
升金湖消落带植被特征及与土壤特性的关系
2021-03-11 08:58张黎明皇甫超河远勇帅孟盈盈
草业科学 2021年1期
张黎明,皇甫超河,远勇帅,孟盈盈,贾 璇
(安徽大学资源与环境工程学院,安徽 合肥 230601)
淡水湖泊是全球重要的生态系统,提供了大量的生态和经济服务:减轻洪水发生、固碳、净化水体和维持生态系统稳定等[1]。作为该系统的主要生产者,消落带植物群落影响着湖泊生态系统的结构和功能[2]。一直以来,对消落带植物群落形成机制的研究是湿地生态系统研究的核心问题,能够为湖泊湿地的保护提供重要的理论依据[3]。
湿地植物群落与湿地的水文、地貌和物理化学环境相互作用创造了多样化的生境[4],为许多其他生物提供了定居、庇护、产卵场所和丰富的食物资源[5]。土壤是植物赖以生存的基础,为植物生长发育提供所需的养分和水分,是生态系统中物质与能量交换的重要场所[6]。土壤性质的变化会导致植物群落发生改变。例如,土壤含水量、pH、有机质以及养分的差异都会导致植物群落的组成和结构发生改变。另外,土壤微生物在植物群落的组成与结构方面也起着重要作用[7]。现有理论和试验证据表明,消落带植物群落组成与结构可能受多种因素控制[7]。在湖泊湿地中,水位波动极大地影响了湿地植被分布格局。这主要通过改变土壤的氧化还原状态、含氧量和植物光利用率等,进而促进或抑制植物的生长和发育[8]。另外水位波动还直接影响土壤水分含量,导致土壤的通气状态以及相关的物理和化学反应在水位变化中产生梯度[3]。因此研究水位波动对植被的直接和间接影响,对理解消落带植物群落的组成与结构具有重要意义。
土壤具有空间异质性,结构复杂性和多样性的特征[9]。水平和垂直养分特征受多种环境因素的影响,例如水文、地形、气候和土壤质地[10]。现有研究表明,土壤碳(C)、氮(N)和磷(P)是生态系统中3 个最重要的化学元素,与生态系统的结构和功能密不可分。因此,C、N 和P 及其比值的变化,可能反映了生态系统的特征[11]。不同湿地生态系统的环境不同,导致土壤化学计量分布格局也不同。研究认为,湿地消落带小尺度水分梯度差异可能是影响植物群落组成和多样性的重要因素[3]。已有湿地消落带土壤化学计量特征研究多集中在自然淡水湖泊中[12]。目前尚不明确人工干预的湖泊型湿地中土壤C、N 和P 的化学元素分布特征随土壤含水量的变化特点,及其与植被特征的对应关系。
