
碱胁迫对唐古特白刺生长及叶片超微结构的影响
2021-03-05 12:36:22张慧芳闫海冰于兆友杨秀清
中国水土保持科学 2021年1期
张慧芳,闫海冰,冯 帆,于兆友,杨秀清
(山西农业大学林学院,030801,山西太谷)
土壤盐碱化已成为一个全球性的资源和环境问题,这一生态灾难严重制约农作物生长和林草等植被生存,打破了生态平衡及其系统稳定[1]。盐碱化土壤对植物造成的胁迫既含有以NaCl和Na2SO4等中性盐为主的盐胁迫,也包括以NaHCO3和Na2CO3等碱性盐为主的碱胁迫[2]。碱胁迫对植物形成除与盐胁迫共有的离子毒害和渗透胁迫外,还有高pH值胁迫[3]。碱胁迫造成的pH值增高严重影响植物根部结构和细胞膜透性,导致根系活力降低,甚至影响植物叶片的结构和生理功能[3],从而产生比盐胁迫更强的生态破坏力。
唐古特白刺(Nitrariatangutorum),一种生态抗逆兼果肉经济型灌木,具有抗风沙、耐盐碱、耐贫瘠等生态适应性,广泛分布于宁夏、内蒙古、新疆等地区,在保持水土、改良盐碱地及防风固沙等方面发挥着积极作用[4]。目前关于唐古特白刺的耐盐性研究多集中在盐胁迫,而对碱胁迫涉及相对较少。笔者拟采用不同浓度的NaHCO3和Na2CO3混合溶液对唐古特白刺进行碱胁迫处理,研究不同浓度碱胁迫下唐古特白刺的生长、生理响应及叶片超微结构的变化,探讨唐古特白刺的耐碱能力及耐碱机制,以期为实现唐古特白刺在抑制盐碱地土壤退化和实现盐碱化土壤的持续生物改良提供参考。
1 材料与方法
1.1 种子采集及幼苗培养
试验所用唐古特白刺种子取自内蒙古巴彦淖尔市磴口县的白刺产地(海拔1 036 m,E 106°28′,N 40°32′)。挑选大小一致、颗粒饱满的种子于0.05% KMnO4溶液避光浸泡12 h,蒸馏水冲洗干净备用。
采用纯沙进行幼苗培养。将0.5% KMnO4溶液杀菌后的纯沙置于16 cm×14 cm营养钵中,每钵播白刺种子20粒,共20钵,种植深度约4 cm,置于温室内培养。培养条件为平均温度26 ℃,光照时间14 h,光照强度1 200 Lux,每3天浇灌蒸馏水100 mL,沙子湿度约为饱和含水量的60%。
1.2 碱胁迫处理
碱溶液配置:NaHCO3和Na2CO3按摩尔比1∶1混合溶解于蒸馏水,设置4个浓度梯度:200、300、400、500 mmol/L。待实生苗长出6~8片真叶时,挑选长势健壮、均匀一致的幼苗15钵,每钵6株,每个浓度处理幼苗3钵,每隔3 d浇灌不同浓度的碱溶液100 mL,对照组为蒸馏水100 mL,浇灌后及时将托盘中渗出的溶液倒回营养钵内,以防盐分流失。处理45 d后进行生长及生理指标的测定。
