
利用DNA片段测序方法探究枳属和富民枳的分类地位
2021-03-24 18:30张余龚洵冯秀彦
广西植物 2021年1期
张余 龚洵 冯秀彦
摘 要: 富民枳(Poncirus polyandra)屬于芸香科(Rutaceae)枳属(Poncirus Raf.)。自发表以来,分类地位一直备受争议,其中在Flora of China中认为富民枳为柑橘杂交种(Poncirus polyandra),把枳属归并于柑橘属(Citrus)。该研究选取枳属的富民枳、枳(Poncirus trifoliata)及柑橘属下8个种共10个种47个个体作为研究材料,以九里香(Murraya exotica)为外类群,利用3个叶绿体片段(trnL-trnF、trnS-trnG、rbcL)、ITS片段和1个单拷贝核基因(Chr5)数据构建系统发育树,探究枳属和富民枳的分类地位。结果表明:基于3个叶绿体片段数据构建的最大似然树(ML)和贝叶斯树(BI)的拓扑结构基本一致, 10个物种聚为两大分支,即柑橘属的8个物种聚为一大分支,富民枳和枳聚为另一大分支。其中,富民枳所有个体聚为一小单系分支,枳的所有个体聚为一小单系分支,支持枳属和富民枳独立存在。2个核DNA片段数据结果显示,枳属的两个种与柑橘属的8个物种聚在一个大分支里,无法确立枳属的单系地位,但富民枳的9个个体聚在一起,暗示富民枳在遗传上是一个独立的类群。综上研究认为,无论是叶绿体DNA数据还是核DNA数据均支持富民枳是一个独立的物种,但核DNA数据不支持枳属成立。
关键词: 枳属, 富民枳, 柑橘属, 系统发育, 分类地位
Abstract: Poncirus polyandra belongs to the genus Poncirus Raf. of Rutaceae. Since it was published, its taxonomic status has always been controversial. In Flora of China, P. polyandra was regarded as a Citrus hybrid (Poncirus polyandra), and the genus Poncirus was included in the genus Citrus. In this study, we used three chloroplast DNA fragments (trnL-trnF, trnS-trnG, rbcL), ITS and one single copy nuclear genes (Chr5) to construct phylogenetic trees to explore the taxonomic status of the genus Poncirus and P. polyandra. We sampled a total of 47 individuals from ten species, including P. polyandra and P. trifoliata of the genus Poncirus as well as eight other species of the genus Citrus by using Murraya exotica was used as the outgroup. The results were as follows: The maximum likelihood trees (ML) and Bayesian