长江流域是中国淡水湖最密集的地区,有数以千计的浅水湖泊,总面积约为15 700 km2[3],记录有约400 种水生植物[13]。升金湖作为典型的浅水通江湖泊,冬季枯水期最低水位比夏季丰水期最高水位低7 m 左右[14],水位的波动会导致消落带植物多样性发生改变[15]。目前对消落带的研究,多限于土地利用[16]、岸坡稳定[17]等方面,而对消落带植物群落多样性的研究较少[18]。本研究旨在探讨升金湖湖岸消落带土壤对植物多样性的影响,阐明湿地生态系统小尺度土壤特性与植被构建模式的关系,为揭示湿地植物多样性的机制提供参考。
1 研究背景
1.1 研究区域
研究区域位于安徽省东至县升金湖上湖杨峨头(30°34′ N,117°02′ E) (图1),升金湖湖区面积达132.8 km2,自西向北由3 个相连的水面(上、中、下湖),以黄湓闸为中心扩展成湖。平均海拔11 m,上湖与中湖、下湖人为筑坝隔开,中湖和下湖与长江之间由黄湓闸连接[19]。每年5 月 − 8 月为丰水期,平均水位12.5 m,11 月 − 次年4 月为枯水期,平均水位8.98 m,其他时期为平水期,平均水位10.88 m[20]。年平均降水量为1 600 mm,受亚热带季风气候的影响,降水主要集中在夏季,约占全年降水量的50%。消落带优势植物陌上菅(Carex thunbergi)一年内有两个生长季节:第一个生长季节是从春季到夏季水位上涨之前(3 月 − 5 月);第二个是秋天的水位降低之后(8 月中旬 − 11 月中旬)[6]。
1.2 采样方法
根据升金湖水位季节动态变化,于2019 年11月上旬进行野外调查,此时为升金湖水位下降2 个月之后,消落带湿生植物处于生长盛期。通过调查发现沿湖岸垂直方向的土壤含水量差距较大,研究区的地下水位落差约19 cm。因此,本研究选择了从水陆交错带一直到植被组成无明显变化为止的代表性样地[21](图1),在该样地上设置垂直于湖岸方向的1 条长100 m 的样线,样线上每隔10 m 设立1 个25 × 25 cm 的样方,同时在样线平行方向每隔100 m设置1 条样线,共设5 条重复,在每个样方中记录物种的种类和组成。根据物种的重要值[22]确定样方中优势物种并单独记录,目测估算每个物种的盖度,并将其总和作为样方中的群落总盖度。收集植物地上部带回实验室进行烘干称量。
