
低温逆境胁迫下小麦ROS 代谢及调控机制研究进展
2021-03-19 07:01柯媛媛倪芊芊张乐乐刘绿洲魏凤珍李金才
大麦与谷类科学 2021年1期
柯媛媛,陈 翔,倪芊芊,张乐乐,刘绿洲,许 辉,魏凤珍,李金才,2*
(1.安徽农业大学农学院/农业部华东地区作物栽培科学观测站,安徽合肥230036;2.江苏省现代作物生产协同创新中心,江苏南京210095)
小麦(TriticumaestivumL.)是世界三大粮食作物之一,其籽粒营养丰富。全球约60%的人口以小麦为主食,其稳产、丰产对保障全球粮食安全具有重要意义[1]。随着全球人口大幅度增长,预计到2050 年,粮食作物产量需增加60%才能满足全球人口的粮食需求,而小麦产量年均增长率要达到2.4%[2-3]。工业革命以来,全球气候变暖已是不争的事实,联合国政府间气候变化专门委员会(IPCC)气候变化与土地特别报告(Special Report on Climate Change and Land,SRCCL)指出,2006—2015 年全球陆地平均气温相较于1850—1900 年约升高了1.53℃,预计到2100年将再上升1.5℃以上[4]。而气候变化导致极端低温事件出现的频率、强度和持续时间不断增加[5]。低温逆境主要包括零上冷害和零下冻害,其中倒春寒对黄淮麦区的小麦生产造成的危害较大。据报道,我国黄淮麦区河南省和山东省倒春寒一般发生频率达30%以上,最高年份甚至达到70%[6]。因此,低温逆境已成为限制小麦生产的主要非生物胁迫因素,影响着小麦的丰产稳产与优质。
植物会产生一系列生理生态变化响应低温逆境,一般认为由逆境胁迫引起的活性氧(reactive oxygen species,ROS)积累进而导致的氧化胁迫是影响植物正常生长发育的主要原因[7]。ROS 是植物响应非生物胁迫网络中的重要组成部分,它是化学性质活泼、氧化能力极强的氧代谢物及其衍生的含氧物质的总称,主要包括超氧阴离子(O2·)、过氧化氢(H2O2)、羟自由基(·OH)和单线态氧(1O2)等[8]。由低温引起的ROS 在小麦不同器官的积累均会影响小麦的生长发育。研究发现,过量ROS 通过影响叶片的光合作用和呼吸作用降低“源”器官的生产,同时导致“库”器官小麦穗部花药的氧化损伤,进而败育影响结实。低温引起的ROS 积累还会导致小麦根系细胞膜受损、膜质产生脱脂化及磷脂游离,阻碍根系养分吸收,从而导致减产。因此,明确低温胁迫下小麦体内ROS 代谢机制,对保障小麦生产的高产和稳产具有重要意义。本文系统阐述了ROS 的产生、功能与清除,从小麦叶片生理、穗部发育与根系生理等方面总结了低温胁迫下ROS 对小麦生长发育的影响,并对未来的研究进行展望,以期为我国小麦生产的防灾减灾与提质增效提供理论参考。