从剩余5钵中取上述同样标准的实生苗,每钵3株,将其根部直接浸于预先装有不同浓度碱液的三角瓶中进行胁迫处理,对照组浸于蒸馏水中。待处理12 h后有幼苗叶片开始发黄或者萎蔫时,取幼苗中上部第2节完好叶片,切成适当小段备用于超微结构观察。
1.3 指标测定及超微结构观察
1.3.1 植株生长及生理指标测定 分别于碱胁迫试验前和结束后测量幼苗株高;试验结束后,每个处理随机选取幼苗3株,蒸馏水冲洗干净,置105 ℃烘箱杀青15 min,90 ℃烘干至恒质量,计算单株生物量。
根系活力测定采用氯化三苯基四氮唑法;电解质渗透率测定采用电导仪法;叶绿素含量测定采用乙醇-丙酮混合液法;MDA含量测定采用硫代巴比妥酸法;SOD活性测定采用氮蓝四唑法;CAT活性测定采用紫外吸收法;脯氨酸含量测定采用茚三酮比色法[5]。
1.3.2 透射电镜样品的制备与观察 取各处理叶片相同部位(避开叶脉)切成1 mm×3 mm的矩形小块,样品在3%的戊二醛溶液中4 ℃固定24 h以上,0.1 mol/L(pH=7.0)磷酸缓冲液漂洗3次,每次15 min,1%锇酸固定2 h后,重复使用0.1 mol/L(pH=7.0)磷酸缓冲液漂洗样品3次,每次15 min。再以30%、50%、70%、80%、90%、95%乙醇逐级脱水1次,每级15 min,100%乙醇脱水2次,每次20 min[6]。之后进行EPON812环氧树脂浸透、包埋、聚合,莱卡EMUC6超薄切片机切片(切片厚度为70~90 nm),收集到镍网上,经醋酸铀30 min、柠檬酸铅10 min双重染色,最后利用日立H-600透射电镜进行观察,拍片[6]。
1.4 数据分析
所有数据均采用方差分析和Duncan′s多重检验,在P<0.05时用R 3.3.1对每个参数进行检验。参数值以标准误差(n=3)表示。
2 结果与分析
2.1 碱胁迫对唐古特白刺生长及生理指标的影响
由图1可知,唐古特白刺株高、生物量、根系活力及叶绿素含量随碱胁迫浓度的增大而下降,且均显著低于对照组。其中,>300 mmol/L的碱胁迫浓度对生物量抑制作用极为显著,生物量降幅达61.8%~81.8%。根系活力对碱胁迫较敏感,200 mmol/L时即显著下降,降幅为58.3%,500 mmol/L时降幅可达83.3%。叶绿素含量随碱胁迫浓度的增大存在22.0%~54.5%的降幅,但300 mmol/L和400 mmol/L碱胁迫处理间无显著差异。
与对照相比,碱胁迫能引起唐古特白刺叶片丙二醛含量和电解质渗透率的增加,且不同浓度处理间存在显著差异(图1)。随碱胁迫浓度的增大,丙二醛含量存在21.4%~114%的增幅,而电解质渗透率在碱胁迫200 mmol/L时即存在55.6%的增幅,500 mmol/L时增幅可达100%。