采用埋管法测定每个样带的地下水位[23]。湖泊水位的季节性变化直接引起地下水位的同步变化,土壤含水量也表现出季节性周期变化,受地下水位影响显著[24]。采样地点的坐标通过手持全球定位系统(GPS)记录,土壤采集参照湖泊湿地灰化苔草分布区采集方法[25]使用多参数土壤水分记录仪(TZS-2x)现场测定样方植物根系密集分布层0 − 20 cm 的土壤含水量。同时在样方内随机用土钻采集0 − 20 cm土壤,混匀装入聚乙烯袋中,送到实验室测定土壤有机碳、全氮、全磷含量。
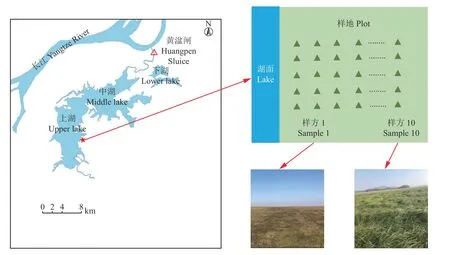
图 1 研究区域及样线布设示意图Figure 1 Schematic diagram of study area and samp le line layout
1.3 分析方法
(1) 土壤理化因子测定:
土壤带回室内风干后,手工捡出杂物、凋落物以及植物根系,过筛(2 mm),采用电位计法测定pH(土壤与蒸馏水1 ꞉ 2.5),重铬酸钾容量−外加热法测量土壤有机碳,半微量凯氏法测量土壤全氮,HClO4-H2SO4消煮法、钼锑抗比色法测量土壤全磷。
(2) 群落物种多样性指数:
对5 条样线的50 个植物群落,利用下列公式,计算每个样带的物种多样性。

式中:S 为群落中物种数,P 为样方中所有物种的个体总数和,Pi为样方内种i 的个体数。由于个体数对于陌上菅和蓼子草(Polygonum criopolitanum)等植物难以计算,因此采用植物盖度数据[26]代替个数计算。植物盖度数据使用改良的Daubenmire 百分比覆盖量表:1,≤ 1%;2,2%~5%;3,6%~25%; 4,26%~50%; 5,51%~75%;6,76%~95%;7,> 95%。先目测估计每个样方中每个物种的覆盖率。然后通过Daubenm ire 标度转换来确定每个物种的覆盖率[27]。
1.4 统计分析
运用SPSS 21.0 对每个样带的土壤特性(土壤含水量、pH、土壤有机碳、全氮、全磷)进行分析,用平均值 ± 标准差(SD)展示,同时使用Shapiro-W ilk 检验验证了数据的正态性Pearson 相关性分析,用于评估土壤化学计量特征(土壤有机碳、全氮、全磷、C ꞉ N、C ꞉ P,N ꞉ P)与土壤含水量、pH 的关系,单因素方差分析检验土壤特性之间差异,并用LSD 