
hUC-MSC在成神经分化过程中的趋化迁移研究
2021-03-09 00:21李晨卞凯鹏熊英张丽娟匡梦岚王岚王恒许键炜
癫痫与神经电生理学杂志 2021年1期
李晨,卞凯鹏,熊英,张丽娟,匡梦岚,王岚,王恒,许键炜
间充质干细胞是一种源自发育早期中胚层,主要分布于骨髓、脂肪、脐血等组织,是具有自我更新、多向分化、免疫调节能力的干细胞[1-2],体外培养扩增的间充质干细胞作为组织工程的种子细胞,可修复各组织和器官,也可用于器官或者组织移植。可以在脂肪、骨髓等组织中分离[3-4]。近来有研究表明,人脐带间充质干细胞(human umbilical cord mesenchymal stem cells,hUC-MSC)不仅有间充质干细胞的特点,还有其独特的优势,如取材简单、细胞增殖迅速、没有伦理问题、连续传代后能维持细胞干性、低免疫原性、在体内不引起畸胎瘤等[5]。其可以稳定分化为神经细胞、肝细胞和软骨细胞等[6-7]。据研究表明,SDF-1与细胞信号传导、胚胎发育、细胞迁移、调节免疫、造血及血管生成相关[8]。
1 材料与方法
1.1 材料
1.1.1 脐带
新生儿脐带来源于贵州医科大学附属医院妇产科。通过医院人体伦理委员会批准,并经产妇知情同意。
1.1.2 仪器
倒置荧光显微镜、Leica Logo型荧光显微镜及成像系统(德国Leica公司);流式细胞仪(美国Beckman公司);含有体积分数为5%CO2培养箱(美国Thermo公司)。
1.1.3 试剂L-DMEM培养基
Gibco公司;RPMI培养基:Gibco公司;FBS:BI公司100U/mL青-链霉素:Gibco公司;0.25%胰蛋白酶(Ttypsin-EDTA):Gibco公司;2%二甲基亚砜 (DMSO) 和200 μmol/L 丁羟基茴香醚(BHA) Solarbio公司; FITC标记的抗人CD105、CD29、CD45和CD34抗体(BD公司):SDF-1 Signaling公司。
1.2 方法
1.2.1 hUC-MSC的原代培养及传代
取健康安全的脐带(足月儿,不带细菌、病毒等),无菌生理盐水清洗脐带表面,去除血凝块,清洗三遍后在75%酒精中浸泡30 s,用无菌生理盐水洗去残余酒精。将清洗后的脐带剪成3 cm长的小段,浸泡于无菌生理盐水中,每条脐带上有一条粗静脉和两条细动脉,将其剥离丢弃,再剥离血管周围的华通氏胶,并留取用来原代培养。用无菌的眼科镊将华通氏胶放入无菌的玻璃小瓶,并加入1 mL的培养基,用无菌眼科剪将华通氏胶剪碎成约1 mm3大小的组织块,将剪碎的华通氏胶均匀地铺在培养瓶中,然后放入37 ℃,5% CO2培养箱中培养。6 d后待原代细胞从华通氏胶组织块爬出后对其进行半换液,再待其融合至80%以上,可以对其进行传代培养。收集P3-P7代传代细胞冻存。