1 低温胁迫下活性氧作用机制
1.1 ROS 的产生
逆境胁迫下ROS 在质膜、细胞壁、线粒体、叶绿体和过氧化物酶体等各种亚细胞室中均可产生(图1)[9],其中叶绿体、线粒体等具有高度氧化代谢活性的细胞器更是产生ROS 的主要来源[10]。叶绿体类囊体中PSⅠ和PSⅡ反应中心是产生ROS 的主要场所[11]。低温胁迫会改变叶绿体内PSⅡ的还原状态,使其产生高水平的ROS,并对光合机构造成破坏[12-14],从而降低植株对光能的吸收效率,同时抑制暗反应中CO2的固定,原本传递给烟酰胺腺嘌呤二核苷酸磷酸(氧化型,NADP+)的电子泄露给氧分子,使其发生单电子还原,产生大量ROS。在
植物的非光合组织中,线粒体是ROS 产生的关键来源。植物线粒体电子传递链上的许多位点都能产生ROS,其包括4 种复合物,分别是NADH 脱氢酶(复合物Ⅰ)、琥珀酸脱氢酶(复合物Ⅱ)、辅酶Q- 细胞色素bc1 还原酶(复合物Ⅲ)和细胞色素c氧化酶(复合物Ⅳ),其中复合物Ⅰ和复合物Ⅲ是ROS 产生的主要位点[15]。低温胁迫会导致其组分的抑制或修饰,最终导致电子载体的过度还原,从而引发ROS 的生成。其中,小部分电子自呼吸链酶复合体Ⅰ和Ⅲ漏出,生成具有较强氧化作用的O2·,并通过特定的化学反应生成·OH、H2O2[16]。
此外,低温胁迫导致细胞内产生的ROS 可作为重要的信号分子参与细胞逆境响应过程,传递低温信号[17],并受质膜上Ca2+激活调节,通过细胞质中Ca2+浓度的变化来将外界刺激转化成植物可以感知的内部信号[18],介导植物完成一系列生理生化反应,从而实现植物对低温环境刺激的感知和响应。
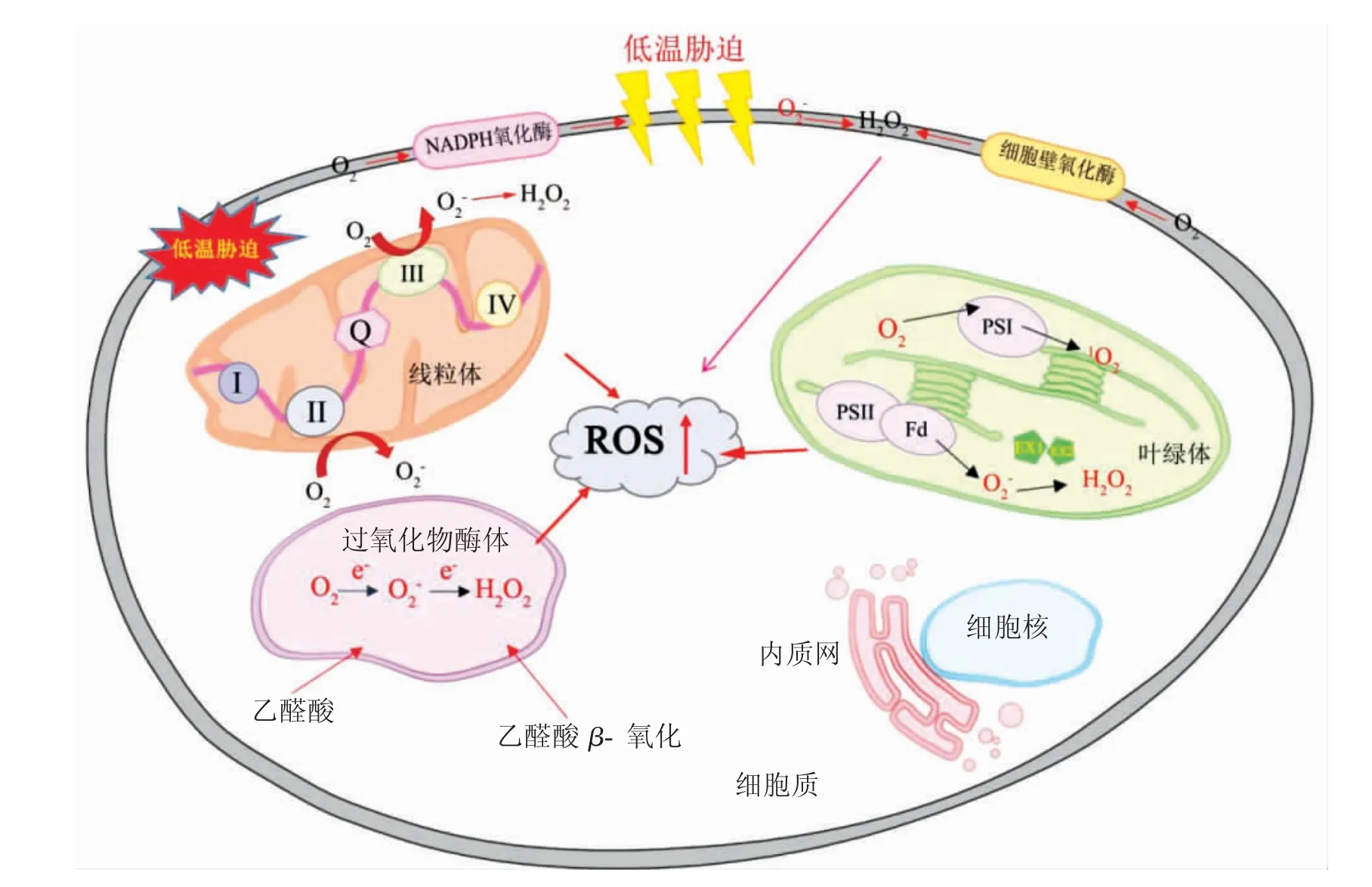
图1 低温胁迫下细胞内ROS 的产生(改自王福祥等[19])
1.2 ROS 的功能
在正常环境下生长的小麦体内,ROS 的生成与消除可通过调节其抗氧化酶系统来维持动态平衡[8]。轻中度低温胁迫下,ROS 在植物体内起着关键的调控作用,可作为信号分子感应外界低温环境的变化,诱导植物表达防御基因,从而激发抗氧化系统发挥作用,消除过量ROS 的不利影响[8]。但随着低温胁迫持续时间和强度的增加,小麦体内ROS 的积累达到一定阈值,超出细胞自身的清除能力,此时ROS 便转换为“细胞杀手”,使小麦整个氧化还原系统的代谢紊乱[20]。重度低温胁迫下,ROS 会触发细胞程序性死亡信号[21]。细胞程序性死亡(programmed cell death,PCD)是植物生长周期中的必要过程,虽然不利于作物产量形成,但它可以消除受损细胞,重新分配营养物质,从而为作物生长提供有利条件[22]。此外,ROS 可通过调节离子通道的关闭、改变细胞内的氧化状态及与受体蛋白结合等方式来发挥第二信使的调控作用[23]。