方法检验单一变量样方之间的差异性,排序法用来确定土壤因子对样方群落植物多样性的影响,通过判别成分分析法(dynam ic component analysis, DCA),得出所有排序轴梯度长度小于3,因此采用冗余分析法(redundancy analysis, RDA)确定主要影响因子。Monte Carlo 检验(9 999 次置换)检测样方群落植物多样性和土壤因子是否存在显著相关关系,排除影响不显著的变量(P > 0.05)。上述统计分析在SPSS21.0、CANOCO 4.5 软件中完成,图表绘制利用Excel 软件完成。
2 结果与分析
2.1 不同水位梯度下植物多样性及土壤理化性状
如表1 所列,随着地下水位的降低,群落Shannon-W iener 指数与均匀度指数表现为减小的趋势。群落Shannon-W iener 指数与均匀度指数的最低值均出现在土壤含水量较低的9、10 样方中。在地下水位与土壤含水量较高的样方中,荸荠(Eleocharis dulcis)在群落中占据优势地位,陌上菅则在地下水位与含水量较低的样方中占据优势地位。随着地下水位上升,样方优势物种逐渐由中生植物陌上菅向湿生植物荸荠转变。对地下水位、土壤含水量、土壤有机碳、全氮、全磷进行单因素方差分析结果表明(表1),土壤含水量、土壤有机碳、全氮、全磷均表现出显著(P < 0.05)差异,而土壤pH 则无显著差异(P = 0.962)。LSD 多重比较表明,除样方6 与7 外,其余样方地下水位均表现出显著差异。随着地下水位的降低土壤含水量总体呈现降低的趋势,样方10 相比样方1 土壤含水量减少了72%,除少数样方含水量差异不显著之外,总体表现出显著差异(P < 0.05)。土壤有机碳与土壤全磷均随着土壤含水量的增加呈现出降低的趋势,样方1 相比样方10 土壤有机碳下降了21%,土壤全磷下降了42%,样方1 到样方10 土壤有机碳与全磷含量显著增加。样方中土壤全氮的含量随土壤含水量的增加呈现增加的趋势,样方1 相比样方10 上升了144%。
2.2 土壤含水量与化学元素及化学计量比的回归分析
对土壤含水量与土壤有机碳、全氮、全磷及其比值进行多元线性回归分析(图2)得出,土壤有机碳、土壤全磷、土壤C ꞉ N、土壤N ꞉ P 与土壤含水量的最佳拟合曲线为幂函数,土壤有机碳、土壤全磷与土壤C ꞉ N 随着土壤含水量的增加而下降,土壤全氮与N ꞉ P 随着土壤含水量的增加而升高。土壤C ꞉ P 与土壤含水量的最佳拟合为二次曲线,土壤C ꞉ P随着土壤含水量的增加而升高。