inference trees (BI) constructed by all data were basically identical in topological structure. The cpDNA results showed that all species were clustered into two clades: eight species of Citrus were clustered into one major clade, and P. polyandra and P. trifoliata were clustered into another major clade. Meanwhile, all individuals of P. polyandra and P. trifoliata were grouped into a monophyletic subclade respectively, supporting the independence of Poncirus and P. polyandra could exist independently; However, the results of two nuclear DNA fragments showed that two species of Poncirus and the eight species of Citrus clustered into a large clade, indicating that the genus Poncirus cannot be recognized independently. Moreover, nine individuals of P. polyandra were monophyletic, suggesting that P. polyandra was a genetically independent lineage. In summary, both chloroplast DNA data and nuclear DNA data supported that P. polyandra was an independent taxa, but nuclear DNA data from our study did not support the independence of Poncirus.
Key words: Poncirus Raf, Poncirus polyandra, Citrus, phylogeny, phylogenetic position
枳属(Poncirus Raf.),又名枸橘属,属下植物有枳(Poncirus trifoliata)和富民枳(P. polyandra)两个种(方德秋,1993),原产于我国,是芸香科(Rutaceae)柑橘亚科下的一属,区别于柑橘、金柑等属,主要分布于长江中游两岸各省及淮河流域一带,分布范围较柑橘属更广。该属在我国传统医药上使用很早,早在李时珍《本草纲目》中有记载。近代中医学研究发现,其药用作用主要是因为存在一种枳苷的物质(胡源祥等,2017a, b)。在生产上,枳属植物是一种优秀的柑橘矮化砧,抗逆性强。枳属的划分一直与枳的系统分类位置有密切关系,枳属的研究曾历经三个阶段:(1)枳属的不认可阶段,在公元1178年韩彦直《橘录》被称作枸橘,1763年分类学家林奈把枳归类于柑橘属;(2)枳属认可阶段,认为枳属区别于柑橘属(Tomimatsu, 1968;Tomimatsu & Hashimoto, 1968);(3)枳属的分类存在争议,2008年Flora of China修订版中把枳属归类到柑橘属(Zhang, 2008),但有些学者认为枳属区别于柑橘属,并且是一种优秀的种质资源(Heo et al.2018;George & Lapointe, 2019)。
