978;赵建成等,2005)。
3.2 原丝体发育特征与生境相关性
两种碎米藓属植物的原丝体细胞形状为半圆球形,生长缓慢。原丝体的分枝均集中分布在绿丝体原始细胞周围,绿丝体均为1~15个细胞组成,原丝体系统生长半径有限。上述特殊的原丝体发育性状特征,均和藓类植物的生长环境有着紧密的相关性。
本研究所涉及到的两种碎米藓属藓类植物生境均为树生环境。有学者对金华市和信阳地区树附生鲜类植物研究时,均采集到了碎米藓属的藓类植物(卢东升和陈世峰,1995;谢小伟和郭水良,2003),说明该属藓类植物生境以树附生为主。同树下腐殖质及土生环境相比,树干的水分条件更为苛刻。本研究两种碎米藓属植物在原丝体发育阶段,均产生了仅由1~15个半圆球状绿丝体细胞构成的不发达的原丝体分枝系统。前期研究的中华缩叶藓(Ptychomitrium sinense)、短叶扭口藓(Barbula tectorum)和毛口蘚(Trichostomum brachydontium)等藓类植物原丝体发育过程中(李敏等,2006;于淑玲,2008),均出现孢子壁内萌发,形成块状原丝体(massive protonema)。原丝体细胞呈现块状和短圆柱状等特征的藓类植物多生长在较为干旱的生长环境中。它们通过缩短绿丝体长度,减少在原丝体发育时期过度消耗生境中的养分和水分,因此,本研究认为半圆球状原丝体是藓类植物为了适应干旱环境而产生的一种适应性状,该性状随着藓类植物进化,逐步演变成一种稳定的发育学性状特征。树附生藓类植物研究中出现频率较高的藓类植物还有钟帽藓(Venturiella sinensis)、中华蓑藓(Macromitrium cavaleriei)、细叶小羽藓(Haplocladium microphyllum)和阔叶丛本藓(Anoectangium clarum)(谢小伟和郭水良,2003),同两种碎米藓属植物一样,以上四种树附生藓类植物的原丝体发育类型均为蓑藓型,其原丝体发育过程特征与生境均有着密切相关性。多数旱生型藓类植物为了应对干旱环境,会主动调整发育周期,减少原丝体发育阶段时间,产生片状、块状或泡状原丝体,还会采取快速产生配子体和孢子体,完成受精作用,以达到短时间内完成生活史的目的(胡人亮,1987)。
3.3 配子体发生特点与生殖策略
面对高大的被子植物,藓类植物生境选择存在压力,身体矮小的藓类植物为了扩大种群数量,保持了生殖策略的多样性,以适应复杂多变的环境。两种碎米藓属植物在实验条件下,均完成了从孢子萌发到原丝体发育,再到配子体产生的过程。实验室培养基水分充足,两种碎米藓属植物保持在单倍体生长阶段,均用20 d左右时间便分化产生了配子体原始细胞,2种配子体原始细胞分化产生位置有共性特点,即均在孢子附近的绿丝体上分化形成。在实验过程中,均未见到两种藓类植物产生孢子体,这似乎说明在水分充足的条件下,两种藓类植物生殖策略选择了r型生态对策,选择通过无性生殖扩大种群,以避免有性生殖带来的生物能量过度消耗。这种现象在前期丛藓科植物和部分侧蒴藓类植物的孢子萌发和原丝体发育实验中也得到了验证(赵建成等,2005;李敏等,2006;于淑玲,2008)。在野外采集时,两种树生藓类植物均生长有孢子体,其野外生境比实验室内环境更干旱少水,植物体面对相对干旱的环境,多选择有性生殖策略,及时完成生活史,产生孢子体以度过恶劣环境。在实验室内培养时,水分条件充足,却未发现产生孢子体。可以看出,对于同一种藓类植物,在面对不同生境时,可随时调整生殖策略,以保持种群数量的有效扩张。
参考文献:
DUCKETT JG, LIGRONE R, 1994. Studies of protonemal morphogenesis in mosses Ⅲ. The perennial gemmiferous protonema of Rhizomnium punctatum (Hedw.) Kop [J]. J Bryol, 18(1): 13-26.
FAN QS, ZHAO JC, HUANG SL, 2004. Experimental observations on the gametophyte development of Pylaisiella polyantha [J]. J Wuhan Bot Res, 22(2): 140-144. [范庆书,赵建成,黄士良,2004. 金灰藓(Pylaisiella polyantha)配子体发生的实验观察 [J]. 武汉植物学研究,22(2):140-144.]