1.2.2 流式细胞仪检测细胞表面分子标志物
收集第3代hUC-MSC,经胰酶消化后离心(1 200 r/min×5 min)使细胞重悬,调整细胞浓度为1×106mL-1,分别加入各种流式抗体:CD105、CD29、CD34和CD45等,常温下孵育30 min,0.1 mol/L PBS冲洗数次后,将其与FITC标记(或PE)的二抗在避光条件下作用30 min,PBS充分洗涤后立即送检。利用倒置显微镜观测P3代的hUC-MSC的细胞形态见图1。
1.2.3 HUC-MSC诱导分化为成神经细胞
将细胞诱导过程设为A、B、C、D、E五个点(图2)。选取生长状况良好的第4代hUC-MSC按5×104/mL接种于含包被PLL盖玻片的35 mm培养皿中。用hUC-MSC生长培养基(L-DMEM+10% FBS)培养(设为A点)。
接种24 h后观察细胞生长状况,待细胞60%铺满盖玻片时,弃生长培养基,加入含10 ng/mL bFGF的诱导培养基。诱导24 h后(设为B点),以后换成无血清的诱导培养基[含2%二甲基亚砜 (DMSO) 和200 μmol/L 丁羟基茴香醚(BHA)]继续诱导6 h即诱导30 h (设为C点)。维持18 h,即诱导48 h(设为D点),之后继续维持24 h,即诱导72 h (设为E点)。另设对照组:始终以hUC-MSC常规培养基(10%FBS+L-DMEM液)培养。定期观察细胞,取诱导前、诱导24 h、诱导30 h、诱导48 h、诱导72 h的细胞进行固定,并进行神经细胞特异性标志物nestin和NSE的免疫荧光染色检测。
1.2.4 免疫荧光染色
细胞去除培养基,用PBS清洗3遍,加入4% 多聚甲醛,室温固定30 min或者4 ℃过夜。PBS洗3次后,分别滴加用PBS+叠氮化钠(NaN3)(0.02%)+牛血清白蛋白(BSA)(3%)+0.2%聚乙二醇辛基苯基醚(TritonX-100)稀释的一抗孵育,4 ℃过夜或室温2 h。PBS洗3次。加入二抗室温孵育1 h,PBS冲洗3次。室温放置5 min并冲洗。用50%甘油缓冲液封片,荧光显微镜观察拍照。本实验所用一抗和二抗稀释比例为羊抗人Nestin抗体 (1∶200);羊抗人NSE抗体 (1∶100)。FITC标记的羊抗人二抗(1∶70)。