富民枳属于芸香科(Rutaceae)枳属,是丁素琴等(1984)发表的一个新种,它是国家二级保护植物,分布于云南富民县的特有种。富民枳是常绿小乔木,区别于枳的形态特征(武晓晓等,2018),分类地位备受争议,作为枳属的一个新种得到了叶片同工酶证据(Fang, 1993)、 SSR 标记的高纯合率(Pang et al., 2003)及核型分析(刘利勤等,2007)的支持,但郭天池等(1996)认为富民枳是一个天然枳橙类型。在当地富民枳常作为药用植物使用,是一种珍贵的药用资源,可以抑制新生血管形成,降低肿瘤细胞表达(胡源详等,2017c)。由于人为因素的影响、原生地的过度放牧以及作为常绿观赏植物的大量挖掘,富民枳已经野外灭绝(邢定生,2008),于2009年被列入云南极小种群拯救计划五个物种之一。但是富民枳的分类地位仍然没有得到解决,2008年Flora of China修订版中认为富民枳是柑橘属的一个杂交种(Poncirus × polyandra)(Zhang, 2008),而张珊珊等(2018)把富民枳作为一个独立的种并进行了遗传多样性的研究。
在形态界定物种存在争议的情况下,可借助分子手段解决。目前,已发展起一系列的DNA分子标记,如SSR (Simple Sequence Repeats,简单重复序列)、ISSR(Inter Simple Sequence Repeat,简单重复序列间多态性)、SRAP (Sequence-Related Amplified Polymorphism,序列相关扩增多态性)等被用来研究富民枳与其近缘属植物间的亲缘关系(庞小明等,2003;张连峰,2006;刘通,2014)。然而,DNA分子片段直接测序技术在国内很多植物研究上均有报道(李映志, 2006),常见的有叶绿体DNA (cpDNA)片段(李小孟,2010)和核基因(nDNA)。因为旁系同源的存在,核基因報道较少,但nDNA作为植物遗传物质最重要的载体,且具双亲遗传特性方面优点,是一种理想的DNA分子标记,常用于种属分类关系的探究(成彩霞等,2018)。本研究利用母系遗传的cpDNA和双亲遗传的nDNA直接测序的方法,以期探究枳属和富民枳的分类地位。
1 材料与方法
1.1 材料
本研究共采集枳属和柑橘属植物的47份分子材料,其中包括9份栽培于中国科学院昆明植物研究所的富民枳新鲜植物叶片材料,这9株富民枳于1992年从富民枳的原生地富民县大营镇老青山冬瓜岭引种(102.49465 E,25.22583 N,海拔2 180 m);8份枳的新鲜植物叶片材料,其中2份来自中国科学院昆明植物研究所;3份红河大翼橙低温冷藏植物叶片材料(中国科学院昆明植物研究所资源室龚洵课题组);6份枳橙的新鲜植物叶片材料;4份柚新鲜植物叶片材料;6份柑橘新鲜植物叶片材料;3份香橼新鲜植物叶片材料;3份金柑新鲜植物叶片材料;2份箭叶橙新鲜植物叶片材料和3份宜昌橙新鲜植物叶片材料。另外采集1份九里香(Murraya exotica)新鲜植物叶片材料作为本研究的外类群。采集的新鲜叶片立即放入硅胶干燥,将干燥好的叶片在-20 ℃冰箱保存备用。植物材料采集信息见表1。
1.2 方法