2.3 土壤因子对植物群落多样性的影响
土壤含水量、土壤全氮以及土壤C ꞉ P 和N ꞉ P与植物Shannon-W iener 指数、均匀度指数极显著正相关(P < 0.001) (表2),土壤有机碳、土壤全磷以及C ꞉ N 与植物Shannon-W iener 指数、均匀度指数极显著负相关(P < 0.001),土壤含水量、土壤全氮以及土壤C ꞉ P 和N ꞉ P 与优势物种盖度极显著负相关(P < 0.001),与优势物种生物量极显著正相关(P <0.001),土壤有机碳、土壤全磷以及C ꞉ N 与优势物种盖度极显著正相关(P < 0.001),与优势物种生物量极显著负相关(P < 0.001)。
通过排序,将土壤因子对消落带植被空间分布的解释能力进行分离,所收集的土壤因子中,土壤N ꞉ P 的解释能力为83.5%,土壤含水量为5.9%,土壤全氮为2.1%,土壤C ꞉ P 为1.6%,土壤全磷为0.2%,SOC 为0.1%。Monte Carlo 随机置换检验表明,所有典范特征轴与样方植物群落变化都极显著相关(P =0.002) (表3)。RDA 排序前两轴的特征值之和占全部排序轴特征值总和的93.48%,包含了排序的绝大部分信息,其中轴1 的特征值占全部特征值总和的91.85%,因此采用前两轴的数据来分析植物群落与环境因素的关系(图3),根据土壤因子与RDA 排序轴的相关性分析可知,土壤含水量、土壤全氮以及土壤C ꞉ P 和N ꞉ P 与样方中物种多样性指标相关性最高(表4、图3),表明在消落带上土壤含水量及土壤N ꞉ P 是决定植物多样性变化的主导因子,土壤全氮和土壤C ꞉ P 对植物多样性变化也具有一定的影响。
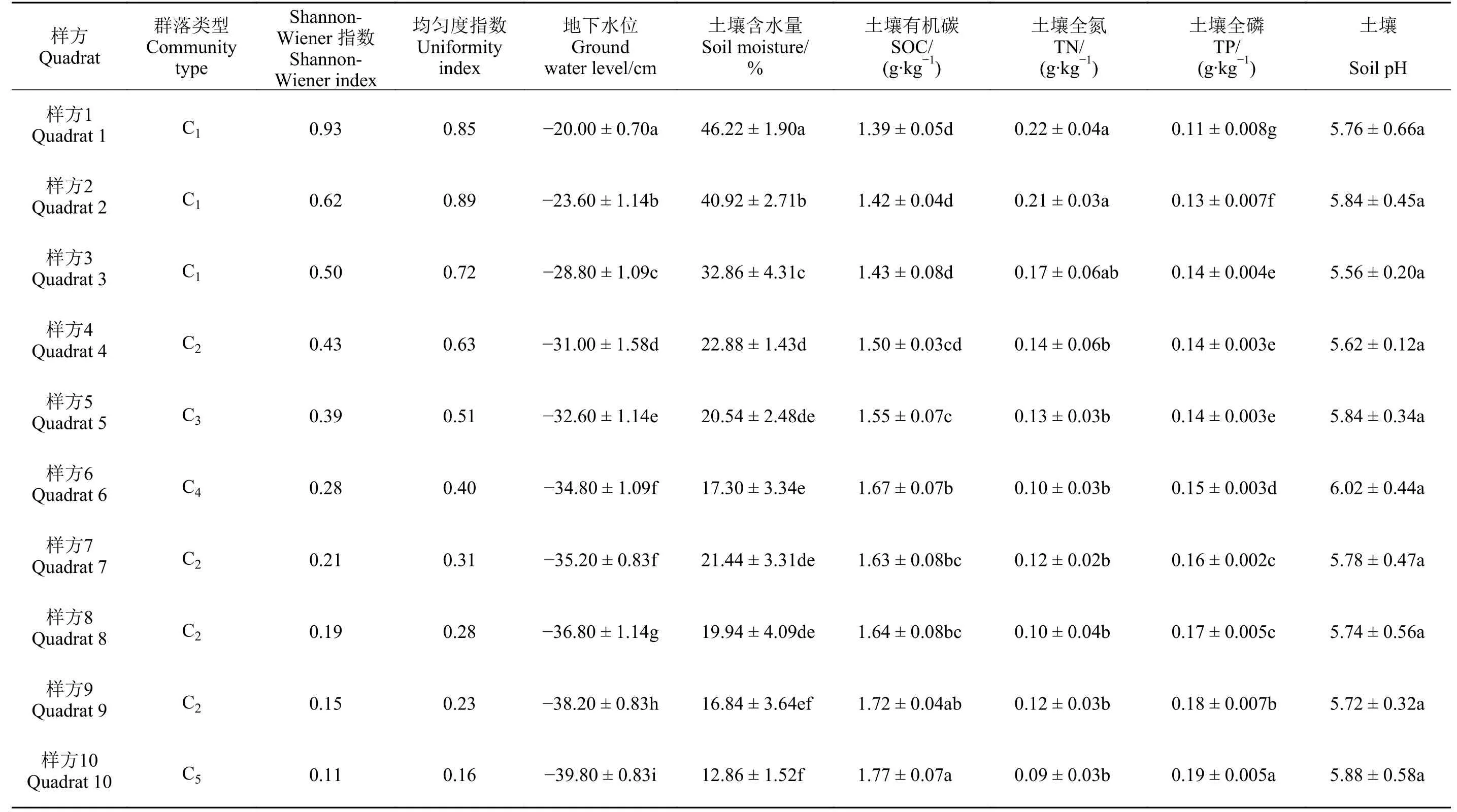
表 1 样方物种多样性与土壤环境因子Table 1 Sample species diversity and soil environmental factors
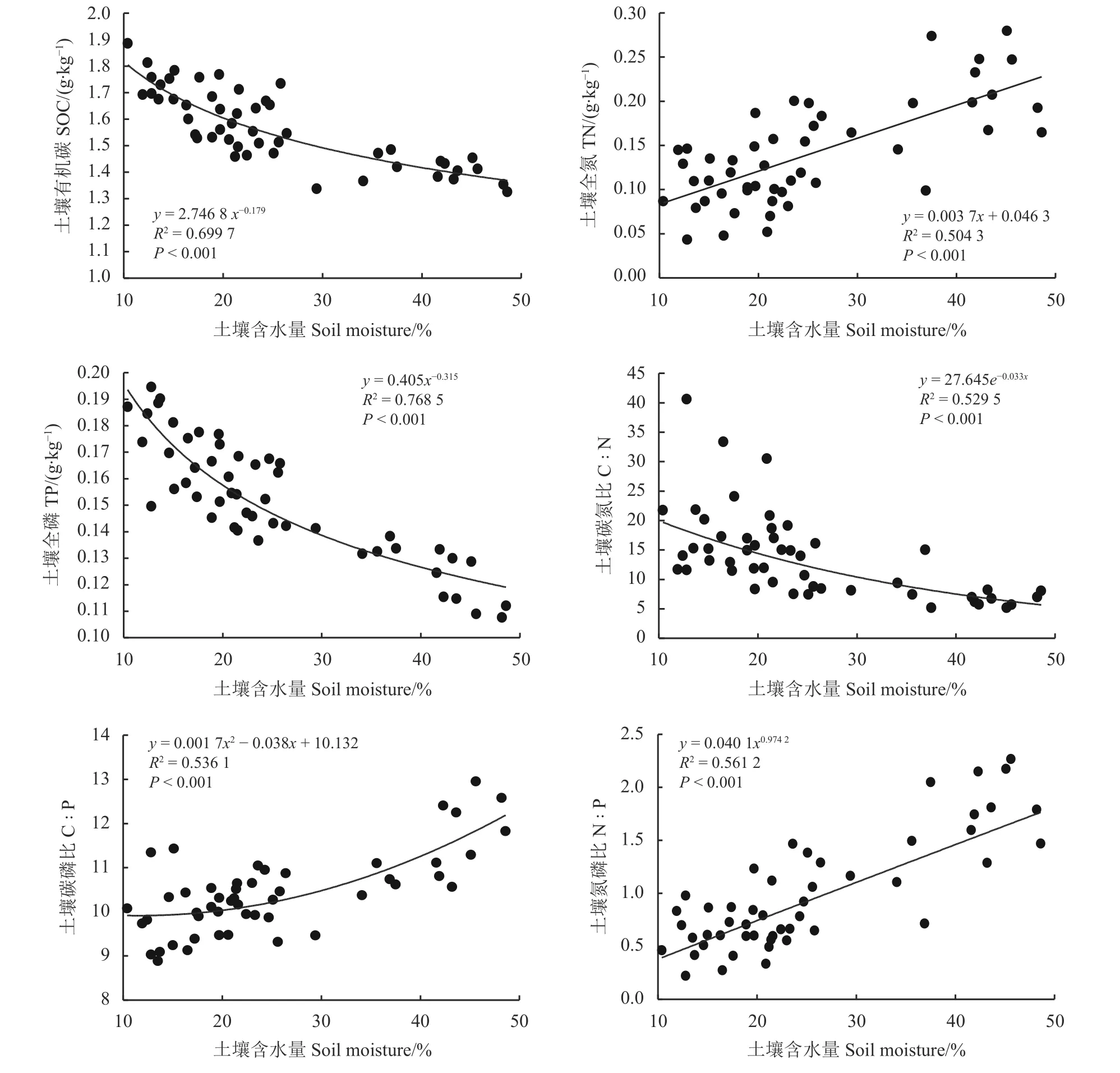