1.2.5 Transwell研究分化细胞的趋化性迁移
Transwell 24孔板上室每孔加入300 μL(含1.0×105个细胞)用不含血清的L-DMEM培养基悬浮的诱导前、预诱导24 h、诱导5 h、维持18 h、维持48 h的不同分化状态的hUC-MSC,下室加入500 μL含有20 ng/mL浓度的SDF-1α的不含血清的L-DMEM培养基。将Transwell移入培养箱内静置6 h后用4%多聚甲醛固定,0.1%结晶紫染色30 min, PBS漂洗后于倒置相差显微镜下拍照,统计迁移至室膜下方的细胞数。将对照组的迁移细胞数标准化定义为1。
1.3 统计学方法
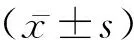
2 结果
2.1 hUC-MSC形态
P3代培养第3天,倒置荧光显微镜观察可见大量细胞贴在皿底,呈长梭状,类似于成纤维细胞,汇聚后细胞排列紧密,呈群落样生长,聚合的细胞呈漩涡状。组织块法培养原代细胞达90%~95%融合,可进行传代培养(图1)。传代数小时后hUC-MSC快速贴壁,3~5 d后细胞可再次达到80%~90%的汇合度。经过数代培养后,细胞形状无明显变化,增殖能力无明显变化。
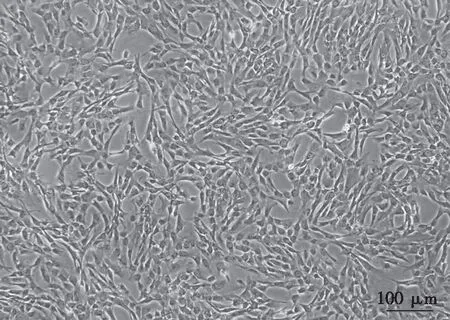
图1 利用倒置显微镜观测P3代的hUC-MSC的细胞形态(Scale bar=100 μm)
2.2 hUC-MSC表型鉴定结果
流式细胞仪检测结果显示CD34、CD45表达为阴性,而CD29和CD105表达为阳性,课题组前期已完成这部分工作[9],提示组织块法培养可获得hUC-MSC。
2.3 hUC-MSC成神经诱导过程中的形态变化及鉴定
hUC-MSC接种24 h(诱导前),细胞呈不规则多角型 (图2A、图3A)。诱导24 h,hUC-MSC由其典型形态变成长梭形,胞体出现收缩,细胞边缘变得不规整(图2B、图3B)。诱导30 h时,胞体均呈圆形或椭圆形,折光性增强,伸出细长的突起,并且多为两极突起(图2 C、图3C)。换维持培养基至48 h后,发生形态改变的细胞数目进一步增加,细胞出现3个或更多分叉及次级分叉,呈现出典型的神经元样(图2 D、E,图3 D、E)。
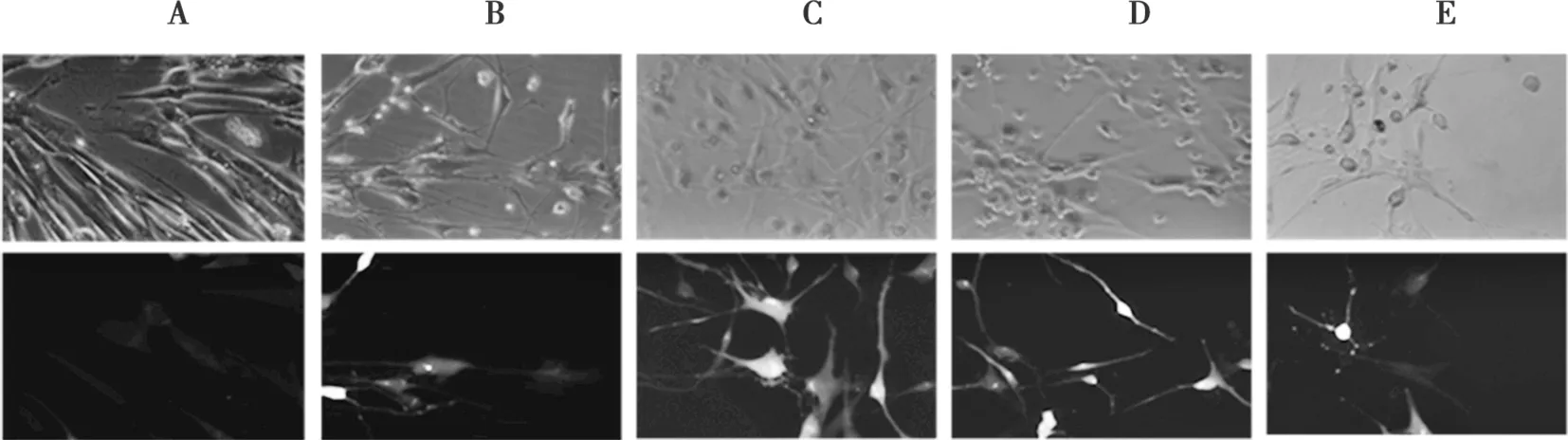
图2 免疫荧光染色检测hUC-MSC在诱导分化过程中nestin的表达情况。A:诱导前; B:诱导24 h; C:诱导30 h; D:诱导48 h; E:诱导72 h。(Scale bar=50 μm)