采用改良的CTAB法从植物材料中提取总DNA (Doyle, 1991)。通过查阅文献,从常用于植物系统学、群体遗传学和保护生物学等的DNA分子片段中进行筛选,共筛选到3个cpDNA片段和2个nDNA片段(唐政, 2014; 杨晓明, 2017),其引物信息见表2。对于目的条带清晰,特异性好的PCR扩增产物,送昆明硕擎生物有限公司进行双向测序以保证测序的准确性,测序使用与扩增相同的引物,测序序列已上传到NCBI,接受号为MN585512-MN585655和MN638942-MN639037。
1.3 数据分析
DNA分子片段经过测序后,利用SEQMAN (Swindell & Plasterer, 1997)软件进行序列的编辑和拼接。然后将拼接好的序列用BioEdit ver. 7.0.4.1 (Hall, 1999)进行序列的比对和手工校对。对于3个叶绿体DNA分子片段,利用PAUP ver. 4.0b10 (Swofford, 2003)软件进行序列联合并进行同质性检验,若P>0.05则说明片段的一致性好,可以用于联合分析。因为2个核基因(ITS和chr5)外显子片段都存在双峰的个体,且一致性检验P<0.05,因此,未将核基因进行联合分析。
以芸香科九里香做为外类群,采用最大似然法(maximum likelihood,ML)和贝叶斯法(Bayesian inference,BI)构建系统发育树以进行枳属和富民枳的分类地位研究。ML分析所需模型及树的构建均在软件MEGA ver. 5 (Tamra et al., 2011)中执行,树的构建选用Nearest-Neighbor-Interchange (NNI) 搜索方法, 支持率的估计采用1 000次重复分支上方的数字代表靴带值(A)和后验概率(B)。下同。
抽样的靴带分析。BI分析所需最佳碱基替代模型在软件Jmodeltest ver. 2.3.1 (Darriba et al., 2012)中计算,BI分析在软件MrBayes ver. 3.1.2 (Ronquist & Huelsenbeck, 2003)中进行,每一个马尔可夫链从一个随机的树开始,运行106个循环,每100次循环取样一次,支持率选用后验概率(Posterior Probability, PP),利用软件Figtree读取最终的树文件。
2 结果与分析
2.1 叶绿体联合片段分析
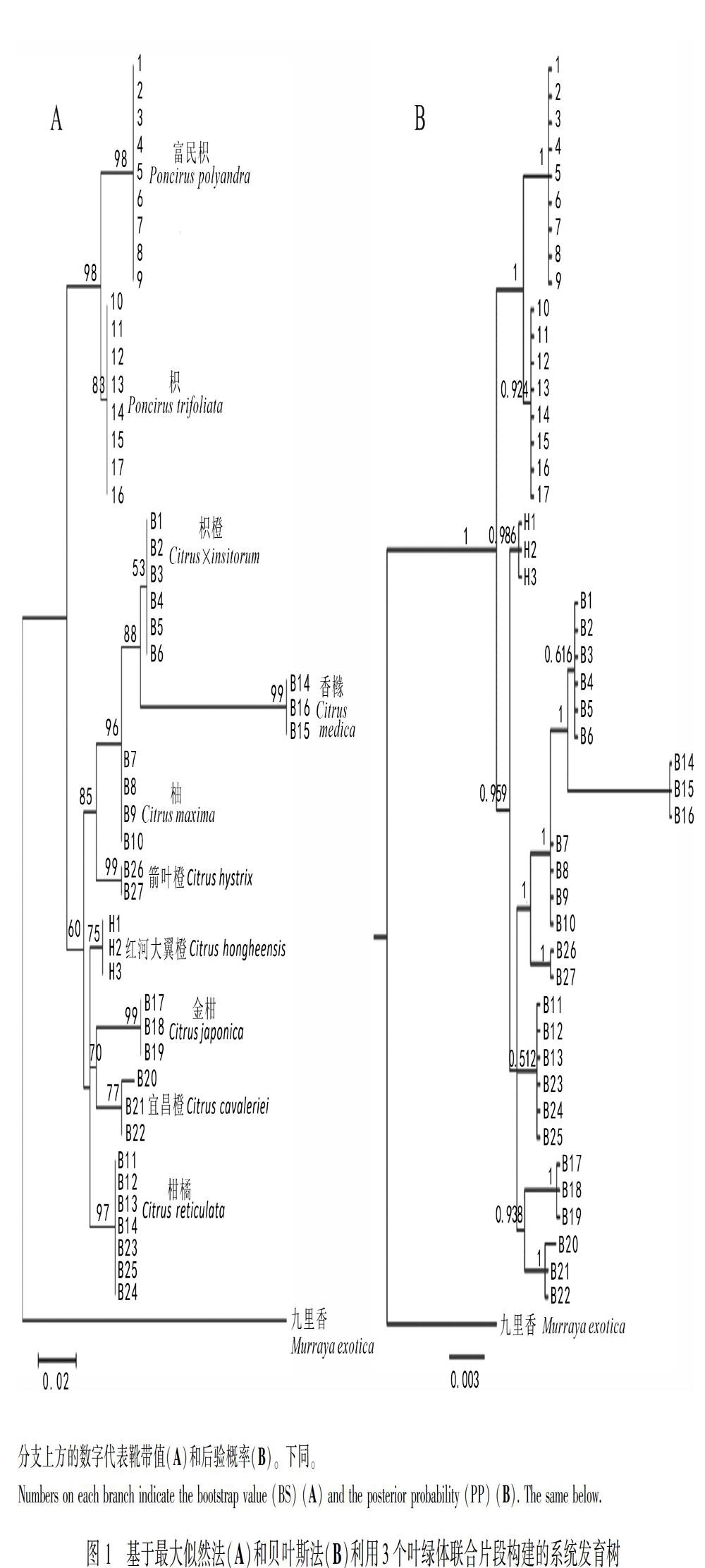
所研究的叶绿体片段长度在类群间和个体间均一致,trnL-trnF的长度为948 bp,共检测到16个多态位点,包括11個位点变异和5个插入/缺失;trnS-trnG的长度为806 bp,共检测到44个多态位点,包括40个位点变异和4个插入/缺失;rbcL的长度为1 266 bp,共检测到18个多态位点,包括9个位点变异和9个插入/缺失。将3个片段进行同质性检验并联合,一致长度为3 020 bp。
基于最大似然法和贝叶斯法构建系统发育树如图1所示,构建最大似然树选用最佳模型为T92模型,贝叶斯分析选用TPMluf-Nei模型,最大似然树和贝叶斯树分支上方的数字分别代表靴带值(Bootstrap value, BS)和后验概率(PP)。两种推导法构建系统树的拓扑结构比较一致,内类群的所有物种聚成两大支且得到了较高的支持率,其中柑橘属的8个物种聚为一大分支(BS=60; PP=0.959),枳属的富民枳和枳聚为另一大分支(BS=98; PP=1),枳属与柑橘属的类群并没有交叉,二者均是单系。