1.3 ROS 的清除
小麦的抗氧化系统主要包含酶促系统和非酶促系统(图2),可缓解因ROS 过度积累对细胞造成的氧化损伤。低温胁迫下,小麦抗氧化防御系统被激活,且通过体内酶活性的改变来抵御低温胁迫。在这些抗氧化酶系统中,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)是最主要的3种抗氧化酶,三者协同作用可有效减缓低温逆境下小麦的氧化损伤。SOD 是ROS 清除的第一道防线,它是首个响应逆境胁迫和参与反应的酶,可催化体内的O2·发生歧化反应,将O2·转化为O2和H2O2,减轻自由基造成的伤害,维持细胞膜结构和功能的稳定性[24]。POD 能够以酚类物质为底物分解H2O2[25]。而CAT 作为一种多功能酶,主要负责过氧化物酶体中H2O2的清除[26]。此外,非酶物质如抗坏血酸(AsA)、谷胱甘肽(GSH)、脯氨酸、类黄酮等可与ROS 直接反应或作为酶的底物清除ROS。AsA-GSH 循环可通过AsA/脱氢型抗坏血酸(DHA)、GSH/氧化型谷胱苷肽(GSSG)和还原型辅酶Ⅱ(NADPH)/NADP+的氧化还原来不断生成AsA 和GSH,从而维持细胞内ROS 的代谢平衡[19]。因此,小麦遭遇低温逆境时,可以通过增强抗氧化系统有效地清除ROS,从而维持小麦植株的正常生命活动。
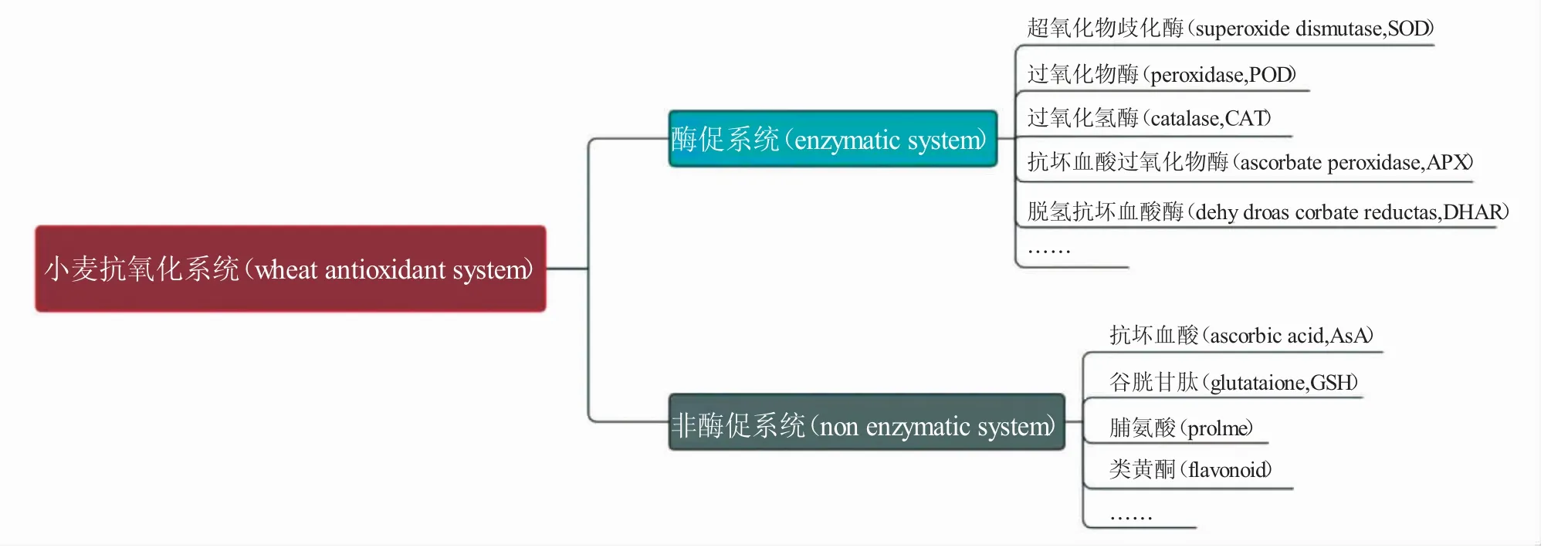
图2 小麦抗氧化防御系统
2 低温胁迫下ROS 对小麦生长发育的影响
2.1 叶片
作为对低温最敏感的生理过程之一,光合作用是小麦产量形成的基础,小麦生物量的90%以上来自光合产物[27-28]。叶片是小麦进行光合作用的主要器官,低温胁迫会破坏叶片叶绿体结构,造成叶片光合器官的损伤,对光能的吸收、传递和利用效率降低(图3)[29]。叶绿体对低温非常敏感,其不仅是进行光合作用的细胞器,还是环境胁迫的传感器[30]。叶绿体类囊体中PSⅠ和PSⅡ反应中心是ROS 产生的主要场所[29]。而低温能改变叶绿体内PSⅡ的还原状态,原本传递给NADP+的电子泄露给氧分子,使其发生单电子还原,产生大量的ROS[32]。另外,叶绿体中产生的1O2会引起核基因表达的重新编程,导致失绿和PCD,并通过2 种与类囊体膜相关的核编码叶绿体蛋白质EXECUTER1(EX1)和EX2,诱导相关反应[33]。