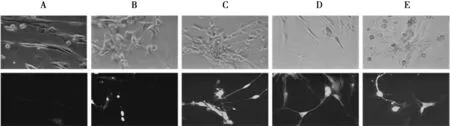
图3 免疫荧光染色检测hUC-MSC在诱导分化过程中NSE的表达情况。A:诱导前; B:诱导24 h; C:诱导30 h; D:诱导48 h; E:诱导72 h。(Scale bar=50 μm)
在hUC-MSC向神经样细胞分化的过程中,细胞形态发生了显著的变化,维持48 h的分化细胞形态与神经元非常相似。通过免疫荧光技术检测nestin和 NSE的表达情况加以鉴定。nestin存在于未分化的间充质干细胞中,约有11.5%的hUC-MSC表达nestin。在诱导24 h后,nestin阳性细胞百分比均显著上升,阳性率达到72.8%,较未诱导的对照组具有显著差异,诱导30 h后阳性率达80 %,随着维持时间的延长nestin表达率下降,维持48 h时nestin阳性率降至48.6%,维持72 h时nestin阳性率降至30.2%。整个诱导分化过程中,nestin的表达呈现出先增高后降低的趋势。NSE在诱导期和维持前期均为阴性,在整个过程中末期才开始表达,到维持48 h时有62.5%的细胞表达,维持72 h时有76.5%的细胞表达。不同诱导时间的hUC-MSC免疫荧光结果的统计见图4。
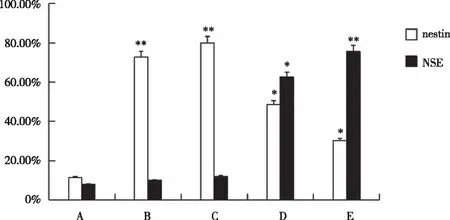
图4 不同诱导时间的hUC-MSC免疫荧光结果的柱状图A:诱导前; B:诱导24 h; C:诱导30 h; D:诱导48 h; E:诱导72 h。*P<0.05,** p<0.01,表示与对照组比较差异有显著意义。
2.4 不同时间诱导培养分化后的细胞迁移能力的测定
不同成神经分化状态的hUC-MSC迁移能力不同(图5)。Transwell趋化迁移实验显示,经过加入含10 ng/mL bFGF的预诱导培养基诱导24 h后(图5B),细胞向SDF-1α的趋化迁移能力增强,换为无血清的诱导培养基(含2% DMSO 和200 μmol/L BHA),诱导6 h(即诱导30 h)后(图5C),细胞向SDF-1α的趋化迁移能力达到最强,换成维持培养基维持18 h(即诱导48 h)后(图5D),细胞向SDF-1α的趋化迁移能力减弱,继续维持至72 h后(图5E),细胞向SDF-1α的趋化迁移能力最弱。而始终以hUC-MSC常规培养基(10%FBS+L-DMEM液)培养为对照组的各时间节点细胞趋化迁移能力与诱导组接种24 h(图5A)细胞比较差异无统计学意义。细胞迁移至膜下相差图(bar=25 μm,图5上)。不同分化状态hUC-MSC迁移至膜下情况统计分析结果见图5下。