由图2可知,唐古特白刺叶片SOD、CAT活性和脯氨酸含量呈碱胁迫低浓度上升、高浓度下降趋势。与对照相比,除500 mmol/L碱胁迫外,其余胁迫浓度均可诱导这3个指标的显著增加,且不同胁迫浓度间差异显著。其中,CAT活性在碱胁迫200 mmol/L时增幅即为220%,300 mmol/L时活性达到最大,增幅为560%;而叶片SOD活性和脯氨酸含量在碱胁迫400 mmol/L时才达到最大,二者的增幅分别为250%和100%。
2.2 碱胁迫对唐古特白刺叶片超微结构的影响
2.2.1 叶肉细胞细胞壁与细胞质膜的变化 对照组中唐古特白刺叶肉细胞细胞壁光滑且透亮,细胞质膜无褶皱或突起(图3)。碱胁迫200 mmol/L时细胞质膜内陷或向内折叠不明显,很少有质膜突起;碱胁迫300 mmol/L时细胞壁有黑色颗粒出现;碱胁迫400 mmol/L时细胞质膜突起呈大小、形状不一的小泡,细胞壁不再光滑,有溶解迹象;碱胁迫500 mmol/L时细胞质膜褶皱或突起明显,细胞壁层次变得模糊(图3)。
2.2.2 叶肉细胞叶绿体形态及其分布的变化 对照组中唐古特白刺叶肉细胞叶绿体为香蕉形,短轴约为长轴的1/4,分布在与空气接触的细胞壁一边(图4)。随碱胁迫浓度的增加,叶绿体开始膨胀、变形并脱离质膜。与对照相比,叶绿体在碱胁迫300 mmol/L时开始肿胀变形为肾形,400 mmol/L时变为龟背形,500 mmol/L时变为花生形,短轴和长轴比例失衡,整个叶绿体离开质膜,且在400 mmol/L胁迫时个别叶绿体甚至游离向细胞中央的位置靠拢(图4)。

图1 碱胁迫对唐古特白刺株高、生物量、根系活力、叶绿素质量分数、丙二醛含量以及电解质渗透率的影响Fig.1 Effects of alkali stress on plant height, biomass, root activity, chlorophyll content, MDA (malondialdehyde) content and electrolyte permeability of Nitraria tangutorum
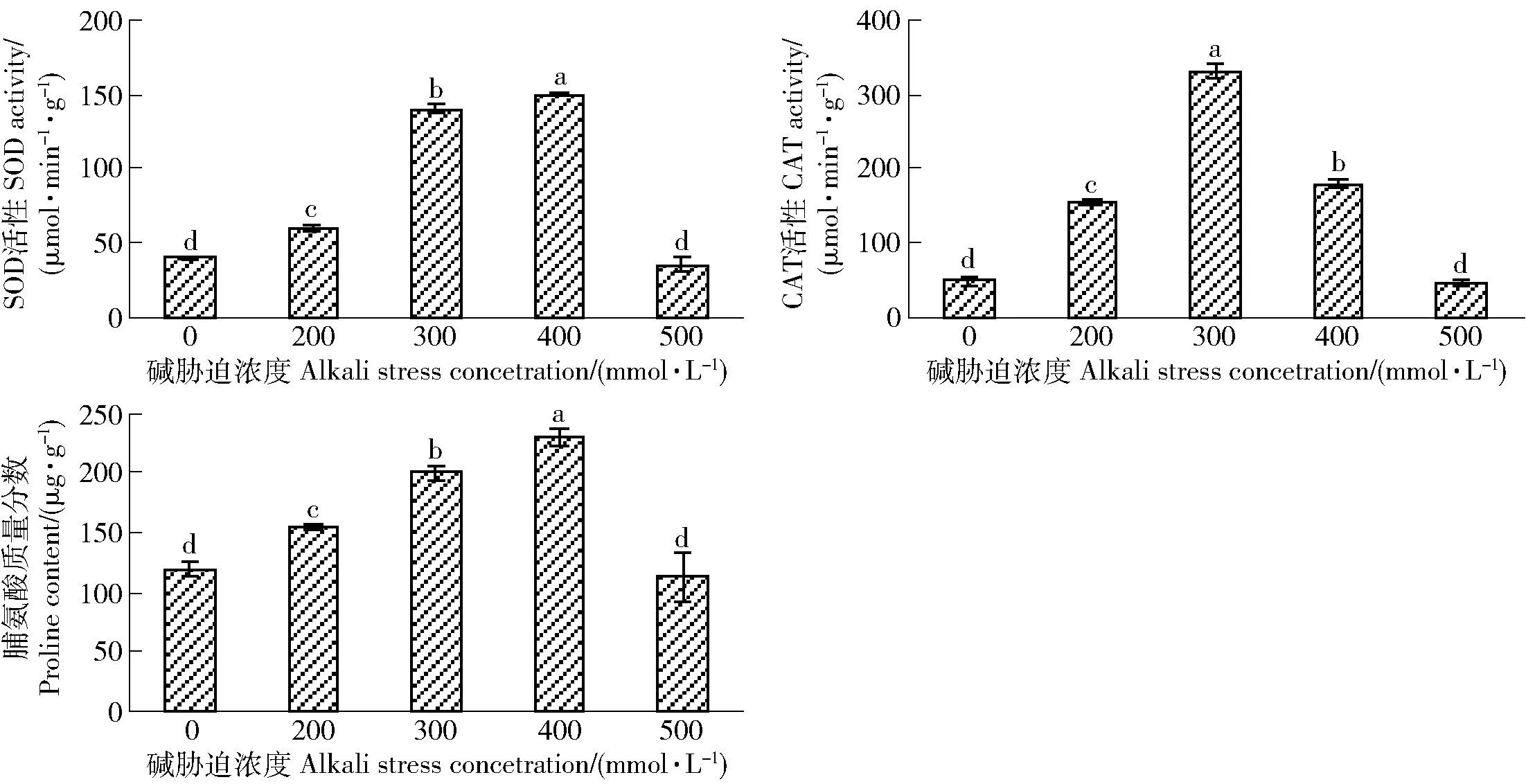
图2 碱胁迫对唐古特白刺SOD、CAT活性和脯氨酸质量分数的影响Fig.2 Effects of alkali stress on SOD (superoxide dismutase), CAT (catalase) activity and proline content of Nitraria tangutorum
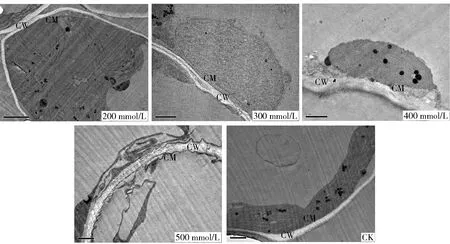