其中在枳属的这一分支里,富民枳的所有个体聚为一小单系分支,枳的所有个体聚为另一小单系分支,均得到了高支持率(BB=98, 83; PP=1, 0.924)。在柑橘属的分支里,每个物种的所有个体都聚为一个小的单系分支,并不存在交叉。但两种方法在对柑橘属物种的系统发育关系的处理上存在稍许差别,如红河大翼橙的系统位置不同,最大似然树显示红河大翼橙位于分支内部位置,而贝叶斯树显示红河大翼橙最早分化出来。两种树均显示枳橙和香橼聚为一支,随后又与柚聚在一起,说明它们的亲缘关系较近;金柑和宜昌橙聚在一支,说明二者具有较近的亲缘关系。
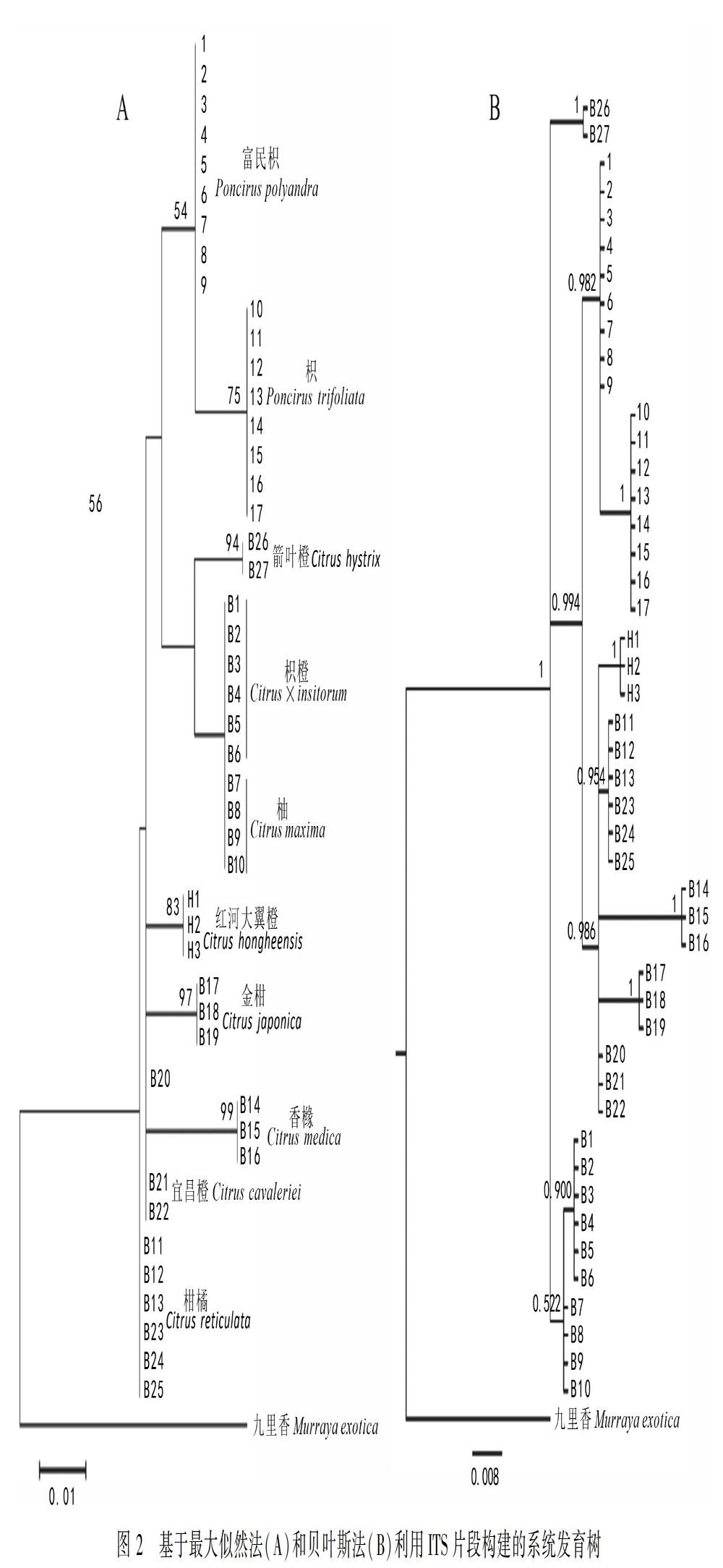
2.2 ITS片段分析
ITS片段经过整理剪切的一致长度为756 bp,共检测到66个多态位点,包括63个位点变异和3个插入/缺失。构建最大似然树选取HKY+G+I为最佳模型,贝叶斯分析选用TIM3+G模型。基于ITS联合片段构建的最大似然树和贝叶斯树见图2,ML树和BI树的拓扑结构并不一致,但两种树的结果均显示富民枳和枳聚为一支(BS=54;PP=0.982),嵌套在柑橘属的分支里,但富民枳和枳与柑橘属的其他物种并没有交叉。ML树所得到的聚类关系支持率均较低,结果显示柑橘最先分化出来,然后是红河大翼橙、金柑、香橼和宜昌橙,而四者呈并系关系(BS<50);柚和枳橙聚为一支,然后二者又与箭叶橙聚在一起(BS=56)。BI树所得到的聚类关系支持率均较高,结果显示所有物种聚为三大支(PP=1),箭叶橙为一分支;富民枳、枳、红河大翼橙、柑橘、香橼、金柑和宜昌橙聚为一大分支(PP=0.994),其中富民枳与枳聚为一小单系分支(PP=0.982),另外5个柑橘属植物聚为另一小单系分支,但它们呈并系关系(PP=0.986);枳橙与柚聚为另一大分支(PP=0.522)。
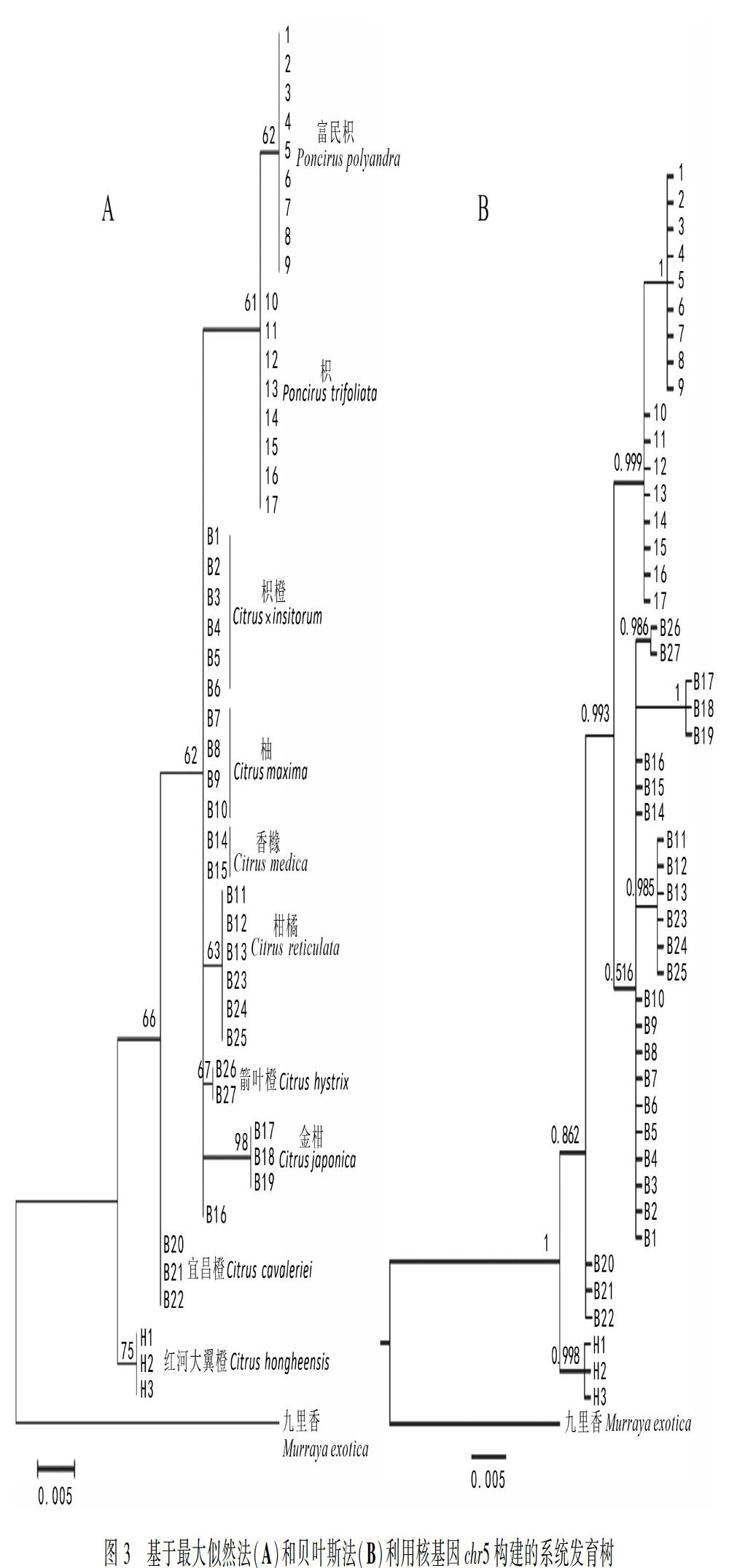
2.3 单拷贝核基因Chr5分析
Chr5片段比对剪切后一致性长度为780 bp,共检测到39个多态位点,全为位点变异,构建的系统发育树见图3,其中ML树构建选择TM93+I模型,BI树的构建模型为TIM1。基于Chr5片段构建的ML树和BI树的拓扑结构是一致的,结果见图3。与ITS数据结果相似,富民枳和枳聚为一支嵌套在柑橘属植物的分支里,但二者与柑橘属的其他物种并没有交叉,ML树的支持率较低,BI树的支持率较高。系统树结果显示红河大翼橙最先分化出来(BS<50;PP=1),然后是宜昌橙(BS=65;PP=0.862),随后枳橙、柚、香橼、柑橘、箭叶橙和金柑聚为一支(BS=62;PP=0.516),最后与富民枳和枳的分支聚为一大分支(BS=62;PP=0.993)。
3 讨论与结论
3.1 枳属的分类地位
本研究以“枳属”单独存在为基础,选择枳属的两个种和柑橘属的8个种为研究对象,利用3个cpDNA片段、ITS和1个核基因对枳属的系统发育进行研究。cpDNA的数据结果均一致支持枳属为一单系,与柑橘属的物种并无交叉,说明两属之间无基因交流。本研究结果和武晓晓等(2018)、杨晓明(2017)及Wu et al.(2018)的研究结果一致,枳属能够作为一个独立的属,而不应将其归并到柑橘属中。但3个叶绿体联合片段的ML树和BI树显示柑橘属物种的系统发育关系是不一致的,如红河大翼橙的系统位置不同,ML树显示红河大翼橙位于分支内部位置,而BI树显示红河大翼橙是最早分化出来的物种。这可能是由于3个叶绿体DNA分子片段信息位点不足,基于不同的方法推断出来的结果存在冲突。