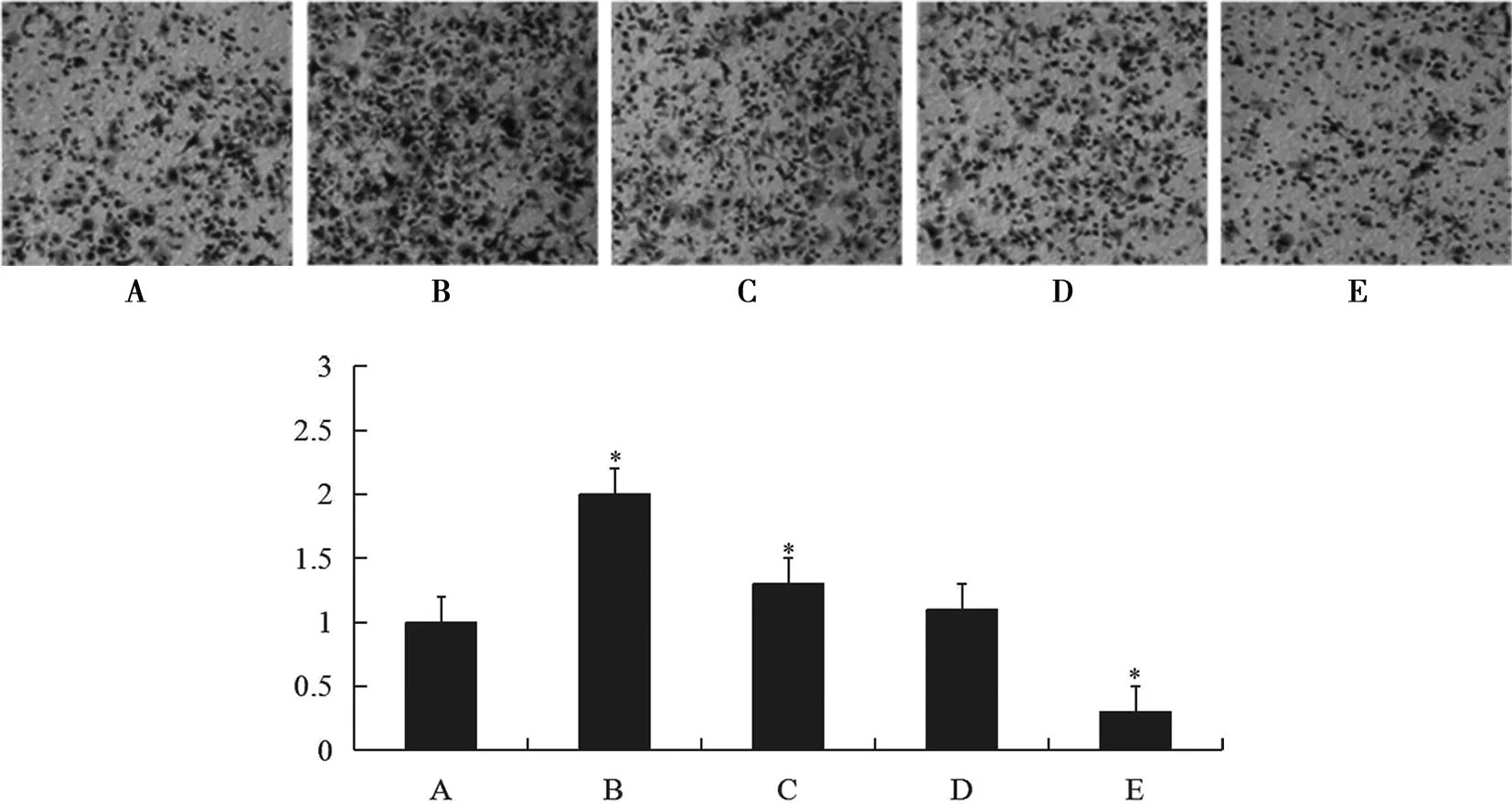
图5 不同分化状态hUC-MSC向SDF-1α的趋化迁移能力 A:诱导前; B:诱导24 h; C:诱导30 h; D:诱导48 h; E:诱导72 h。注:将未分化状态细胞(A)迁移至膜下的细胞数标准化为1;*P<0.05表示B、C、D、E组与A组相比。
3 讨论
神经细胞为不可再生细胞,一旦损伤难以再生,而且神经系统的修复能力有限,目前在临床上,一直缺少有效的治疗神经损伤的手段,难以满足临床医生和患者的治疗期望。如何最大程度地恢复或者改善损伤神经的功能是一个急需解决的问题,也是一个值得深入研究的课题。
hUC-MSC不仅有着MSC的共性,还具有取材简单、没有伦理问题、低免疫原性、在体内不引起畸胎瘤而且在体外培养细胞增殖迅速等优点,所以已经成为干细胞的理想来源[10],hUC-MSC由于低表达主要组织相容性复合体MHC-Ⅰ类分子、不表达MHC-Ⅱ分子使其具有低免疫原性;hUC-MSC在不同的病理条件下能迁移至炎症组织和肿瘤组织即细胞归巢。这一特性在细胞移植、组织工程材料、基因靶向治疗等方面具有广泛的临床应用价值[5]。本次研究所用的hUC-MSC是利用足月儿的脐带利用组织块贴壁法分离培养得到的,然后对分离得到的hUC-MSC利用流式细胞仪对细胞表面的标志物进行检测,发现细胞表面标志物CD71、CD29,CD105高表达,而CD34、CD45低表达,符合MSC的生物学特征[11]。
hUC-MSC具有成体干细胞的基本特性,可以跨胚层诱导分化为骨、软骨、真皮组织、骨骼肌、心肌细胞及神经元等多种类型细胞[12],在组织再生和创伤修复等方面具有重要作用。目前国内外多家干细胞治疗中心已开展了hUC-MSC治疗帕金森病、脊髓损伤、脑瘫、多发性硬化、老年痴呆、脊髓小脑共济失调等神经系统疾病的临床研究。
当细胞损伤时,多种相关生长因子产生,从而促进损伤的修复,本研究利用bFGF的基础诱导培养基。再利用诱导培养基(含DMSO和BHA),将hUC-MSC诱导分化为成神经细胞,并进行神经细胞特异性标志物nestin和NSE的免疫荧光染色检测[13-14]。发现在hUC-MSC向神经样细胞分化的过程中,细胞形态发生了显著的变化,维持48 h的分化细胞形态与神经元非常相似。通过免疫荧光技术检测nestin和NSE的表达情况加以鉴定。nestin存在于未分化的间充质干细胞中,约有11.5%的hUC-MSC表达nestin。在诱导分化后,nestin阳性细胞百分比均显著上升,阳性率达到72.8%,较未诱导的对照组具有显著差异,并且随着诱导时间延长,nestin阳性率也逐渐升高。说明hUC-MSCs可以稳定分化为成神经细胞。