CW: 细胞壁 Cell wall. CM: 细胞膜 Cell membrane.图3 不同浓度碱胁迫下唐古特白刺叶肉细胞壁和细胞膜的变化Fig.3 Mesophyll cell wall and cell membrane changes of Nitraria tangutorum under different alkali stress
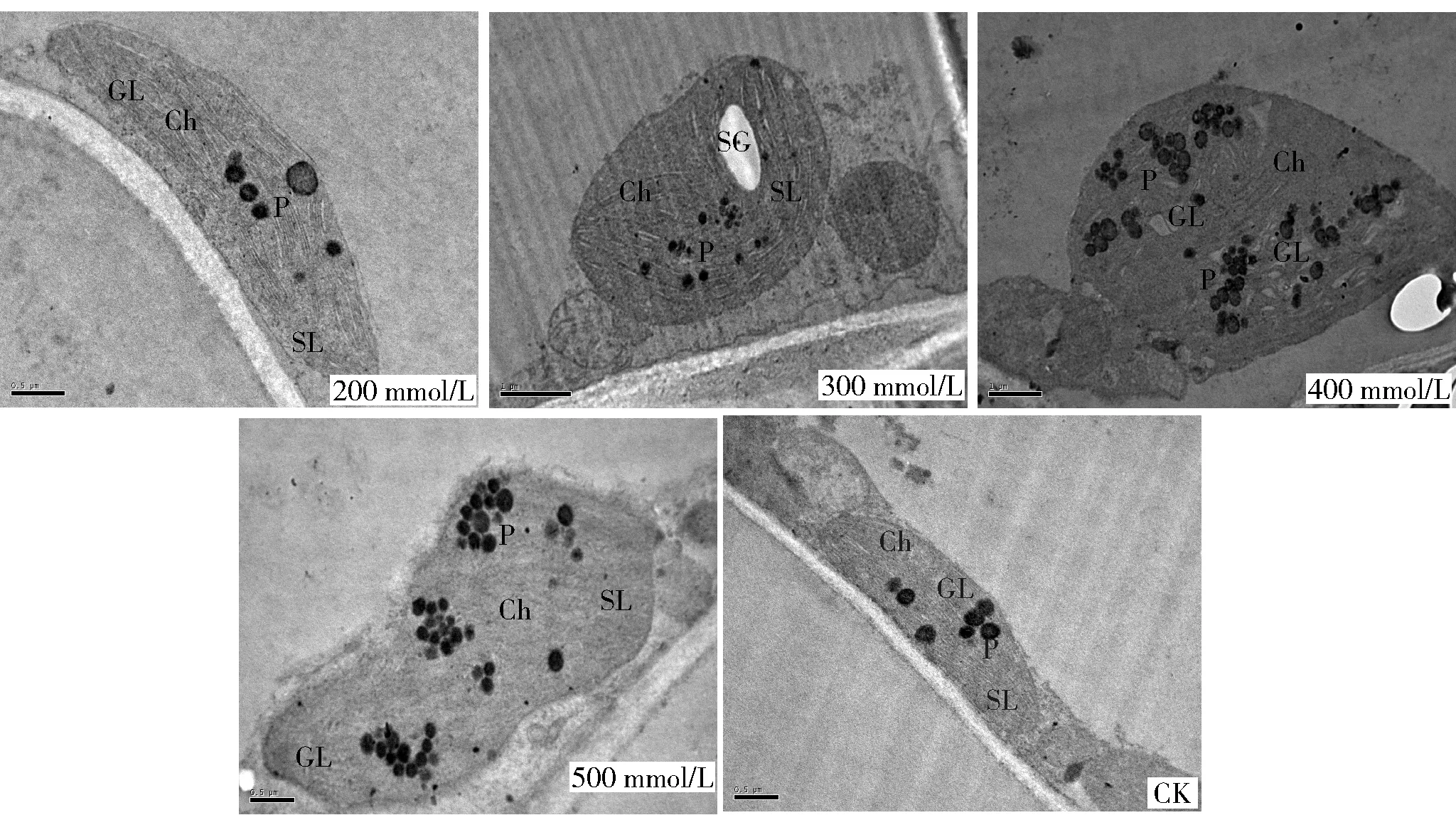
GL: 基粒片层 Grana lamella. Ch: 叶绿体 Chloroplas. SL: 基质片层 Stroma lamellae. SG: 淀粉粒 Starch granules. P: 质粒 Plastoglobules. 图4 不同浓度碱胁迫下唐古特白刺叶绿体形态和分布的变化Fig.4 Mesophyll chloroplast morphology and distribution of Nitraria tangutorum under different alkali stress
2.2.3 叶肉细胞叶绿体类囊体结构的变化 对照组中唐古特白刺叶肉细胞叶绿体外膜完整平滑,无折叠现象,膜间隙与基质被内膜明显隔离开,基质呈均质状,类囊体沿叶绿体长轴平行有序排列,基粒片层结构紧致清晰(图4)。基粒类囊体在碱胁迫300 mmol/L时排列散乱,400 mmol/L时出现囊泡化,500 mmol/L时有溶解,片层结构消失迹象(图4)。
2.2.4 叶肉细胞叶绿体淀粉粒和嗜锇颗粒的变化 300 mmol/L碱胁迫时唐古特白刺叶肉细胞叶绿体中间膨大部位出现淀粉粒,淀粉粒长度约为叶绿体长度的1/5,沿叶绿体长轴分布(图4)。其他浓度碱胁迫下叶绿体中未见淀粉粒的出现。碱胁迫处理下,叶绿体中均出现嗜锇颗粒,且其数量、体积较对照都有不同程度增加。碱胁迫400 mmol/L和500 mmol/L时有大量嗜锇颗粒集聚成堆出现(图4)。
3 讨论
3.1 碱胁迫下唐古特白刺的生长及生理响应
生长抑制是植物在盐碱胁迫下的综合表现[7],本研究中碱胁迫后唐古特白刺株高、生物量的降低即是最好的例证。逆境除抑制植物生长外,还会引起一系列的生理代谢紊乱,而植物通过相应的生理变化来适应或抵制逆境胁迫[8]。非生物胁迫中,根系是首先并直接遭受伤害的部位[9]。低浓度碱胁迫时唐古特白刺根系活力即显著下降,可能由于碱胁迫造成其根系周围的高pH值引起金属离子和磷的积累,损害根部组织结构,导致根系吸收功能减弱,根系活力降低[10]。叶绿素是光合作用的主要色素[11],随碱胁迫浓度的增大,其含量减少,可能由于碱胁迫使叶绿素酶活性降低,叶绿素分解加速,或者碱胁迫使其叶绿体蛋白和叶绿素结合受到影响,导致更多的叶绿素受损[12]。另有研究表明,碱胁迫处理可能打破了镁离子在植物体内的正常状态,从而导致叶绿素形成受阻[11]。同时,唐古特白刺叶片MDA含量及电解质渗透率随碱胁迫浓度的增大而持续上升,这与张磊等[10]的研究结果相似,即碱胁迫浓度的增大会加剧膜脂过氧化程度,破坏质膜的选择透过性,导致电解质大量外渗,加剧对细胞膜的伤害程度。