图 2 土壤含水量与土壤化学元素及其比值的曲线拟合Figure 2 Curve fitting of soil moisture and soil chem ical elements and their ratios
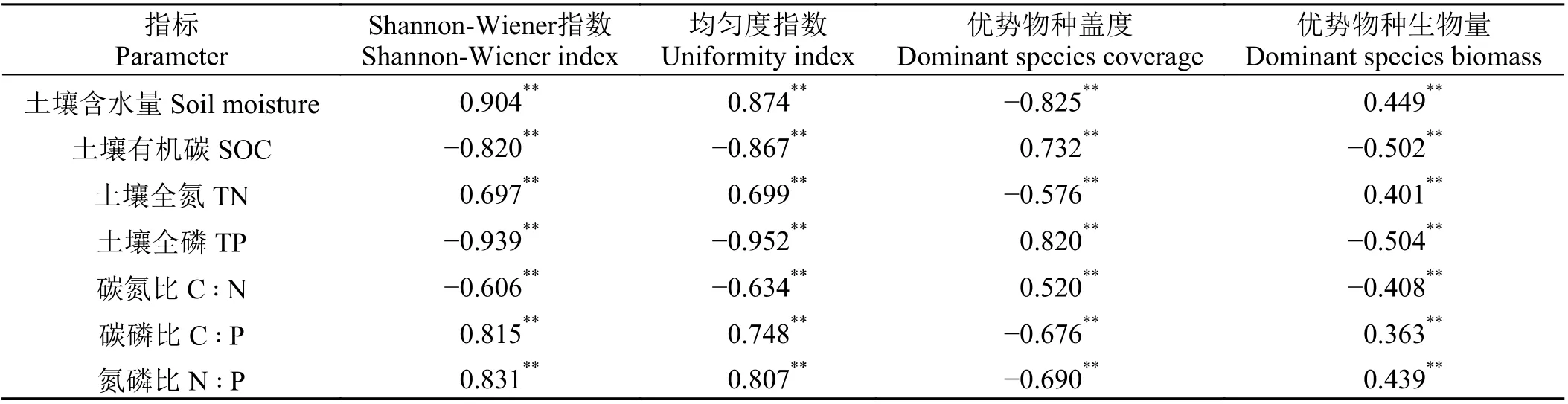
表 2 土壤含水量以及化学元素及其比值与物种多样性指标之间的Pearson 相关性Table 2 Pearson correlation between soil moisture, stoichiometric elements, and their ratios w ith species diversity indicators
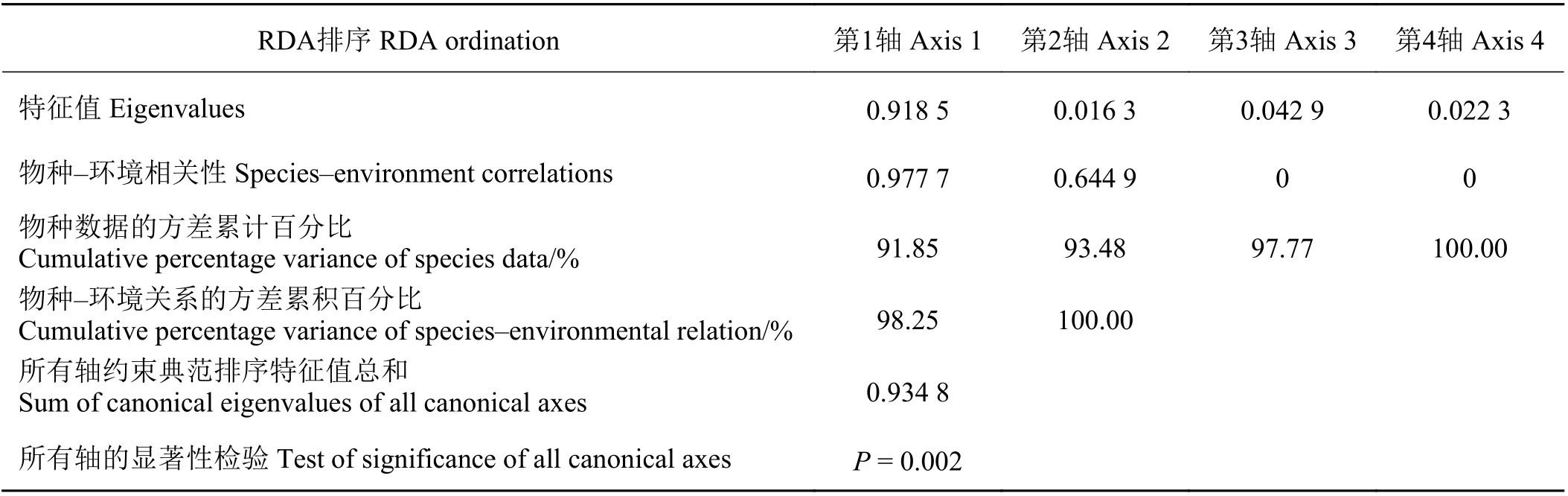
表 3 升金湖消落带植物多样性RDA 排序Table 3 RDA ordination of p lant diversity in the riparian zone of Shengjin Lake
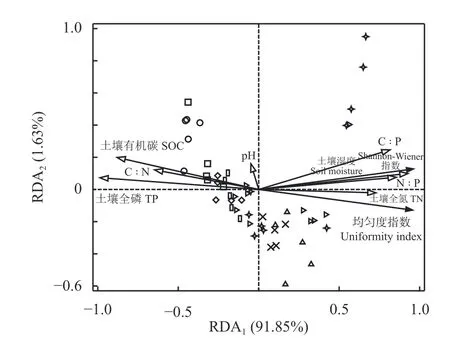
图 3 样方群落植物多样性与土壤因子RDA 排序图Figure 3 Ordination diagram of p lant diversity and soil factor RDA of the sam ple community
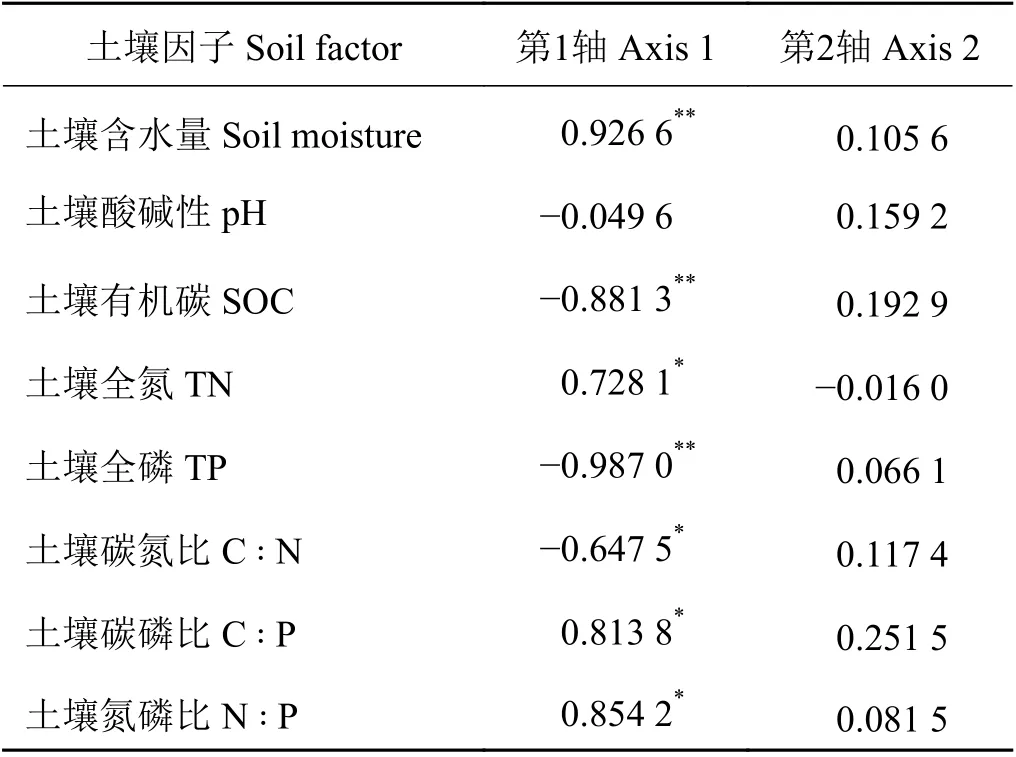
表 4 土壤因子与RDA 排序轴相关性Table 4 Correlation between soil factors and RDA ranking axis
3 讨论
3.1 不同水位梯度下植被与土壤的分布特征
研究发现,地下水位对湿地植被多样性的形成非常重要。地下水位较高的样方植物多样性要高于地下水位较低的样方(表1),并且随着地下水位的升高,样方中植被由湿生植物向中生植物转变,这可能是由于人为设置闸口控制湖泊水位,减少了湖岸湿地的面积,增加了中生植物侵入原先湿生植物生境的风险[6]。此外,随着水位上升植物多样性增加,这可以由中度干扰假说来解释[28]。具体来说,样方1 的地下水位波动可以看作是中度干扰,水位的升高增强了生态位的异质性,减少了种间竞争,从而增加了植物多样性[6]。地下水位的变化对土壤含水量也有着直接的影响(表1)。