与cpDNA的研究结果不同的是ITS和Chr5的数据结果显示枳属并不能与柑橘属分开,而是嵌套在柑橘属的分支里,说明两个核DNA的数据并不支持枳属能够单独成属,而应归并在柑橘属里,这与2008年Flora of China修订版(Zhang, 2008)中对枳属的处理是一致的,但与Wu et al.(2018)基于基因组数据的研究结果不一致。另外,这2个核DNA数据显示柑橘属8个物种的关系也不一致,比如红河大翼橙,在ITS构建的系统树上处于内部分支里,而在Chr5构建的系统树上显示最早分化出来。不同核DNA分子片段数据揭示的结果不一致,可能是由于核基因的不完全谱系分选造成的,亦或它们受到的选择压力和进化速率不一致造成的。另外,在核基因中往往存在重组现象,可能对物种系统发育关系也会存在影响。
本研究中,不同类型的DNA分子片段数据在对枳属和柑橘属物种的系统发育关系解决上存在差异,cpDNA数据支持枳属能够独立成属,而核DNA数据显示枳属嵌套在柑橘属里不能单独成属。因为质体基因是母系遗传的,而核基因是双亲遗传的,质体基因所拥有的Ne是核基因的1/4,Ne可以影响一个物种单倍型成为单系的速度(Moore, 1995)。因此,基于1个质体基因物种的单倍型成为单系所需的时间是基于1个核基因所需时间的1/4,即质体基因能更快地体现出一个物种的单倍型成为单系。可能在解决属的分类地位时,叶绿体数据也能尽早体现出来。所以在本研究中,叶绿体数据能够解决枳属的分类地位,而两个核DNA则不行。另外,根据Moore的观点,在1个质体基因能95%成功地解决一个物种是单系时,就需要用16或者40个核基因才能达到如此成功率。所以,想要解决一个属或者物种的分类地位就需要采用更多的核基因,这也可能造成本研究中较少的核基因不能解决枳属的分类地位。另外,叶绿体片段数据与核基因数据结果存在差异,也可能暗示枳属杂交起源,这需要采用更多的核基因数据或从组学水平上进行深入探究。
3.2 富民枳的分类地位
序列位点差异直观反映各个研究材料之间基因层次上的差异性,这种研究方式是直觀有效的,这种方法在很多植物领域都有利用。本研究通过具有母系遗传特性的3个叶绿体片段和具有双亲遗传特性的核基因片段ITS和Chr5数据来研究富民枳的分类地位以及它与枳、红河大翼橙、枳橙、柚、柑橘、香橼、金柑、箭叶橙这些种的系统发育关系。基于ML法和BI法构建系统发育树,所有数据结果均显示富民枳的9个个体聚在一起,并未与其他任何物种有交叉,支持富民枳是一个遗传上相对独立的类群,而不支持Flora of China将其作为柑橘属的一个杂交种的分类处理。因此,富民枳不是一个柑橘杂交种,而是为一个独立“好种”,且与前人研究结果一致(吴兴恩等, 2006;龚桂芝等, 2008)。本研究结果表明,虽然核DNA数据并不支持枳属的成立,但是,无论叶绿体DNA数据还是核DNA数据均支持富民枳是一个独立的物种,是一种优质的种质资源(张珊珊等,2018),具有重要的保护价值。
参考文献:
CHENG CX, SU X, GAO T, et al., 2018. nrDNA ITS sequence analysis of Grayia spinosa (Chenopodiaceae) from different regions in the Western United States[J]. Guihaia, 38(5): 617-625. [成彩霞, 苏雪, 高婷, 等, 2018. 美国西部不同地区Grayia spinosa核DNA ITS序列分析[J]. 广西植物, 38(5):617-625.]