本研究中,唐古特白刺叶片SOD和CAT活性在低浓度碱胁迫时上升,高浓度碱胁迫时下降。由此可知,一定浓度的碱胁迫可以诱导SOD和CAT活性的增加。SOD和CAT作为生物防御体系的关键酶,可有效保护细胞免受活性氧伤害[13]。低浓度碱胁迫时,唐古特白刺可能通过增强SOD和CAT酶活性以消除和转化超氧阴离子及过氧化氢,维持细胞正常的代谢活动。但高浓度碱胁迫时,细胞代谢紊乱加剧,产生过多超氧阴离子及过氧化氢,使SOD和CAT大量消耗,同时,膜脂过氧化加剧,膜系统损伤加重,影响SOD和CAT的分泌和激活过程,从而导致SOD和CAT活性的下降[13]。另试验发现,300 mmol/L碱胁迫时唐古特白刺叶片CAT活性显著升高,其原因可能是CAT对过氧化氢亲和力较低,只有当SOD分解超氧阴离子产生的过氧化氢得到积累时CAT活性才能被大幅度激活。
脯氨酸(proline)是植物在逆境胁迫下累积的主要有机分子[14]。曲元刚等[9]研究发现,Na2CO3引起的Pro增加幅度远大于NaCl,认为Pro是植物体内调节pH的一种缓冲物质。本研究中,Pro含量在碱胁迫浓度≤400 mmol/L时得以积累可能由于较低浓度的碱胁迫促使Pro合成酶的活化,抑制Pro的生物降解造成的[8]。也有研究表明,逆境胁迫导致线粒体膜对Pro选择透性下降也是造成胞质内Pro积累的重要原因[15]。500 mmol/L碱胁迫时Pro含量减少可能由于高浓度碱胁迫下细胞代谢紊乱,叶绿体、线粒体结构受损严重,酶活性降低,而催化Pro合成的酶位于胞质或叶绿体中,氧化降解酶位于线粒体中[15],导致Pro合成受阻,含量减少。
3.2 碱胁迫下唐古特白刺超微结构的变化
膜结构和细胞器的破坏,是植物受到盐害的普遍特征,其破坏程度与植物的耐盐碱性息息相关[16]。唐古特白刺经碱胁迫后,细胞壁不再光滑,出现沉淀或溶解趋势;细胞质膜则出现褶皱或突起产生形状、大小不一的小泡,其中以高浓度胁迫时变化最为显著。这些变化不仅说明碱胁迫对唐古特白刺细胞壁及质膜造成一定损伤,也说明其利用自身结构的改变来积极适应不良环境,既可以消除过多破损的膜,加快更新,也可以把过多的盐分收集在液泡中,提高液泡浓度,增加细胞的渗透势[17]。