趋化因子是指具有趋化作用的细胞因子,近年来的研究表明:趋化因子不仅在保护宿主、清除病原因子、炎症反应、过敏反应、免疫细胞的发育、分化及免疫系统的自身稳定等方面起作用,而且在肿瘤的形成及转移、移植免疫排斥、中枢神经系统神经网络的发育、心血管系统的发育和损伤的修复等方面发挥重要的作用[15-16]。SDF-1α属于α类趋化因子,SDF-1 有SDF-1α 和SDF-1β两种亚型,SDF-1α 为其主要亚型,它通过与其表面受体CXCR4特异性结合发挥作用。且在一些病理性疾病如炎症、缺血和缺氧, SDF-1 的表达量会显著升高。SDF-1 具有典型的细胞趋化作用,能趋化骨髓间充质干细胞、造血干细胞、内皮祖细胞。本文着重研究了hUC-MSC向神经样细胞的诱导分化及不同分化状态细胞向SDF-1α的趋化迁移特点[17]。我们发现诱导6 h的分化细胞对浓度为20 ng/mL的SDF-1α表现了更强的趋化迁移性,表明趋化性迁移与其分化状态紧密相关。用于临床移植,为更好归巢迁移至病灶部位的细胞分化状态选用提供了实验数据参考[18]。
本研究为探索性实验研究,实验检测了神经干细胞特异性标志蛋白Nestin和NSE,作为检测指标,说服力仍显不足,后续研究中课题组还将进一步检测微管相关蛋白、β微管蛋白Ⅲ等神经标志物以验证实验结果。但在临床转化前仍需对一些问题进行深入研究,例如如何优化间充质干细胞的来源、间充质干细胞应用时的注射剂量和细胞数量等。深入了解间充质干细胞在神经修复方面的作用机制,将会更利于探索其在周围神经损伤重建中的作用及潜在的应用价值。
猜你喜欢
数学年刊A辑(中文版)(2022年1期)2022-08-20
数学物理学报(2021年5期)2021-11-19
昆明医科大学学报(2021年3期)2021-07-22
昆明医科大学学报(2021年5期)2021-07-22
现代临床医学(2021年2期)2021-03-29
世界科学技术-中医药现代化(2021年10期)2021-03-02
中国生殖健康(2020年2期)2021-01-18
数学年刊A辑(中文版)(2020年2期)2020-07-25
健康博览(2019年10期)2019-12-02
中国生殖健康(2018年2期)2018-11-06