有证据表明,土壤含水量和化学元素等环境因素在调节植物群落结构中起着决定性的作用[29]。实际上,土壤含水量和化学元素对植物的影响不是独立的。土壤水文会通过控制土壤通气状态来改变土壤化学元素的梯度、养分可用性等[30]。本研究发现土壤含水量与土壤碳、氮、磷元素显著相关(表1、图2)。土壤的含水量较高,有利于可溶性有机碳的溶出并为有机碳的矿化过程提供能量[31]。另外,土壤含水量较高容易造成厌氧的环境,不利于土壤有机质的矿化,导致土壤中全氮的积累[32-33]。土壤中磷的含量受到母质[33]、气候、生物[34]和土壤中的地球化学过程[35]等一系列因素的影响。本研究中,土壤全磷与土壤含水量极显著负相关(图2),这可能是土壤含水量增加加快了土壤风化速率和磷元素的淋溶,从而导致土壤全磷含量的降低[36]。土壤pH 是影响土壤肥力和微生物活动的重要因素之一,能够直接影响碳、氮、磷的存在形态和转化方式,同时对土壤养分元素的有效性产生一定的影响[37]。研究区内土壤pH 与土壤含水量、有机碳、全氮、全磷及其比值相关性不显著,可能与pH 的变化范围较小有关[37]。土壤C ꞉ N是土壤有机质组成和质量程度的一个重要指标[38],也是预测有机质分解的一个很好的指标,通常与有机质的分解速率成反比,土壤含水量增加有机质分解速率加快[39],因此,会导致土壤C ꞉ N 的下降,本研究也得到一直结论(图2)。土壤含水量显著影响了土壤N ꞉ P 和C ꞉ P,土壤含水量增加有机碳减小的程度要大于全磷减小的程度(表1),表现为C ꞉ P随土壤含水量的增加而上升。土壤含水量增加导致全氮增加,因此土壤N ꞉ P 表现随土壤含水量的增加而上升。
3.2 土壤因子对物种多样性影响
RDA 分析结果表明,土壤含水量、土壤N ꞉ P 以及土壤全氮含量和C ꞉ P 是决定植物多样性变化的主导因子。本研究时间处于该湖泊的枯水期,土壤含水量较丰水期低,因此土壤含水量可能是影响植物群落组成的重要因素[40]。土壤含水量的RDA 排序轴显著正相关(表4),在对鄱阳湖湿地植物的研究中也发现了这一结论,土壤含水量较高,优势物种与伴生种对水分需求的生态位差异[41],导致了优势物种的盖度减少,为伴生种的生长提供空间从而使得群落物种多样性增加[42]。