DARRIBA D, TABOADA GL, DOALLO R, et al., 2012. Model test 2: more models, new heuristics and parallel computing[J]. Nat Methods, 9(8): 772-772.
DING SQ, ZHANG XN, BAO ZR, et al., 1984. A new species of Poncirus from China [J]. Acta Bot Yunnan, 6(3): 292-293. [丁素琴, 张显努, 暴卓然, 等, 1984. 中国枳属一新种[J]. 云南植物研究, 6(3):292-293.]
DOYLE J, 1991. DNA protocols for plants-CTAB total DNA isolation [M]//HEWITT GM, JOHNSON A (eds.). Molecular techniques in taxonomy. Berlin: Springer: 101-115.
FANG DQ, 1993. Intra and intergeneric relationships of Poncirus polyandra: investigation by leaf isozymes[J]. J Wuhan Bot Res, 11(1): 34-40. [方德秋, 1993. 富民枳的分类地位及其在进化中的作用研究——叶片同工酶证据[J]. 武汉植物学研究, 11(1):34-40.]
GEORGE J, LAPOINTE SL, 2019. Host-plant resistance associated with Poncirus trifoliata influence oviposition, development and adult emergence of Diaphorina citri (Hemiptera: Liviidae) [J]. Pest Mang Sci, 75(1): 279-285.
GONG GZ, HONG QB, PENG ZC, et al., 2008. Genetic diversity of Poncirus and its phylogenetic relationships with relatives as revealed by nuclear and chloroplast SSR[J]. Acta Horti Sin, 35(12): 1742-1750. [龚桂芝, 洪棋斌, 彭祝春, 等, 2008. 枳属种质遗传多样性及其与近缘属植物亲缘关系的SSR和cpSSR分析[J]. 园艺学报, 35(12): 1742-1750.]
GUO TC, CHEN QY, YE YM, 1996. Poncirus trifoliata germplasm resources[M]. South China Fruit, (3):8-10. [郭天池, 陈全友, 叶荫民, 1996. 枳的种质资源[M]. 中国南方果树, (3):8-10.]
HALL TA, 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[J]. Nucl Acids Symposium Ser, 41(41): 95-98.
HEO Y, CHO Y, JU KS, et al., 2018. Antiviral activity of Poncirus trifoliata seed extract against oseltamivir-resistant influenza virus[J]. J Microbiol, 56(8): 586-592.
HU YX, SONG YP, TAN SS, et al., 2017a. Extraction process of Poncirin in Fructus Ponciri trifoliate[J]. J Jinggangshan Univ (Nat Sci Ed), 38(2): 83-86. [胡源祥, 宋玉鹏, 谭舒舒, 等, 2017a.绿衣枳壳中枳属苷的提取工艺研究[J].井冈山大学学报(自然科学版), 38(2): 83-86.]
HU YX, CHEN HF, SONG YP, et al., 2017b. Determination of flavonoids in Fructus Ponciri trifoliatae and Fructus Aurantii by HPLC[J]. J Pharm Anal, 37(12): 2162-2166. [胡源祥, 陈海芳, 宋玉鹏, 等, 2017b. HPLC法测定酸橙枳壳和枸橘枳壳中黄酮苷的含量[J]. 药物分析杂志, 37(12):2162-2166.]
HU YX, ZHU HQ, TAN SS, et al., 2017c. Isolation and purification of Poncirin form Fructus Ponciri trifoliatae by multi-solvent partition[J]. J Jinggangshan Univ (Nat Sci Ed), 38(5): 85-88. [胡源祥, 朱海强, 谭舒舒, 等, 2017c. 多溶剂萃取法分离制备绿衣枳壳中枳属苷[J]. 井冈山大学学报(自然科学版), 38(5):85-88.]
LI YZ, 2006. Genetic diversity and phylogeny of Citrus reticulate Blanco [D]. Wuhan: Huazhong Agricultural University. [李映志, 2006. 宽皮柑橘(Citrus reticulate Blanco)的遗传多样性及系统发育研究[D]. 武汉:华中农业大学.]
LI XM, 2010. Molecular phylogeny of the true citrus fruit trees group (Aurantioideae, Rutaceae) and the origin of cultivated Citrus[D]. Chongqing: Southwestern University. [李小孟, 2010. 柑橘及其近缘属植物的分子进化与栽培柑橘的起源研究[D]. 重庆:西南大学.]