2.2 穗部
作为小麦光合同化物的重要储存器官,穗分化的小穗数和小花数与产量的关系极为密切。倒春寒发生时,小麦穗部正处于生长发育的敏感时期[34],当外界环境低温达到一定的阈值时,花药内产生大量ROS,可引起小麦花药氧化还原体系的紊乱,造成花药的氧化胁迫(图3)。大量研究表明,花药内ROS 的过量积累是引起花药败育的主要因素之一[35-36]。刘子涵等研究发现,D2型细胞质是细胞质雄性不育小麦的重要胞质来源,ROS 的过量积累诱导抗氧化酶基因的异常表达,造成小麦绒毡层细胞的延迟凋亡和小孢子的结构异常,最终导致D2型细胞质雄性不育系的花粉败育[37]。Wang 等研究发现,小麦生理型雄性不育系中花药内O2·的生成速率和H2O2的含量均显著高于相应的保持系[35]。张自阳等研究发现,小麦的抗氧化酶系统不能及时清除具有危害的O2·和H2O2,使小麦幼穗组织细胞受到伤害,导致穗部结实率下降[38]。因此,低温逆境下小麦花药内会积累大量ROS,造成花药的氧化损伤,导致正常发育的花药发生细胞凋亡,最终导致小花败育无法结实。
2.3 根系
根系是小麦吸收水分和运输土壤N、P、K 等矿质营养的主要器官,其生理活性直接影响小麦地上部的生长和生物量的积累,进而影响小麦的产量和品质[39]。低温胁迫会导致小麦根系细胞中ROS 代谢失调,根系活力下降,地上部分生长迟缓(图3)。裴红宾等研究表明,低温胁迫后小麦根系SOD 和POD 活性显著提高,这可能是因为低温导致小麦根细胞中ROS 积累,从而激活体内的抗氧化酶系统产生适应性反应,调节其生理代谢过程,减缓逆境伤害,但随着胁迫时间的延长,保护酶系统受到抑制[40]。冯汉清等研究发现,小麦根系中H2O2和MDA 含量不断上升,导致小麦根系的细胞死亡和生长减缓[41]。张文静等研究指出,分蘖期和拔节期低温胁迫,小麦根系产生应激反应,SOD、POD 和CAT 活性均显著提高,且抗寒性强的品种对于低温胁迫的适应调节能力强于抗寒性弱的品种[42]。此外,低温胁迫会导致根系细胞膜的脂质过氧化程度加剧,细胞内营养物质流失,透性增大,相对电导率升高,造成小麦根系膜系统结构的损伤。姜丽娜等研究表明,低温胁迫后小麦根系相对电导率均升高,说明随着胁迫温度的降低,根系细胞膜受伤害程度增大[43]。因此,低温胁迫下根系中的ROS 大量积累,致使膜脂中不饱和脂肪酸发生过氧化作用,造成膜系统结构及功能的损伤,最终导致小麦的根系养分吸收受阻,地上部生长缓慢,植株细弱。

图3 ROS 对小麦植株的伤害