土壤中的化学元素作为重要的环境控制因子,为植物的生长提供了物质和能量。沈瑞昌等[6]在对鄱阳湖的研究中指出,土壤含水量通过改变土壤化学元素或植物生物量间接影响植物多样性。根据RDA 分析的结果,土壤中氮元素与植物多样性显著相关(表2),并且氮元素与土壤含水量具有显著的线性关系(图2),表明土壤含水量与土壤氮元素之间存在一定的耦合关系。因此,土壤含水量与土壤氮元素会单独或共同对植物群落产生影响。研究表明,湿地土壤氮元素的转化受多重因素的影响,邢梦林等[43]研究发现河滨湿地中,在一定范围内土壤含水量越高,硝化速率越快。也有关于土壤含水量与反硝化速率成正相关的报道[44]。W illiams 等[45]发现沼泽湿地中氮含量受水位的影响显著, 并随水位增加而增加。研究区内土壤氮含量与植物多样性显著正相关(表2)。可能是由于土壤含水量的增加,土壤中有机氮的矿化过程加快[46]。由于研究区内不同的植被类型、多变的水文过程以及氮元素转化的多样性和复杂性,其具体转化过程有待进一步研究。另外RDA 分析表明,土壤N ꞉ P 是决定植物多样性变化的主要因子,N ꞉ P 是限制性养分的判断的重要指标[38],研究区内磷的含量变化幅度相对较小(表1)。因此,相对而言,氮元素对植物生长的影响可能要大于磷元素,而氮元素在土壤含水量增加下提高了物种多样性,因此N ꞉ P 的增加也有利于物种多样性的增加(图3)。
4 结论
土壤含水量、全氮、C ꞉ P 和N ꞉ P 在不同地下水位梯度上具有显著差异,土壤含水量、全氮随地下水位升高有显著增加的趋势。土壤含水量对土壤化学元素及其比值具有一定的影响,土壤全氮、N ꞉ P以及C ꞉ P 与土壤含水量极显著正相关。土壤含水量与土壤N ꞉ P 是决定该区域植物多样性变化的主要限制限制因子,同时土壤全氮和C ꞉ P 对物种多样性的变化也起到一定的作用,其中土壤含水量可能直接影响植物群落的多样性,同时也通过影响土壤氮的转化以及磷元素的有效性间接影响消落带植物多样性。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
浙江农业学报(2016年7期)2016-06-15
现代农业(2016年3期)2016-04-14
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
土壤与作物(2015年3期)2015-12-08
环境科技(2015年2期)2015-11-08