GAO Q, ZHANG Y, 1986. A preliminary study on spore germination and protonema development of the moss in China [J]. J Wuhan Bot Res, 4(2): 123-132. [高谦,张钺,1986. 中国藓类植物孢子萌发和原丝体发育的初步研究 [J]. 武汉植物研究,4(2):123-132.]
HU RL, 1987. Bryophyte botany [M]. Beijing: Higher Education Press: 144-159. [胡人亮, 1987. 苔藓植物学 [M]. 北京:高等教育出版社:144-159.]
HUANG SL, WANG XR, JIN HX, et al., 2015. Studies on the characteristics of spore germination and protonema development of eighteen species in family Pottiaceae [J]. J Hebei Norm Univ, 1(39): 71-78. [黄士良,王晓蕊,金红霞,等,2015. 丛藓科18种藓类植物孢子萌发与原丝体发育特征研究 [J]. 河北师范大学学报,1(39):71-78.]
HUANG SL, 2009. Study on the spore germination and protonema development of pleurocarpous mosses [D]. Shijiazhuang: Hebei Normal University. [黄士良,2009. 侧蒴藓类植物孢子萌发与原丝体发育研究 [D]. 石家庄:河北师范大学.]
JIA Y, HE S, 2013. Species catalogue of China(Vol. 1) [M]. Beijing: Science Press. [贾渝,何思,2013. 中国生物物种名录 (第一卷) [M]. 北京:科学出版社.]
KATSUMATA T, FUKAZAWA J, MAGOME H, et al., 2011. Involvement of the CYP78A subfamily of cytochrome P450 monooxygenases in protonema growth and gametophore formation in the moss Physcomitrella patens [J]. Biosci Biotechnol Biochem, 11(9):311-316.
LI M, ZHAO JC, HUANG SL, et al., 2006. Characteristics of spore germination and protonemal development in Ptychomitrium sinense [J]. Acta Bot Boreal-Occident Sin, 26(8): 1521-1525. [李敏,趙建成,黄士良,等,2006. 中华缩叶藓孢子萌发与原丝体发育特征研究 [J]. 西北植物学报,26(8):1521-1525.]
LIU BD, CONG YZ, 2003. Studies on spores culture and developmental biology of Atrichum undulatum [J]. Bull Bot Res, 23(2):159-163. [刘保东, 丛迎芝,2003. 波叶仙鹤鲜的孢子培养及发育生物学研究 [J]. 植物研究,23(2):159-163.]
LU DS, CHEN SF, 1995. A study of the musei on trees from Xinyang [J]. J Xinyang Technol Coll(Nat Sci Ed), 4(8): 425-428. [卢东升,陈世峰,1995. 信阳地区树生鲜类植物研究 [J]. 信阳师范学院学报(自然科学版),4(8):425-428.]
NISHIDA Y, 1978. Studies on the sporeling types in mosses [J]. J Hatt Bot Lab, 44: 371-454.
PRESSEL S, MATCHAM HW, DUCKETT JG, et al., 2005. Studies of protonemal morphogenesis in mosses. X. Ephemeraceae revisited: new dimensions underground [J]. Transact British Bryol Soc, 27: 311-318.
WU PC, JIA Y, 2004. Flora Bryophytorum Sinicorum(Vol. 8) [M]. Beijing: Science Press: 88-315. [吴鹏程,贾渝,2004. 中国苔藓志(第八卷) [M]. 北京:科学出版社:88-315.]
XIE XW, GUO SL, 2003. Studies on ecology of epiphytic moss species in Jinhua suburb [J]. Bull Sci Technol, 19(5): 407-412. [谢小伟,郭水良,2003. 金华市郊树附生藓类植物生态学研究 [J]. 科技通报,19(5):407-412.]
YU SL, 2008. Study on the characteristics of protonema development in Barbula tectorum C. Muel [J]. J Huazhong Norm Univ (Nat Sci Ed), 42(3): 440-443. [于淑玲,2008. 短叶扭口藓原丝体发育特征的实验研究 [J]. 华中师范大学学报(自然科学版),42(3):440-443.]
ZHAO JC, LI XQ, ZHANG HZ, 2002. A preliminary study on spore germination and protonema development of ten species of mosses [J]. Arid Zone Res, 19(1): 32-38. [赵建成,李秀芹,张慧中,2002. 十种藓类植物孢子萌发与原丝体发育的初步研究 [J]. 干旱区研究,19(1):32-38.]
ZHAO JC, HUANG SL, LI M, et al., 2005. Experimental studies on the characteristics of gemma development in Barbula indica [J]. Bu Bot Res, 25(2): 169-172. [赵建成,黄士良,李敏,等,2005. 小扭口藓(Barbula indica)芽胞发育特征的实验研究 [J]. 植物研究,25(2):169-172.]
(责任编辑 周翠鸣)