3 外源化调剂在低温胁迫下小麦ROS 清除中的应用
应用外源化学调控物质可以通过激活小麦抗氧化系统来增强小麦抗寒能力,是缓解低温胁迫的有效手段。荆恩恩等发现外源氯化胆碱(CC)可减少MDA 含量和降低O2·产生速率,增强SOD、POD 和CAT 活性,有效减少膜脂损伤和缓解低温胁迫对小麦的伤害[44]。还有研究表明,外源水杨酸(SA)、脱落酸(ABA)、茉莉酸(AsA)、茉莉酸甲脂(JA)可显著降低低温胁迫后小麦叶片H2O2的含量和O2·的产生速率,提高植株的抗寒能力[45-46]。Foyer 等研究表明,低温胁迫下外源ABA 通过上调小麦叶片和分糵节中AsA-GSH 循环中关键酶的活性并诱导编码vtc1基因的表达,进而减轻冷胁迫所诱导的氧化胁迫对植物的伤害[47]。因此,应用外源化学调控物质可降低低温逆境下植物体内ROS 的含量和产生速率,增强小麦的耐寒能力,在一定程度上减缓或抑制低温对小麦的胁迫作用。然而,ABA、SA 等化调剂使用成本较高,无法在实际生产中大范围推广和应用。因此,在未来的研究中,应寻找效益好且可大批量投入大田生产中的化调物质,同时与无人机飞防相结合,提升我国小麦栽培管理和生产服务能力。
4 结论与展望
作为世界三大粮食作物之一,小麦的丰产稳产与优质对保障全球粮食安全具有极其重要的意义。尤其在全球变暖的趋势下,低温事件频发,严重影响小麦生产。本文系统阐述了低温胁迫下小麦植株活性氧的作用机制,基于源库理论从叶片、穗部、根系等方面阐述活性氧对小麦的危害机理以及小麦抗氧化系统对活性氧的清除机制。研究表明,ROS作为信号分子,除了介导环境胁迫的响应外,对调控植物的发育进程也起着重要作用,且与不同类型的植物激素产生关联。此外,也有相关研究利用ROS 来提高转录因子表达量、增加转录因子稳定性、增强转录因子结合DNA 能力来调节转录因子的功能[48],进一步调节逆境响应相关基因表达,从而改善植物抗逆性,维持作物的稳产和高产。通过基因工程技术调控活性氧的水平来提高植物的抗逆性会有很广泛的应用前景,但当前仍有许多问题尚待进一步阐释,对ROS 在响应非生物胁迫方面的研究还需要进行深入探索。
猜你喜欢
麦类作物学报(2022年8期)2022-08-08
做人与处世(2022年6期)2022-05-26
疯狂英语·新阅版(2022年5期)2022-04-29
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年2期)2016-04-11
天津农业科学(2015年9期)2015-09-02
高中生学习·高三版(2014年3期)2014-04-29