叶绿体是众多细胞器中对盐碱胁迫最为敏感的[18]。近年来研究者们通过对逆境胁迫下桂花(Osmanthusfragrans)[16]、核桃(Juglansregia)[19]、棉花(Gossypiumhirsutum)[20]、刺槐(Robiniapseudoacacia)[21]等叶绿体超微结构的研究,发现由于不同植物对逆境的适应能力不同,导致叶绿体形态变化存在差异,同时,逆境胁迫可导致叶绿体类囊体排列松散变形,基粒片层垛叠数量减少或模糊不清,破坏叶绿体被膜甚至失去完整膜结构等[16]。笔者发现,碱胁迫处理下的唐古特白刺叶绿体由香蕉形变为肾形、龟背形和花生形不等,这首先从外部轮廓上印证了不同浓度碱胁迫对叶绿体损伤程度的大小不一。其次,各碱胁迫浓度下叶绿体均不同程度的剥离细胞壁,而植物叶绿体通常紧贴细胞壁分布于与空气接触的质膜旁,这样的分布有利于叶绿体与外界进行气体交换,当叶绿体离开质膜时,叶绿体内正常光合运转功能可能受阻。叶绿体剥离细胞壁可能是由于随碱胁迫浓度的增加,叶绿体膜受损发生渗漏,体积缩小造成的。
300 mmol/L碱胁迫时唐古特白刺叶绿体中淀粉粒的出现,可能由于线粒体膜受到损伤,出现能量供应障碍,导致光合产物以淀粉粒的形式沉积,也可能是由于叶绿体结构受损,高浓度盐碱阻碍淀粉的水解和向外运输造成的[7]。叶绿体内淀粉粒补偿性增多,体积变大,可弥补逆境胁迫造成的能量供应不足,提高细胞质浓度,保证水和无机营养的供给[13]。未胁迫时,唐古特白刺叶绿体中嗜锇颗粒数量相对较少,体积相对较小,而高浓度碱胁迫时数量增多,体积变大,且集聚成堆出现。嗜锇颗粒的存在可以提供脂质供膜的更新,以保持膜结构的完整性,同时,逆境胁迫时嗜锇颗粒会降解为糖,提高细胞中糖的浓度,进而增加植物的抗性[17]。因此,淀粉粒的出现和嗜锇颗粒的增多变大对唐古特白刺完成在极端环境下的生活史意义重大[6],也进一步验证了唐古特白刺具有一定的耐盐碱性。
4 结论
低浓度碱胁迫对唐古特白刺生长具有一定抑制作用,≥300 mmol/L的碱胁迫浓度对白刺生长及叶片超微结构产生明显的抑制及损伤。碱胁迫下SOD、CAT活性和Pro质量分数的提高,嗜锇小体增多变大以及淀粉粒的出现,表现唐古特白刺对碱胁迫有较强的适应能力。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
蔬菜(2022年12期)2022-01-01 11:32:10
农村百事通(2019年1期)2019-01-30 13:36:34
农学学报(2018年10期)2018-11-01 01:19:30
中成药(2018年7期)2018-08-04 06:04:06
西夏学(2018年1期)2018-04-29 09:09:02
西夏研究(2017年1期)2017-07-10 08:16:55
西南农业学报(2016年5期)2016-05-17 05:42:28
农家顾问(2014年11期)2014-12-01 13:29:34
天然产物研究与开发(2014年6期)2014-04-27 14:15:58