LIU LQ, YANG J, GU ZJ, 2007. Karyomorphology and taxonomic position of Poncirus polyandra (Rutaceae)[J]. Acta Bot Yunnan, 29 (2):198 -200. [ 刘利勤, 杨静, 顾志建, 2007. 富民枳的核型与分类位置探讨[J]. 云南植物研究, 29(2):198-200.]
LIU T, 2014. Genetic diversity analysis of Guangxi Citrus germplasms using SSR and SPAP[D]. Wuhan: Huazhong Agricultural University. [刘通, 2014. 应用SSR和SRAP技术研究广西柑橘资源遗传多态性[D]. 武汉:华中农业大学.]
MOORE WS, 1995. Inferring phylogenies from mtDNA variation: mitochondrial-gene trees versus nuclear-gene trees[J]. Evolution, 49(4): 718-726.
PANG XM, HU CG, DENG X, 2003.Phylogenetic relationships mmong Citrus and its relatives as revealed by SSR markers[J]. J Genet, 30(1): 81-87. [龐晓明, 胡春根, 邓秀, 2003. 用SSR标记研究柑橘属及其近缘属植物的亲缘关系[J]. 遗传学报, 30(1):81-87.]
RONQUIST F, HUELSENBECK JP, 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models[J]. Bioinformatics, 19(12): 1572-1574.
SWINDELL SR, PLASTERER TN, 1997. SEQMAN [M]//SWINDELL SR (ed.). Sequence data analysis guidebook. Vol. 70. New York: Humana Press: 75-89.
SWOFFORD DL, 2003. PAUP: phylogenetic analysis using parsimony, Version 4.0 b10 [J]. Sunderland: Sinauer Associates: 33-39.
TAMURA K, PETERSON D, PETERSON N, et al., 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Mol Biol Evol, 28(10): 2731-2739.
TANG Z, 2014. The polymorphism of Citrus and its relatives based on single-copy nuclear gene (SCNGs) DNA sequences[D]. Wuhan: Huazhong Agricultural University. [唐政, 2014. 基于單拷贝核基因(SCNGs)DNA序列的柑橘多样性研究[D]. 武汉:华中农业大学.]
TOMMIMATSU T, 1968. Studies on the chemical components of Rutaceae plants. I. Components of the root of Poncirus trifoliata Rafinesque (1)[J]. Yakugaku Zasshi-J Pharm Soc J, 88(5): 643-645.
TOMMIMATSU T, HASHIMOTO M, 1968. Studies on the chemical components of Rutaceae plants. II. Components of the root of Poncirus trifoliata Rafinesque (2)[J]. Yakugaku Zasshi-J Pharm Soc J, 88(10).
WU GA, TEROL J, IBANEZ V, et al.,
2018. Genomics of the origin and evolution of Citrus[J]. Nature, 554(7692): 311.
WU XE, FAN MT, GONG X, et al. 2006. The ISSR analysis of 22 Citrus resources[J]. J Yunnan Agric Univ, (1): 36-41. [吴兴恩, 范眸天, 龚洵, 等, 2006. 22份柑桔资源的ISSR分析[J]. 云南农业大学学报, (1):36-41.]
WU XX, TANG Y, DENG CL, et al., 2018. Observation of citrus pollen morphology by scanning electron microscopy[J]. J Fruit Sci, 35(7): 794-801. [武晓晓, 唐艳, 邓崇岭, 等, 2018. 柑橘不同种属花粉形态观察[J]. 果树学报, 35(7):794-801.]
XING DS, 2008-10-21. Carry out investigation on wild plants and alien invasive organisms [N]. Yuxi Daily, (005). [邢定生, 2008-10-21. 开展野生植物及外来入侵生物调查[N]. 玉溪日报, (005).]
YANG XM, 2017.The phylogenomic of Aurantioideae and phylogeography of wild citron(Citrus medic) and ichang papeda (Citrus ichangensis)[D]. Wuhan: Huazhong Agricultural University. [杨晓明, 2017. 柑橘亚科植物系统发育基因组学及野生枸橼、宜昌橙谱系地理学研究[D]. 武汉:华中农业大学.]
ZHANG DX, 2008. Flora of China [M]. Beijing: Beijing Science Press, 11(7): 90-96.
ZHANG LF, 2006. Phylogenetic relationships among Fortunella Swingle and its relatives as revealed by ISSR and SSR markers [D]. Chongqing: Southwestern University. [张连峰, 2006. 金柑属( Fortunella Swingle)及其近缘属植物的ISSR及SSR分析[D]. 重庆:西南大学.]
ZHANG SS, GAN YH, YANG WZ, et al., 2018.Genetic diversity of Poncirus polyandra resources[J]. J West Chin For Sci, 47(4): 14-20. [张珊珊, 甘云浩, 杨文忠, 等, 2018. 富民枳种质资源的SSR遗传多样性研究[J]. 西部林业科学, 47(4):14-20.]
(责任编辑 何永艳)
